
Rapid Creation of Interspecific Hybrid Progeny to Broaden Genetic Distance through Double Haploid (DH) Inducer in Brassica napus


Abstract
:1. Introduction
2. Results
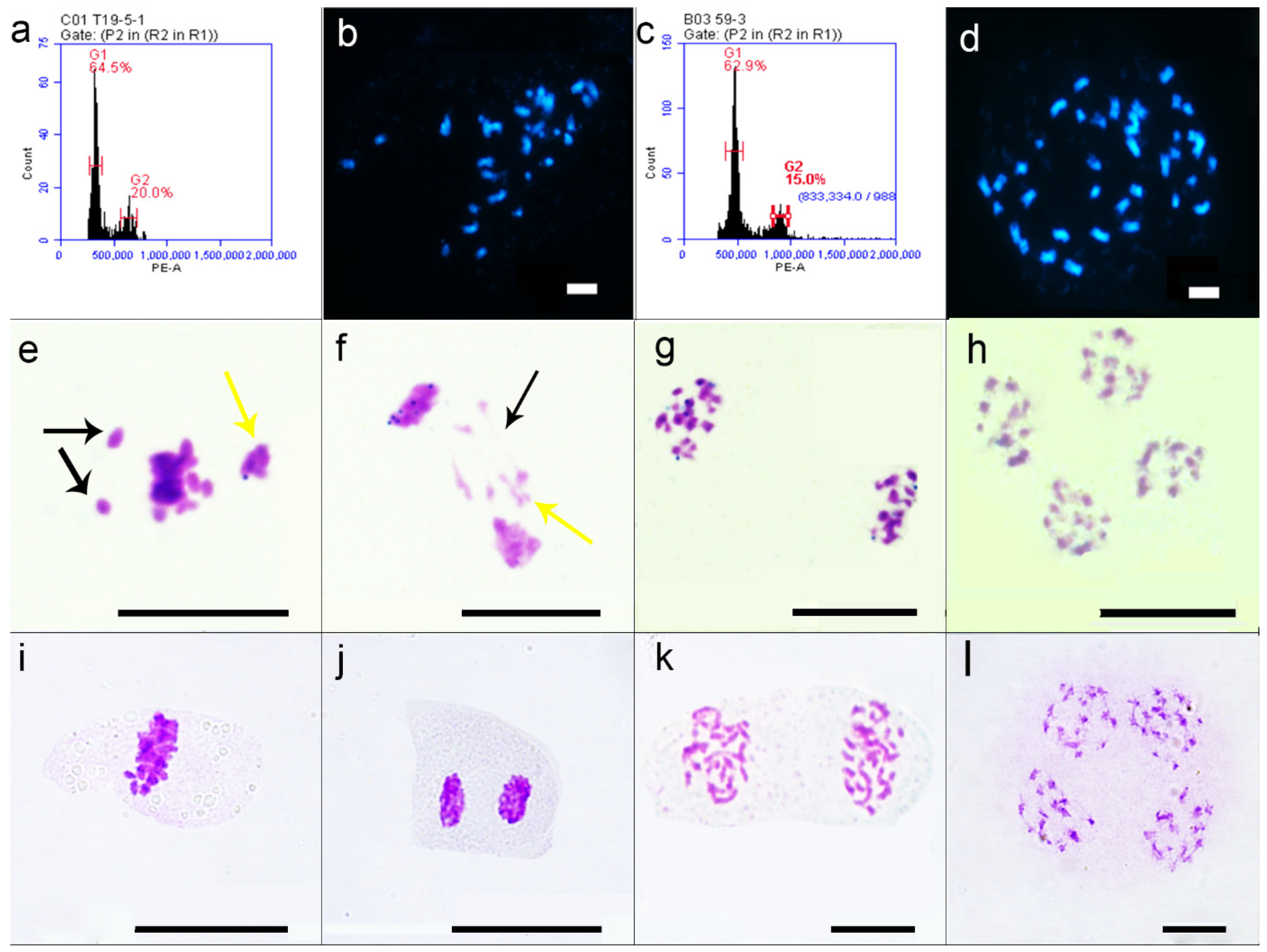
2.1. Phenotype, Fertility, and Flow Cytometric Identification of Interspecific Hybrid F1 and Induced F1 Plants
2.2. Cytological Observation of Interspecies Hybrid F1 and Induced F1
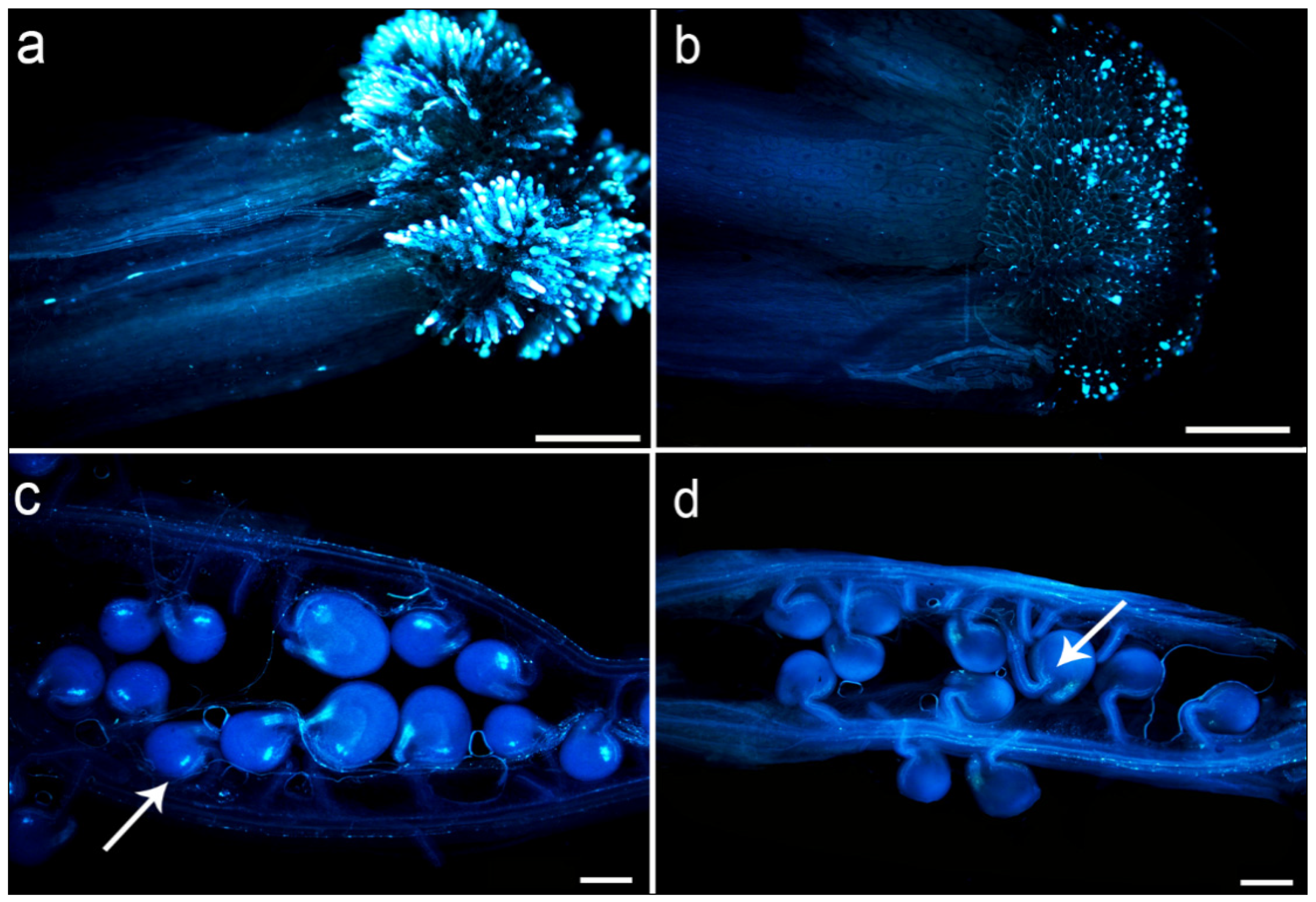
2.3. Observation of Germination, Fertilization Rate, and Seed Setting Rate in Interspecific Hybrid F1 Pollen Tube
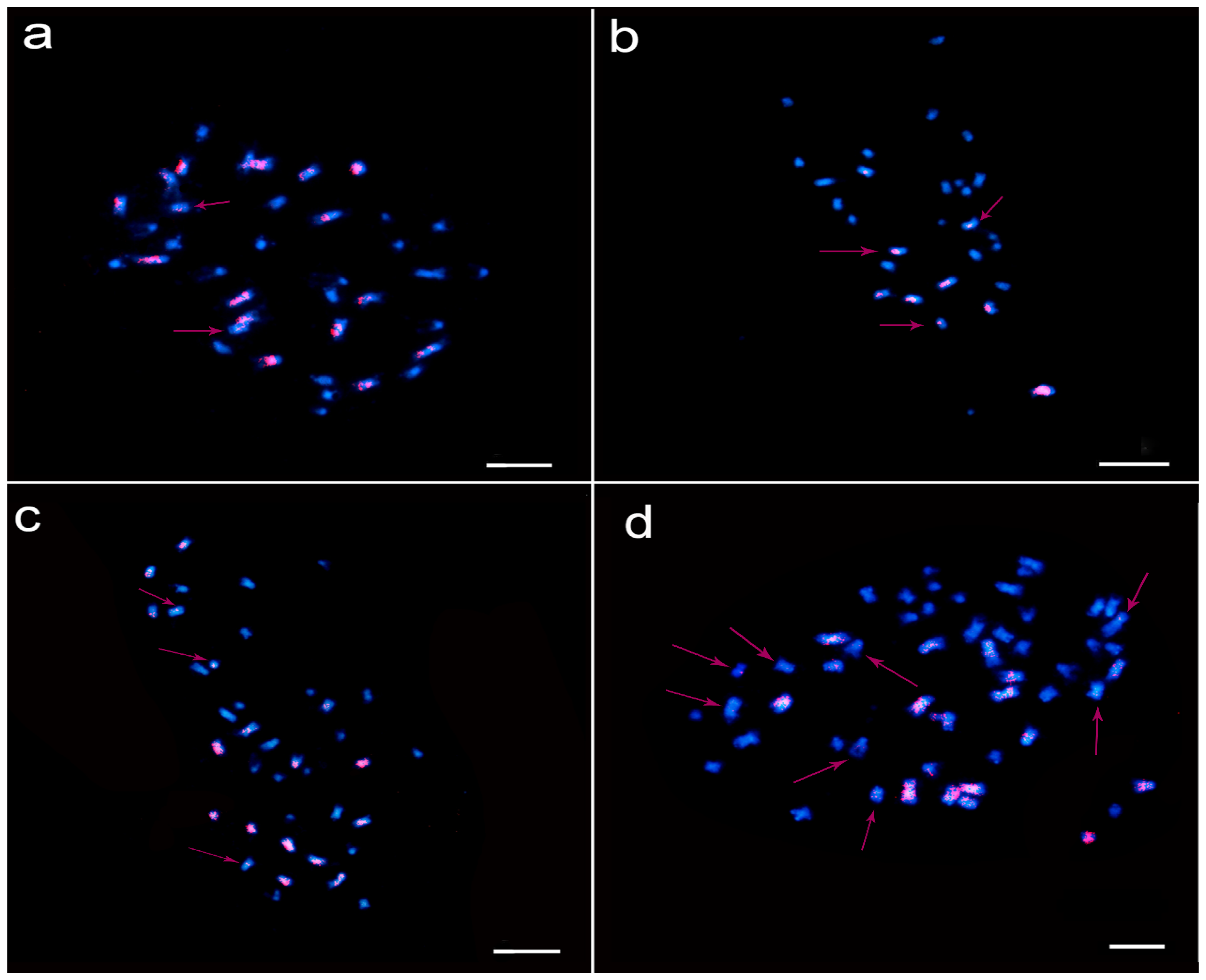
2.4. Interspecific Hybridization F1 and Induced F1 Fluorescence In Situ Hybridization
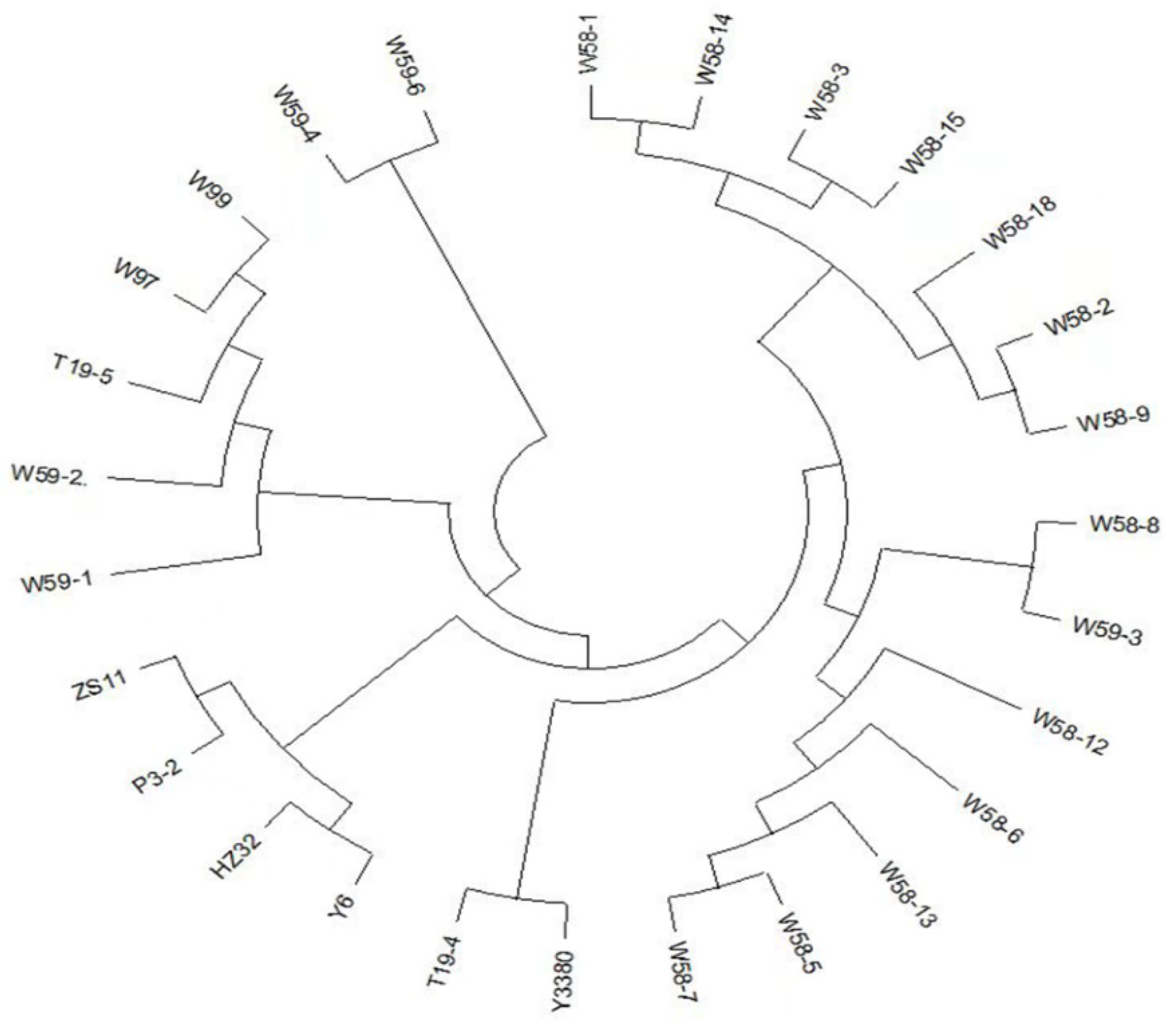
2.5. Homozygous Rate and Genetic Distance Cluster Analysis of Induced F1 Generation SNP Loci
3. Discussion
3.1. The Reasons for the Low Seed Setting Rate of Interspecific Hybridization between B. napus and B. rapa and the Genetic Stability of Egg Cells Provides the Possibility for DH to Induce Egg Cells
3.2. Ploidy Changes and Cytological Genetic Stability of Induced F1 Progeny by Interspecific Hybridization
3.3. The F1 Progeny Produced by the Interspecific Hybridization Retains the Rich Genetic Variation of B. napus and B. napus
3.4. The Important Role of DH Inducible Lines in Obtaining Interspecific Hybrid Progeny
4. Materials and Methods
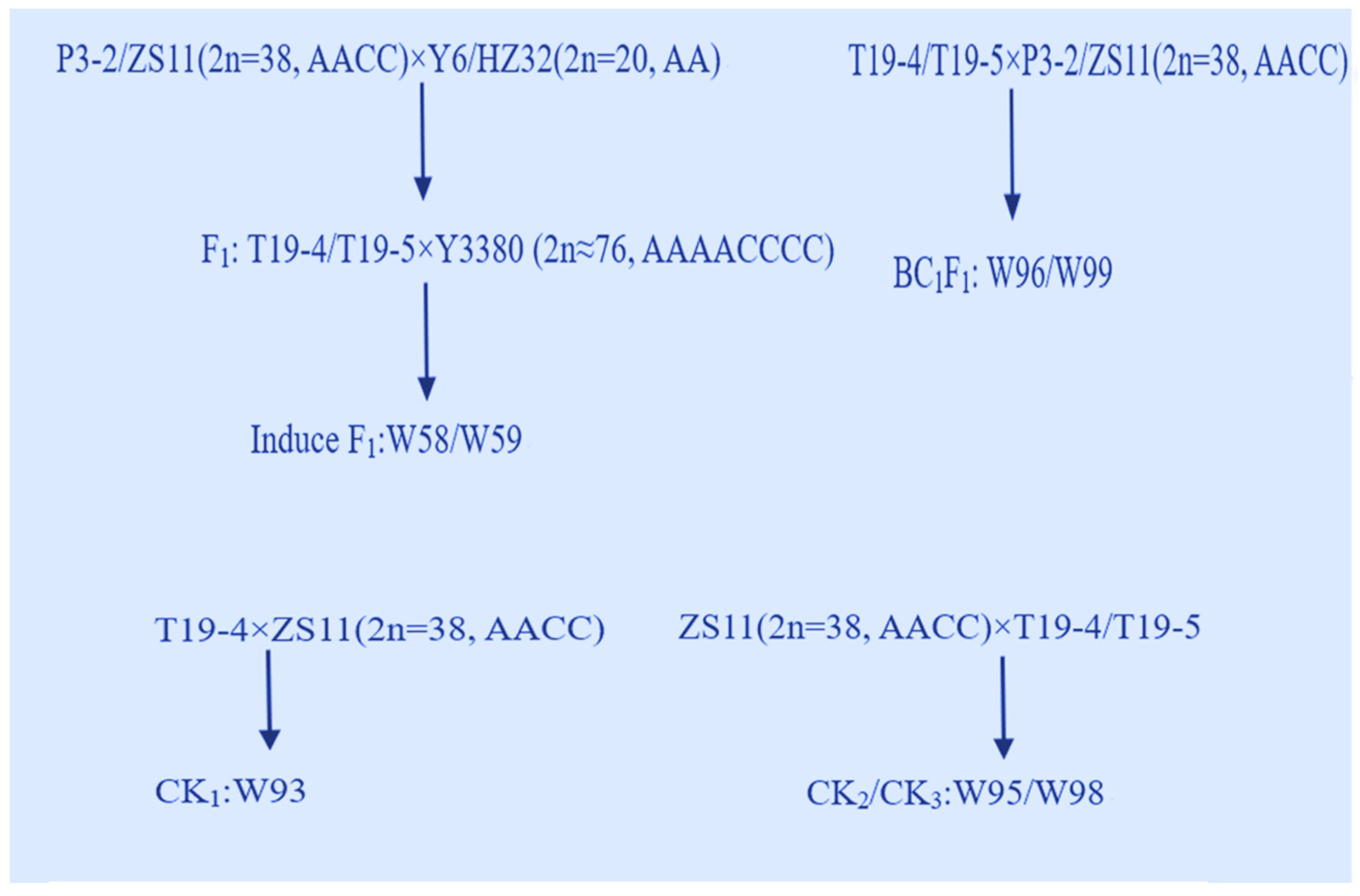
4.1. Planting Material and Hybrid Combination
4.2. Morphological Analysis
4.3. Flow Cytometry to Identify Ploidy
4.4. General Cytology Observation
4.5. Germination Dynamics of Pollen Tube in Silique
4.6. Fluorescence In Situ Hybridization
4.7. Analysis of Homozygous Rate and Genetic Distance of SNP Loci
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Qian, W.; Meng, J.; Li, M.; Frauen, M.; Sass, O.; Noack, J.; Jung, C. Introgression of genomic components from Chinese Brassica rapa contributes to widening the genetic diversity in rapeseed (B. napus L.), with emphasis on the evolution of Chinese rapeseed. Theor. Appl. Genet. 2006, 113, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Jesske, T.; Olberg, B.; Schierholt, A.; Becker, H.C. Resynthesized lines from domesticated and wild Brassica taxa and their hybrids with B. napus L.: Genetic diversity and hybrid yield. Theor. Appl. Genet. 2013, 126, 1053–1065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaebelein, R.; Alnajar, D.; Koopmann, B.; Mason, A.S. Hybrids between Brassica napus and B. nigra show frequent pairing between the B and A/C genomes and resistance to blackleg. Chromosome Res. 2019, 27, 221–236. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.; Shi, S.; Gan, L.; Li, Z.; Qu, X. The production of yellow-seeded Brassica napus (AACC) through crossing interspecific hybrids of B. campestris (AA) and B. carinata (BBCC) with B. napus. Euphytica 1998, 103, 329–333. [Google Scholar] [CrossRef]
- Jiang, J.; Wang, Y.; Xie, T.; Rong, H.; Li, A.; Fang, Y.; Wang, Y. Metabolic characteristics in meal of black rapeseed and yellow-seeded progeny of Brassica napus–Sinapis alba hybrids. Molecules 2015, 20, 9761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, A.; Wei, C.; Jiang, J.; Zhang, Y.; Snowdon, R.J.; Wang, Y. Phenotypic variation in progenies from somatic hybrids between Brassica napus and Sinapis alba. Euphytica 2009, 170, 289–296. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Ding, G.; Li, L.; Cai, H.; Ye, X.; Zou, J.; Xu, F. Identification and characterization of improved nitrogen efficiency in interspecific hybridized new-type Brassica napus. Ann. Bot. 2014, 114, 549–559. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Liu, X.; Li, X.; Liu, X.; Hu, Y.; Chen, T. Obtaining and genetic analysis of hybrid progeny between Brassica napus cv. Xiangyou 15 and Brassica campestris ssp. pekinensis. Acta Agric. Zhejiangensis 2021, 33, 1170–1176. [Google Scholar]
- Pengfang, Z.; Xing, W.; Jian, Z.; Yutang, W. Studies on the Compatibility of Backcrosses of Interspecific and Inter Varietal Hybrids in Brassica. Chin. Agric. Sci. Bull. 2011, 27, 149–152. [Google Scholar]
- Mikkelsen, T.; Jensen, J.; Jørgensen, R.B. Inheritance of oilseed rape (Brassica napus) RAPD markers in a backcross progeny with Brassica campestris. Theor. Appl. Genet. 1996, 92, 492–497. [Google Scholar] [CrossRef] [PubMed]
- Ferrie, A. Microspore culture of Brassica species. In Doubled Haploid Production in Crop Plants; Springer: Berlin/Heidelberg, Germany, 2003; pp. 205–215. [Google Scholar] [CrossRef]
- Hansen, M. Protocol for microspore culture in Brassica. In Doubled Haploid Production in Crop Plants; Springer: Berlin/Heidelberg, Germany, 2003; pp. 217–222. [Google Scholar] [CrossRef]
- Ferrie, A.M.; Möllers, C. Haploids and doubled haploids in Brassica spp. for genetic and genomic research. Plant Cell Tissue Organ Cult. (PCTOC) 2011, 104, 375–386. [Google Scholar] [CrossRef] [Green Version]
- Barro, F.; Fernandez-Escobar, J.; De La Vega, M.; Martin, A. Doubled haploid lines of Brassica carinata with modified erucic acid content through mutagenesis by EMS treatment of isolated microspores. Plant Breed. 2001, 120, 262–264. [Google Scholar] [CrossRef]
- Zhang, W.; Fu, Q.; Dai, X.; Bao, M. The culture of isolated microspores of ornamental kale (Brassica oleracea var. acephala) and the importance of genotype to embryo regeneration. Sci. Hortic. 2008, 117, 69–72. [Google Scholar] [CrossRef]
- Shumilina, D.; Kornyukhin, D.; Domblides, E.; Soldatenko, A.; Artemyeva, A. Effects of Genotype and Culture Conditions on Microspore Embryogenesis and Plant Regeneration in Brassica Rapa ssp. Rapa L. Plants 2020, 9, 278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ari, E. Shed-Microspore Culture in Ornamental Peppers for Doubled Haploid Plant Production. In Doubled Haploid Technology; Springer: Berlin/Heidelberg, Germany, 2021; pp. 251–266. [Google Scholar] [CrossRef]
- Fu, S.; Yin, L.; Xu, M.; Li, Y.; Wang, M.; Yang, J.; Fu, T.; Wang, J.; Shen, J.; Ali, A. Maternal doubled haploid production in interploidy hybridization between Brassica napus and Brassica allooctaploids. Planta 2017, 247, 113–125. [Google Scholar] [CrossRef]
- Shaohong, F.; Li, Y.; Yang, J.; Wang, J.; Zou, Q.; Lanrong, T.; Zeming, K.; Rong, T. Method for Breeding Brassica napus Varieties and Materials with Double Haploid Induction Line of Rapeseed. U.S. Patent No. 16/310,497, 22 August 2019. [Google Scholar]
- Li, C.; Sang, S.; Sun, M.; Yang, J.; Shi, Y.; Hu, X.; Li, Y.; Hao, M.; Chu, W.; Zhang, H.; et al. Direct Modification of Multiple Gene Homoeologs in Brassica oleracea and Brassica napus Using Doubled Haploid Inducer-Mediated Genome Editing System. Plant Biotechnol. J. 2021, 19, 1889–1891. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Ma, Y.; Zhu, Z.; Huang, L.; Ali, A.; Luo, X.; Zhou, Y.; Li, Y.; Xu, P.; Yang, J. Maternal karyogene and cytoplasmic genotype affect the induction efficiency of doubled haploid inducer in Brassica napus. BMC Plant Biol. 2021, 21, 207. [Google Scholar] [CrossRef]
- Luo, X.; Yang, J.; Zhu, Z.; Huang, L.; Ali, A.; Javed, H.H.; Zhang, W.; Zhou, Y.; Yin, L.; Xu, P. Genetic characteristics and ploidy trigger the high inducibility of double haploid (DH) inducer in Brassica napus. BMC Plant Biol. 2021, 21, 538. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Kato, M. Fertilization fitness and relation to chromosome number in interspecific progeny between Brassica napus and B. napus: A comparative study using natural and resynthesized B. napus. Breed. Sci. 2001, 51, 73–81. [Google Scholar] [CrossRef] [Green Version]
- Leflon, M.; Eber, F.; Letanneur, J.-C.; Chelysheva, L.; Coriton, O.; Huteau, V.; Ryder, C.; Barker, G.; Jenczewski, E.; Chevre, A. Pairing and recombination at meiosis of Brassica rapa (AA)× Brassica napus (AACC) hybrids. Theor. Appl. Genet. 2006, 113, 1467–1480. [Google Scholar] [CrossRef]
- Nicolas, S.D.; Mignon, G.L.; Eber, F.; Coriton, O.; Monod, H.; Clouet, V.; Huteau, V.; Lostanlen, A.; Delourme, R.; Chalhoub, B. Homeologous recombination plays a major role in chromosome rearrangements that occur during meiosis of Brassica napus haploids. Genetics 2007, 175, 487–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francis, K.E.; Lam, S.Y.; Harrison, B.D.; Bey, A.L.; Berchowitz, L.E.; Copenhaver, G.P. Pollen tetrad-based visual assay for meiotic recombination in Arabidopsis. Proc. Natl. Acad. Sci. USA 2007, 104, 3913–3918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, L.; Zhu, Z.; Luo, X.; Huang, L.; Li, Y.; Mason, A.S.; Yang, J.; Ge, X.; Long, Y.; Wang, J. Genome-Wide Duplication of Allotetraploid Brassica napus Produces Novel Characteristics and Extensive Ploidy Variation in Self-Pollinated Progeny. G3 Genes Genomes Genet. 2020, 10, 3687–3699. [Google Scholar] [CrossRef] [PubMed]
- Chalhoub, B.; Denoeud, F.; Liu, S.; Parkin, I.A.; Tang, H.; Wang, X.; Chiquet, J.; Belcram, H.; Tong, C.; Samans, B. Early allopolyploid evolution in the post-Neolithic Brassica napus oilseed genome. Science 2014, 345, 950–953. [Google Scholar] [CrossRef] [Green Version]
- Rygulla, W.; Snowdon, R.; Eynck, C.; Koopmann, B.; Von Tiedemann, A.; Lühs, W.; Friedt, W. Broadening the genetic basis of Verticillium longisporum resistance in Brassica napus by interspecific hybridization. Phytopathology 2007, 97, 1391–1396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peterka, H.; Budahn, H.; Schrader, O.; Ahne, R.; Schütze, W. Transfer of resistance against the beet cyst nematode from radish (Raphanus sativus) to rape (Brassica napus) by monosomic chromosome addition. Theor. Appl. Genet. 2004, 109, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Zhu, Z.; Huang, L.; Luo, X.; Li, Y.; Xiao, C.; Yang, J.; Wang, J.; Zou, Q.; Tao, L.; et al. DNA repair- and nucleotide metabolism-related genes exhibit differential CHG methylation patterns in natural and synthetic polyploids (Brassica napus L.). Hortic. Res. 2021, 8, 142. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Meng, D.; Chen, S.; Luo, H.; Zhang, Q.; Jin, W.; Yan, J. Single nucleus sequencing reveals spermatid chromosome fragmentation as a possible cause of maize haploid induction. Nat. Commun. 2017, 8, 991. [Google Scholar] [CrossRef] [PubMed]
- Jugulam, M.; Ziauddin, A.; So, K.K.; Chen, S.; Hall, J.C. Transfer of dicamba tolerance from Sinapis arvensis to Brassica napus via embryo rescue and recurrent backcross breeding. PLoS ONE 2015, 10, e0141418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoekstra, F.A.; Bruinsma, J. Respiration and vitality of binucleate and trinucleate pollen. Physiol. Plant. 1975, 34, 221–225. [Google Scholar] [CrossRef]
- Ureta, M.S.; Carbonell, F.T.; Pandolfo, C.; Presotto, A.D.; Cantamutto, M.A.; Poverene, M. IMI resistance associated to crop-weed hybridization in a natural Brassica rapa population: Characterization and fate. Environ. Monit. Assess. 2017, 189, 101. [Google Scholar] [CrossRef] [PubMed]
- Crowley, L.C.; Chojnowski, G.; Waterhouse, N.J. Measuring the DNA content of cells in apoptosis and at different cell-cycle stages by propidium iodide staining and flow cytometry. Cold Spring Harb. Protoc. 2016, 2016, pdb-prot087247. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Tang, L.; Yin, Y.; Zhang, A.; Yu, Z.; Yang, E.; Tang, Z.; Fu, S.; Yang, Z. Molecular dissection of Secale africanum chromosome 6R afr in wheat enabled localization of genes for resistance to powdery mildew and stripe rust. BMC Plant Biol. 2020, 20, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Lang, T.; Li, G.; Wang, H.; Yu, Z.; Chen, Q.; Yang, E.; Fu, S.; Tang, Z.; Yang, Z. Physical location of tandem repeats in the wheat genome and application for chromosome identification. Planta 2019, 249, 663–675. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, H.; Higashiyama, T. Tip-localized receptors control pollen tube growth and LURE sensing in Arabidopsis. Nature 2016, 531, 245–248. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Z.; Pires, J.C. Karyotype and identification of all homoeologous chromosomes of allopolyploid Brassica napus and its diploid progenitors. Genetics 2011, 187, 37–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knytl, M.; Kalous, L.; Symonová, R.; Rylková, K.; Ráb, P. Chromosome studies of European cyprinid fishes: Cross-species painting reveals natural allotetraploid origin of a Carassius female with 206 chromosomes. Cytogenet. Genome Res. 2013, 139, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.; Chen, L.; Wang, Y.; Li, M.; Yang, Z.; Qiu, L.; Yan, B.; Ren, Z.; Tang, Z. Oligonucleotide probes for ND-FISH analysis to identify rye and wheat chromosomes. Sci. Rep. 2015, 5, 10552. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cross Combination | Average Fertilization Rate (the Number of Ovules with Fluorescent Dots in the Silique/Total Number of Ovules in the Silique) % | Average Effective Seed Setting Rate (the Number of Seeds/ the Number of Effective Siliques × 25) % |
|---|---|---|
| ZS11 × T19-4 | (10/43) 23.26 | (330/(115 × 25)) 11.48 |
| ZS11 × T19-5 | (9/34) 26.47 | (40/(21 × 25)) 7.62 |
| T19-4 × ZS11 | (35/58) 60.34 | (216/(38 × 25)) 22.74 |
| T19-5 × ZS11 | (45/65) 69.23 | (173/(29 × 25)) 23.86 |
| T19-4 × Y3380 | (47/56) 83.93 | (86/(13 × 25)) 26.46 |
| T19-5 × Y3380 | (28/38) 73.68 | (92/(18 × 25)) 20.44 |
| T19-4 ⊗ | (3/48) 6.25 | (13/(13 × 25)) 4.00 |
| T19-5 ⊗ | (5/42) 11.90 | (35/(27 × 25)) 5.19 |
| ZS11 ⊗ | (48/53) 90.57 | (402/(32 × 25)) 50.25 |
| W58 ⊗ | No data | (572/(78 × 25)) 29.33 |
| W59 ⊗ | No data | (313/(32 × 25)) 39.13 |
| Sample Number | Total Number of Plants | Peak Range (D Thousand Lines) | Average Peak (D Thousand Lines) | Number of Tetraploid Plants | Number of Triploid Plants | Number of Other Ploidy Plants |
|---|---|---|---|---|---|---|
| P3-2 | 3 | 482.62–498.71 | 493.36 | 3 | 0 | 0 |
| Y6-1 | 3 | 261.16–313.41 | 287.18 | 0 | 0 | 2 |
| ZS11 | 3 | 495.89–510.69 | 502.73 | 3 | 0 | 0 |
| HZ32 | 3 | 243.76–303.69 | 274.48 | 0 | 0 | 2 |
| T19-4 | 27 | 312.12–482.46 | 351.02 | 1 | 26 | 0 |
| T19-5 | 23 | 307.29–493.68 | 349.64 | 1 | 22 | 0 |
| W58 | 18 | 421.45–661.17 | 531.65 | 15 | 0 | 3 |
| W59 | 6 | 478.27–1037.64 | 676.33 | 3 | 0 | 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Y.; Yang, M.; Zhao, S.; Shi, H.; Li, Y.; Gong, W.; Yang, J.; Wang, J.; Zou, Q.; Tao, L.; et al. Rapid Creation of Interspecific Hybrid Progeny to Broaden Genetic Distance through Double Haploid (DH) Inducer in Brassica napus. Plants 2022, 11, 695. https://doi.org/10.3390/plants11050695
Zhou Y, Yang M, Zhao S, Shi H, Li Y, Gong W, Yang J, Wang J, Zou Q, Tao L, et al. Rapid Creation of Interspecific Hybrid Progeny to Broaden Genetic Distance through Double Haploid (DH) Inducer in Brassica napus. Plants. 2022; 11(5):695. https://doi.org/10.3390/plants11050695
Chicago/Turabian StyleZhou, Ying, Meicui Yang, Shihui Zhao, Haoran Shi, Yun Li, Wanzhuo Gong, Jin Yang, Jisheng Wang, Qiong Zou, Lanrong Tao, and et al. 2022. "Rapid Creation of Interspecific Hybrid Progeny to Broaden Genetic Distance through Double Haploid (DH) Inducer in Brassica napus" Plants 11, no. 5: 695. https://doi.org/10.3390/plants11050695