
Anti-Inflammatory Functions of Methanol Extract from Malus baccata (L.) Borkh. Leaves and Shoots by Targeting the NF-κB Pathway

 ,
, 
Abstract
:1. Introduction
2. Results
2.1. Effect of Mb-ME on Nitric Oxide Production
2.2. Effect of Mb-ME on the mRNA Expression of Inflammatory Genes
2.3. Effect of Mb-ME on the Translocation of Transcription Factors
2.4. Effect of Mb-ME in Activating the NF-κB Upstream Signaling Pathway
2.5. Effect of Mb-ME on In Vivo EtOH/HCl-Induced Gastritis
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Plant Information and Extraction Methods
4.3. Cell Culture
4.4. Drug Preparation
4.5. Animals
4.6. Preparation of Peritoneal Macrophages
4.7. Nitric Oxide Production Assay
4.8. Cell Viability Assay
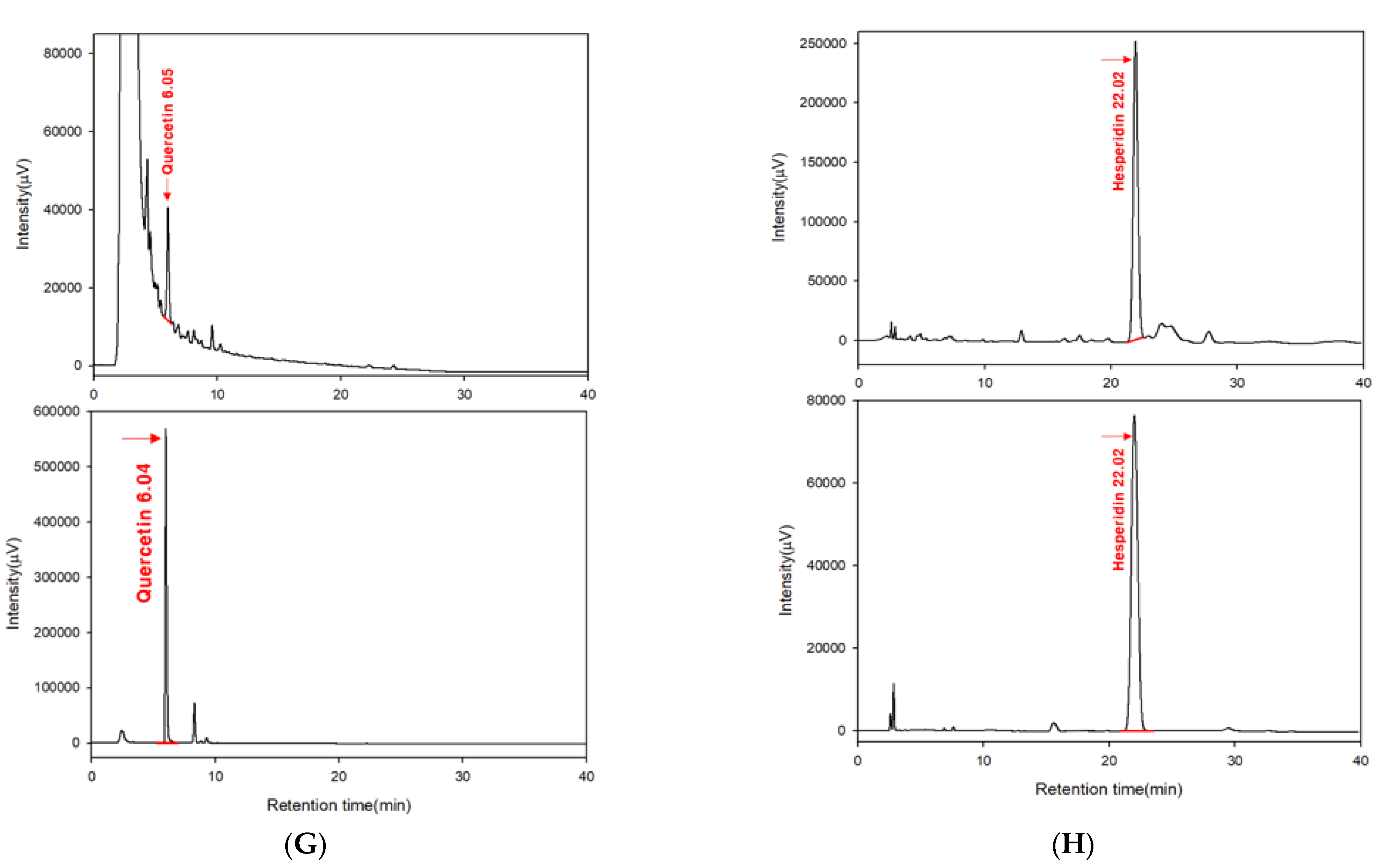
4.9. High-Performance Liquid Chromatography (HPLC)
4.10. Plasmid Transfection and Luciferase Reporter Assay
4.11. mRNA Analysis by a Reverse Transcription Polymerase Chain Reaction
4.12. Preparation of Total Cell and Nuclear Lysates
4.13. Immunoblotting
4.14. Immunoprecipitation
4.15. HCl/EtOH-Induced Gastritis
4.16. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AP-1 | Activator protein 1 |
| COX-2 | Cyclooxygenease-2 |
| HPLC | High-performance liquid chromatography |
| iNOS | Inducible nitric oxide synthase |
| IL-1β | Interleukin 1β |
| IL-6 | Interleukin 6 |
| JAK-STAT | Janus kinase-signal transducer and activator of transcription |
| LPS | Lipopolysaccharides |
| Mb-ME | Malus baccata methanol extract |
| MyD88 | Myeloid differentiation primary response 88 |
| MMP9 | Matrix metallopeptidase 9 |
| NF-κB | Nuclear factor-κB |
| Na-CMC | Sodium carboxymethyl cellulose |
| PRR | Pattern recognition receptor |
| pp85 | Phospho-PI3 kinase p85 |
| RT-PCR | Reverse transcription polymerase chain reaction |
| Src | Non-receptor tyrosine kinase |
| Syk | Spleen tyrosine kinase |
| TLR | Toll-like receptors |
| TRIF | TIR-domain-containing adapter-inducing interferon-β |
| TIRAP | TIR domain containing adaptor protein |
| TRAM | TRIF-related adaptor molecule |
| TNF-α | Tumor necrosis factor α |
| TAK1 | Transforming growth factor beta-kinase 1 |
References
- Chang, Y.L.; Liao, Y.W.; Chen, M.H.; Chang, S.Y.; Huang, Y.T.; Ho, B.C.; Yu, S.L. IKKε isoform switching governs the immune response against EV71 infection. Commun. Biol. 2021, 4, 663. [Google Scholar] [CrossRef] [PubMed]
- Garibotto, G.; Carta, A.; Picciotto, D.; Viazzi, F.; Verzola, D. Toll-like receptor-4 signaling mediates inflammation and tissue injury in diabetic nephropathy. J. Nephrol. 2017, 30, 719–727. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Bruneau, R.C.; Brennan, G.; Rothenburg, S. Battle Royale: Innate recognition of poxviruses and viral immune evasion. Biomedicines 2021, 9, 765. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.; Srivastava, M.; Saqib, U.; Liu, D.; Faisal, S.M.; Sugathan, S.; Bishnoi, S.; Baig, M.S. Potential therapeutic targets for inflammation in toll-like receptor 4 (TLR4)-mediated signaling pathways. Int. Immunopharmacol. 2016, 40, 79–89. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, L.A.J.; Bowie, A.G. The family of five: TIR-domain-containing adaptors in Toll-like receptor signalling. Nat. Rev. Immunol. 2007, 7, 353–364. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.J.; Jung, J.I.; Cho, H.J.; Choi, M.-S.; Sung, M.-K.; Yu, R.; Kang, Y.-H.; Park, J.H.Y. Berteroin present in cruciferous vegetables exerts potent anti-inflammatory properties in murine macrophages and mouse skin. Int. J. Mol. Sci. 2014, 15, 20686–20705. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204. [Google Scholar] [CrossRef] [Green Version]
- Yeung, Y.T.; Aziz, F.; Guerrero-Castilla, A.; Arguelles, S. Signaling pathways in inflammation and anti-inflammatory therapies. Curr. Pharm. Des. 2018, 24, 1449–1484. [Google Scholar] [CrossRef]
- Hemmati, S.; Haque, T.; Gritsman, K. Inflammatory signaling pathways in preleukemic and leukemic stem cells. Front. Oncol. 2017, 7, 265. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Liang, H.; Zen, K. Molecular mechanisms that influence the macrophage M1–M2 polarization balance. Front. Immunol. 2014, 5, 614. [Google Scholar] [CrossRef] [Green Version]
- Ahuja, A.; Kim, J.H.; Kim, J.H.; Yi, Y.S.; Cho, J.Y. Functional role of ginseng-derived compounds in cancer. J. Ginseng Res. 2018, 42, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Yi, Y.S.; Kim, M.Y.; Cho, J.Y. Role of ginsenosides, the main active components of Panax ginseng, in inflammatory responses and diseases. J. Ginseng Res. 2017, 41, 435–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, Q.; He, N.; Wang, Z.; Fu, X.; Aung, L.H.H.; Liu, Y.; Li, M.; Cho, J.Y.; Yang, Y.; Yu, T. Functional roles and mechanisms of ginsenosides from Panax ginseng in atherosclerosis. J. Ginseng Res. 2021, 45, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Lorz, L.R.; Kim, M.Y.; Cho, J.Y. Medicinal potential of Panax ginseng and its ginsenosides in atopic dermatitis treatment. J. Ginseng Res. 2020, 44, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Dadwal, V.; Agrawal, H.; Sonkhla, K.; Joshi, R.; Gupta, M. Characterization of phenolics, amino acids, fatty acids and antioxidant activity in pulp and seeds of high altitude Himalayan crab apple fruits (Malus baccata). J. Food Sci. Technol. 2018, 55, 2160–2169. [Google Scholar] [CrossRef]
- Sharma, R.; Nath, A.K. Antioxidant levels and activities of reactive oxygen-scavenging enzymes in crab apple fruits (Malus baccata). Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2016, 86, 877–885. [Google Scholar] [CrossRef]
- Mulabagal, V.; Van Nocker, S.; Dewitt, D.L.; Nair, M.G. Cultivars of apple fruits that are not marketed with potential for anthocyanin production. J. Agric. Food Chem. 2007, 55, 8165–8169. [Google Scholar] [CrossRef]
- Wei, X.; Zhao, R.; Sun, Y.-H.; Cong, J.-P.; Meng, F.-G.; Zhou, H.-M. The leaf extract of Siberian Crabapple (Malus baccata (Linn.) Borkh) contains potential fatty acid synthase inhibitors. J. Enzym. Inhib. Med. Chem. 2009, 24, 234–240. [Google Scholar] [CrossRef]
- Wang, L.; Li, X.; Wang, Z. Whole body radioprotective effect of phenolic extracts from the fruits of Malus baccata (Linn.) Borkh. Food Funct. 2016, 7, 975–981. [Google Scholar] [CrossRef]
- Yoshizawa, Y.; Sakurai, K.; Kawaii, S.; Soejima, J.; Murofushi, N. Antiproliferative and antioxidant properties of crabapple juices. Food Sci. Technol. Res. 2004, 10, 278–281. [Google Scholar] [CrossRef] [Green Version]
- Sharma, J.N.; Al-Omran, A.; Parvathy, S.S. Role of nitric oxide in inflammatory diseases. Inflammopharmacology 2007, 15, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Yang, W.S.; Yu, T.; Sung, G.-H.; Park, K.W.; Yoon, K.; Son, Y.-J.; Hwang, H.; Kwak, Y.-S.; Lee, C.-M.; et al. ATF-2/CREB/IRF-3-targeted anti-inflammatory activity of Korean red ginseng water extract. J. Ethnopharmacol. 2014, 154, 218–228. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.A.; Lee, C.Y.; Mitra, A.; Kim, H.; Woo, B.Y.; Hong, Y.D.; Noh, J.K.; Yi, D.K.; Kim, H.G.; Cho, J.Y. Anti-inflammatory effects of Huberia peruviana Cogn. methanol extract by inhibiting Src activity in the NF-kappaB pathway. Plants 2021, 10, 2335. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.A.; Oh, J.; Choi, S.R.; Lee, C.H.; Lee, B.H.; Lee, M.N.; Hossain, M.A.; Kim, J.H.; Lee, S.; Cho, J.Y. Anti-gastritis and anti-lung injury effects of pine tree ethanol extract targeting both NF-kappaB and AP-1 pathways. Molecules 2021, 26, 6275. [Google Scholar] [CrossRef]
- Jo, M.; Lee, J.; Kim, H.G.; Kim, J.K.; Kim, H.; Shin, K.K.; Bach, T.T.; Eum, S.M.; Lee, J.S.; Choung, E.S.; et al. Anti-inflammatory effect of Barringtonia angusta methanol extract is mediated by targeting of Src in the NF-kappaB signalling pathway. Pharm. Biol. 2021, 59, 799–810. [Google Scholar] [CrossRef]
- Korhonen, R.; Lahti, A.; Kankaanranta, H.; Moilanen, E. Nitric oxide production and signaling in inflammation. Curr. Drug Targets Inflamm. Allergy 2005, 4, 471–479. [Google Scholar] [CrossRef]
- Liu, Y.; Krueger, J.G.; Bowcock, A.M. Psoriasis: Genetic associations and immune system changes. Genes Immun. 2007, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Sisto, M.; Ribatti, D.; Lisi, S. Understanding the complexity of Sjögren’s syndrome: Remarkable progress in elucidating NF-κB mechanisms. J. Clin. Med. 2020, 9, 2821. [Google Scholar] [CrossRef]
- Lawrence, T. The nuclear factor NF-kappaB pathway in inflammation. Cold Spring Harb. Perspect. Biol. 2009, 1, a001651. [Google Scholar] [CrossRef] [Green Version]
- Hoesel, B.; Schmid, J.A. The complexity of NF-κB signaling in inflammation and cancer. Mol. Cancer 2013, 12, 86. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.S.; Jeong, D.; Nam, G.; Yi, Y.-S.; Yoon, D.H.; Kim, T.W.; Park, Y.C.; Hwang, H.; Rhee, M.H.; Hong, S.; et al. AP-1 pathway-targeted inhibition of inflammatory responses in LPS-treated macrophages and EtOH/HCl-treated stomach by Archidendron clypearia methanol extract. J. Ethnopharmacol. 2013, 146, 637–644. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.G.; Yang, W.S.; Sung, G.H.; Kim, J.H.; Baek, G.S.; Kim, E.; Yang, S.; Park, Y.C.; Sung, J.M.; Yoon, D.H.; et al. IKK β -targeted anti-inflammatory activities of a butanol fraction of artificially cultivated Cordyceps pruinosa fruit bodies. Evid.-Based Complement. Altern. Med. 2014, 2014, 562467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.G.; Son, Y.-J.; Kim, M.-Y.; Cho, J.Y. Syk and IRAK1 contribute to immunopharmacological activities of anthraquinone-2-carboxlic acid. Molecules 2016, 21, 809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, W.S.; Jeong, D.; Yi, Y.-S.; Lee, B.-H.; Kim, T.W.; Htwe, K.M.; Kim, Y.-D.; Yoon, K.D.; Hong, S.; Lee, W.-S.; et al. Myrsine seguinii ethanolic extract and its active component quercetin inhibit macrophage activation and peritonitis induced by LPS by targeting to Syk/Src/IRAK-1. J. Ethnopharmacol. 2014, 151, 1165–1174. [Google Scholar] [CrossRef]
- Jeong, H.Y.; Sung, G.-H.; Kim, J.H.; Yoon, J.Y.; Yang, Y.; Park, J.G.; Kim, S.H.; Yi, Y.-S.; Yang, W.S.; Yoon, D.H.; et al. Syk and Src are major pharmacological targets of a Cerbera manghas methanol extract with kaempferol-based anti-inflammatory activity. J. Ethnopharmacol. 2014, 151, 960–969. [Google Scholar] [CrossRef]
- Hong, Y.H.; Kim, J.H.; Cho, J.Y. Ranunculus bulumei methanol extract exerts anti-inflammatory activity by targeting Src/Syk in NF-κB signaling. Biomolecules 2020, 10, 546. [Google Scholar] [CrossRef] [Green Version]
- Yi, Y.-S.; Cho, J.Y.; Kim, D. Cerbera manghas methanol extract exerts anti-inflammatory activity by targeting c-Jun N-terminal kinase in the AP-1 pathway. J. Ethnopharmacol. 2016, 193, 387–396. [Google Scholar] [CrossRef]
- González, M.I.; Martín-Duque, P.; Desco, M.; Salinas, B. Radioactive labeling of milk-derived xxosomes with 99mTc and in vivo tracking by SPECT imaging. Nanomaterials 2020, 10, 1062. [Google Scholar] [CrossRef]
- Li, Z.-H.; Guo, H.; Xu, W.-B.; Ge, J.; Li, X.; Alimu, M.; He, D.-J. Rapid identification of flavonoid constituents directly from PTP1B inhibitive extract of raspberry (Rubus idaeus L.) leaves by HPLC-ESI-QTOF-MS-MS. J. Chromatogr. Sci. 2016, 54, 805–810. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.G.; Kim, M.-Y.; Cho, J.Y. Alisma canaliculatum ethanol extract suppresses inflammatory responses in LPS-stimulated macrophages, HCl/EtOH-induced gastritis, and DSS-triggered colitis by targeting Src/Syk and TAK1 activities. J. Ethnopharmacol. 2018, 219, 202–212. [Google Scholar] [CrossRef]
- Qomaladewi, N.P.; Aziz, N.; Kim, M.Y.; Cho, J.Y. Piper cubeba L. methanol extract has anti-inflammatory activity targeting Src/Syk via NF-κB inhibition. Evid.-Based Complement. Altern. Med. 2019, 2019, 1548125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.-O.; Yang, W.S.; Park, J.G.; Jeong, D.; Kim, H.G.; Yoon, K.D.; Aravinthan, A.; Kim, J.-H.; Kim, E.; Cho, J.Y. Src and Syk contribute to the anti-inflammatory activities of Achyranthes aspera ethanolic extract. J. Ethnopharmacol. 2017, 206, 1–7. [Google Scholar] [CrossRef]
- Liu, T.; Li, X.; Wu, M.; Qin, L.; Chen, H.; Qian, P. Seneca valley virus 2C and 3C(pro) induce apoptosis via mitochondrion-mediated intrinsic pathway. Front. Microbiol. 2019, 10, 1202. [Google Scholar] [CrossRef]
- Mishra, M.; Tiwari, S.; Gomes, A.V. Protein purification and analysis: Next generation Western blotting techniques. Expert Rev. Proteom. 2017, 14, 1037–1053. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Yang, W.S.; Yu, T.; Yi, Y.-S.; Park, J.G.; Jeong, D.; Kim, J.H.; Oh, J.S.; Yoon, K.; Kim, J.-H.; et al. Novel anti-inflammatory function of NSC95397 by the suppression of multiple kinases. Biochem. Pharmacol. 2014, 88, 201–215. [Google Scholar] [CrossRef] [PubMed]
- Seok Yang, W.; Lee, J.; Woong Kim, T.; Hye Kim, J.; Lee, S.; Hee Rhee, M.; Hong, S.; Youl Cho, J. Src/NF-κB-targeted inhibition of LPS-induced macrophage activation and dextran sodium sulphate-induced colitis by Archidendron clypearia methanol extract. J. Ethnopharmacol. 2012, 142, 287–293. [Google Scholar] [CrossRef] [PubMed]
- Shin, K.K.; Park, J.G.; Hong, Y.H.; Aziz, N.; Park, S.H.; Kim, S.; Kim, E.; Cho, J.Y. Anti-inflammatory effects of Licania macrocarpa Cuatrec methanol extract target Src- and TAK1-mediated pathways. Evid.-Based Complement. Altern. Med. 2019, 2019, 4873870. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Analyzing Compound | Quercetin | Hesperidin |
|---|---|---|
| Column | CAPCELL PAK C18 MG, 4.6 mm I.D. × 250 mm | CAPCELL PAK C18 MG, 4.6 mm I.D. × 250 mm |
| Wavelength (nm) | 254 | 285 |
| Mobile phase | A: 2% acetic acid in water B: 0.5% acetic acid in water: CAN = 50:50 | Methanol: water of acetic acid (pH 3) = 30:70 |
| Gradient or isocratic conditions | 0: A (28%), B (72%) 20 min: B (100%) 30 min: B (100%) | 40 min: 100% |
| Oven temperature (°C) | 35 | 50 |
| Name | Primer | Sequence (5′ to 3′) |
|---|---|---|
| iNOS | Forward | GTGAAGAAAACCCCTTGTGCTG |
| Reverse | AGTTCCGAGCGTCAAAGACC | |
| IL-1β | Forward | CAGGATGAGGACATGAGCACC |
| Reverse | CTCTGCAGACTCAAACTCCAC | |
| IL-6 | Forward | GCCTTCTTGGGACTGATGCT |
| Reverse | TGGAAATTGGGGTAGGAAGGAC | |
| TNF-α | Forward | TTGACCTCAGCCGTGAGTTG |
| Reverse | CCTGTAGCCCACGTCGTAGC | |
| MMP-9 | Forward | TCTTCCCCAAAGACCTGAAA |
| Reverse | TGATGTTATGATGGTCCCAC | |
| COX-2 | Forward | CACTACATCCTGACCCACTT |
| Reverse | ATGCTCCTGCTTGAGTATGT | |
| GADPH | Forward | ACCACAGTCCATGCCATCAC |
| Reverse | CCACCACCCTGTTGCTGTAG |
| Name | Primer | Sequence (5′ to 3′) |
|---|---|---|
| TNF-α | Forward | TGCCTATGTCTCAGCCTCTT |
| Reverse | GAGGCCATTTGGGAACTTCT | |
| iNOS | Forward | CGAAACGCTTCACTTCCAA |
| Reverse | TGAGCCTATATTGCTGTGGCT | |
| IL-6 | Forward | GTCCTTCCTACCCCAATTTCCA |
| Reverse | TAACGCACTAGGTTTGCCGA | |
| GADPH | Forward | GGGTCCCAGCTTAGGTTCATC |
| Reverse | TACGGCCAAATCCGTTCACA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, C.; Chen, H.; Kim, S.A.; Lee, J.S.; Choung, E.S.; Zhang, Z.; Kim, S.-Y.; Kim, J.-H.; Cho, J.Y. Anti-Inflammatory Functions of Methanol Extract from Malus baccata (L.) Borkh. Leaves and Shoots by Targeting the NF-κB Pathway. Plants 2022, 11, 646. https://doi.org/10.3390/plants11050646
Song C, Chen H, Kim SA, Lee JS, Choung ES, Zhang Z, Kim S-Y, Kim J-H, Cho JY. Anti-Inflammatory Functions of Methanol Extract from Malus baccata (L.) Borkh. Leaves and Shoots by Targeting the NF-κB Pathway. Plants. 2022; 11(5):646. https://doi.org/10.3390/plants11050646
Chicago/Turabian StyleSong, Chaoran, Hongxi Chen, Soo Ah Kim, Jong Sub Lee, Eui Su Choung, Zhiyun Zhang, Soo-Yong Kim, Jong-Hoon Kim, and Jae Youl Cho. 2022. "Anti-Inflammatory Functions of Methanol Extract from Malus baccata (L.) Borkh. Leaves and Shoots by Targeting the NF-κB Pathway" Plants 11, no. 5: 646. https://doi.org/10.3390/plants11050646