
The Fruit Proteome Response to the Ripening Stages in Three Tomato Genotypes


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Protein Extraction
2.3. Protein Quantitation
2.4. Two-Dimensional Electrophoresis (2-DE)
2.5. Peptide Extraction
2.6. Protein Identification
2.7. Statistical Analysis
3. Results
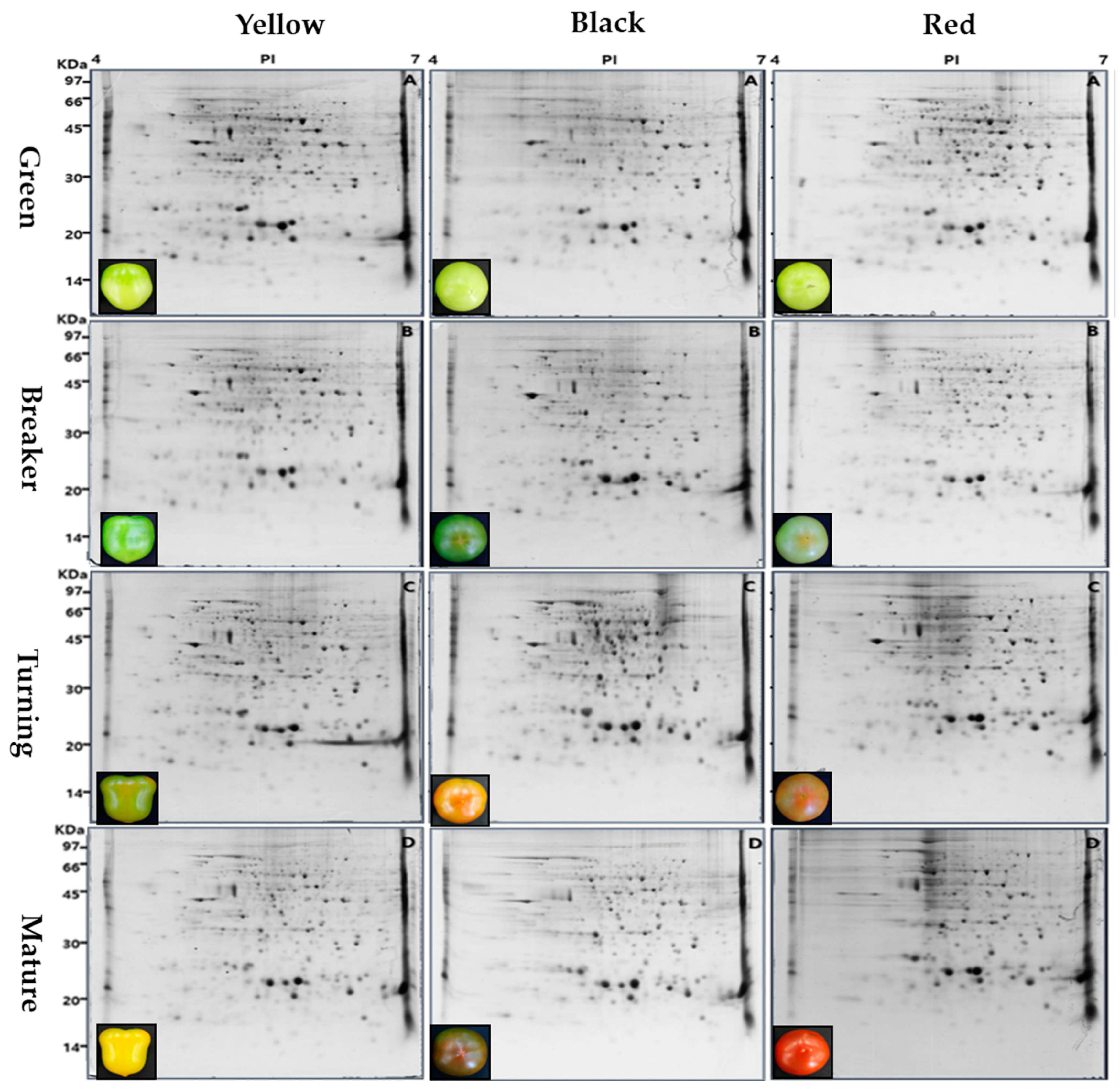
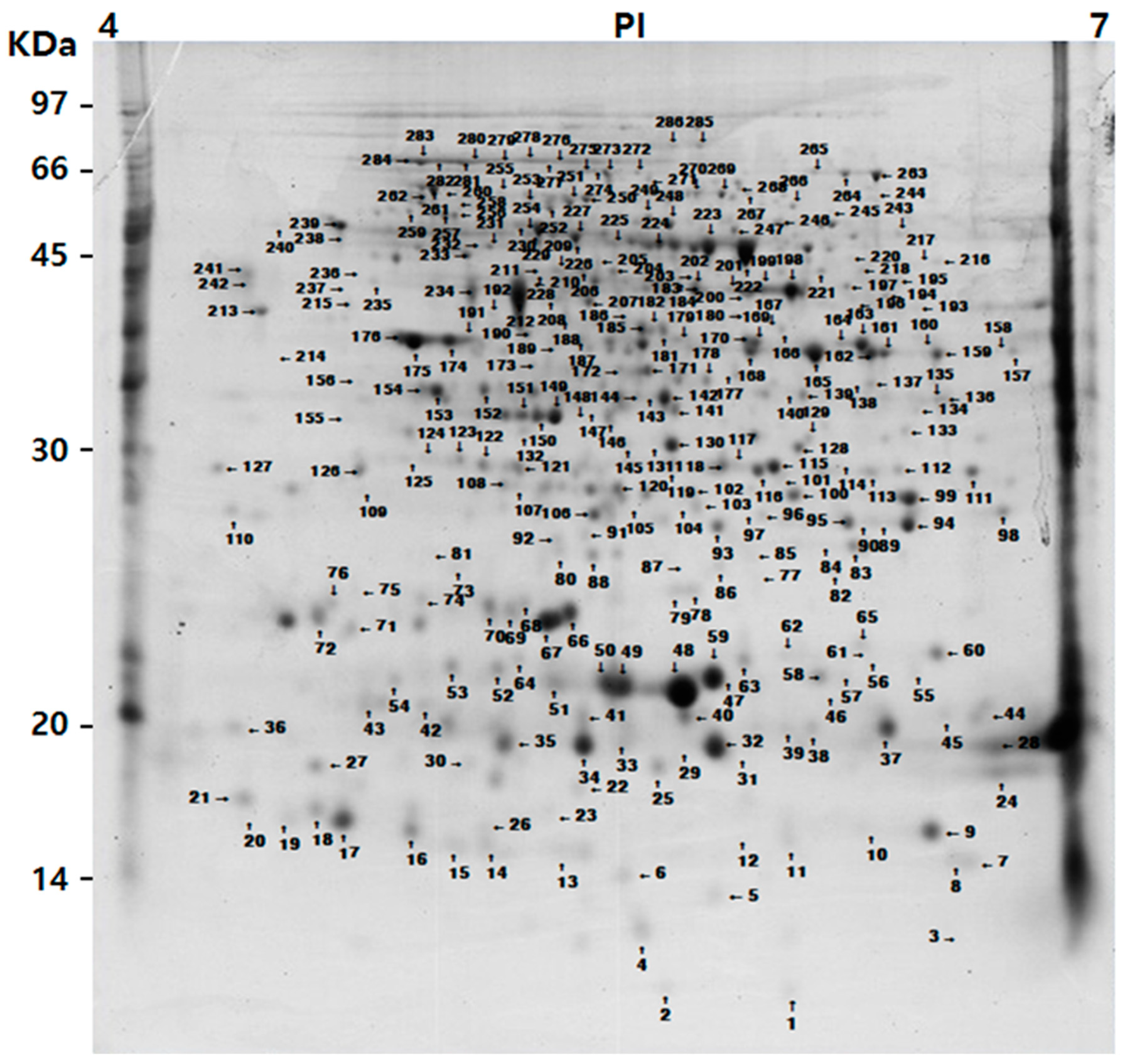
3.1. 2-DE Gel Imagery and a Matched Set of Spots in Different Tomato Cultivars
3.2. Classified and Identified Protein Spots
3.3. Differential Protein Expression Abundance According to Genotypes and Ripening Stages
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Choi:, S.H.; Kim, D.S.; Kozukue, N.; Kin, H.J.; Nishitani, Y.; Mizuno, M.; Levin, C.E.; Friedman, M. Protein, free amino acid, phenolic, b-carotene, and lycopene content, and antioxidative and cancer cell inhibitory effects of 12 greenhouse-grown commercial cherry tomato varieties. J. Food Compos. Anal. 2014, 34, 115–127. [Google Scholar] [CrossRef]
- Vela-Hinojosa, C.; Escalona-Buendía, H.B.; Mendoza-Espinoza, J.A.; Villa-Hernández, J.M.; Lobato-Ortíz, R.; Rodríguez-Pérez, J.E.; Pérez-Flores, L.J. Antioxidant balance and regulation in tomato genotypes of different color. J. Am. Soc. Hortic. Sci. 2019, 144, 45–54. [Google Scholar] [CrossRef] [Green Version]
- Yao, G.F.; Wei, Z.Z.; Li, T.T.; Tang, J.; Huang, Z.Q.; Yang, F.; Li, Y.H.; Han, Z.; Hu, F.; Hu, L.Y.; et al. Modulation of enhanced antioxidant activity by hydrogen sulfide antagonization of ethylene in tomato fruit ripening. J. Agric. Food Chem. 2018, 66, 10380–10387. [Google Scholar] [CrossRef]
- El-Bendary, N.; Hariri, E.; Hassainen, A.E.; Badr, A. Using machine learning techniques for evaluating tomato ripeness. Expert Syst. Appl. 2015, 42, 1892–1905. [Google Scholar] [CrossRef]
- Garcia, M.B.; Ambat, S.; Adao, R.T. Tomato, tomahto: A machine learning approach for tomato ripening stage identification using pixel-based color image classification. In Proceedings of the 2019 IEEE 11th International Conference on Humanoid, Nanotechnology, Information Technology, Communication and Control, Environment, and Management (HNICEM), Laoag, Philippines, 29 November–1 December 2019; IEEE: Piscataway, NJ, USA, 2019; pp. 1–6. [Google Scholar]
- Xiao, Y.; Kang, B.; Li, M.; Xiao, L.; Xiao, H.; Shen, H. Transcription of lncRNA ACoS-AS1 is essential to trans-splicing between SlPsy1 and ACoS-AS1 that causes yellow fruit in tomato. RNA Bio 2020, 17, 596–607. [Google Scholar] [CrossRef]
- Park, M.H.; Sangwanangkul, P.; Baek, D.R. Changes in carotenoid and chlorophyll content of black tomatoes (Lycopersicone sculentum L.) during storage at various temperatures. Saudi J. Biol. Sci 2018, 25, 57–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blando, F.; Berland, H.; Majorano, G.; Durante, M.; Mazzucato, A.; Picarella, M.E.; Nicoletti, I.; Gerardi, C.; Mita, G.; Andersen, Ø.M. Nutraceutical characterization of anthocyanin-rich fruits produced by “Sun Black” tomato line. Front. Nutr. 2019, 6, 133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotikova, Z.; Hejtmankova, A.; Lachman, J. Determination of the influence of variety and level of maturity on the content and development of carotenoids in tomatoes. Czech J. Food Sci. Spec. Issue 2009, 27, 200–203. [Google Scholar] [CrossRef] [Green Version]
- Yoo, H.J.; Park, W.J.; Lee, G.M.; Oh, C.S.; Yeam, I.; Won, D.C.; Kim, C.K.; Lee, J.M. Inferring the genetic determinants of fruit colors in tomato by carotenoid profiling. Molecules 2017, 22, 764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.; Yang, X.; Yu, Y.; Si, X.; Zhai, X.; Zhang, H.; Dong, W.; Gao, C.; Xu, C. Domestication of wild tomato is accelerated by genome editing. Nat. Biotechnol. 2018, 36, 1160–1163. [Google Scholar] [CrossRef]
- McQuinn, R.P.; Gapper, N.E.; Gray, A.G.; Zhong, S.; Tohge, T.; Fei, Z.; Fernie, A.; Giovannoni, J.J. Manipulation of ZDS in tomato exposes carotenoid and ABA specific effects on fruit development and ripening. Plant Biotechnol. J. 2020, 18, 2210–2224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bizouerne, E.; Buitink, J.; Vu, B.L.; Vu, J.L.; Esteban, E.; Pasha, A.; Provart, N.; Verdier, J.; Leprince, O. Gene co-expression analysis of tomato seed maturation reveals tissue-specific regulatory networks and hubs associated with the acquisition of desiccation tolerance and seed vigor. BMC Plant Biol. 2021, 21, 124. [Google Scholar] [CrossRef] [PubMed]
- Batista-Silva, W.; Nascimento, V.L.; Medeiros, D.B.; Nunes-Nesi1, A.; Ribeiro, D.M.; Zsögön, A.; Araújo, W.L. Modifications in organic acid profiles during fruit development and ripening: Correlation or causation? Front. Plant Sci. 2018, 9, 1689. [Google Scholar] [CrossRef] [PubMed]
- Bertero, V.G.; Pratta, G.R.; Arce, D.P. Independent transcriptomic and proteomic networks reveal common differentially expressed chaperone and interactor genes during tomato cv. Micro-Tom fruit ripening. Plant Gene 2021, 28, 100346. [Google Scholar] [CrossRef]
- Gao, Y.; Wei, W.; Zhao, X.; Tan, X.; Fan, Z.; Zhang, Y.; Jing, Y.; Meng, L.; Zhu, B.; Zhu, H.; et al. A NAC transcription factor, NOR-like 1, is a new positive regulator of tomato fruit ripening. Hortic. Res. 2018, 5, 75. [Google Scholar] [CrossRef] [Green Version]
- Quinet, M.; Angosto, T.; Yuste-Lisbona, F.J.; Blanchard-Gros, R.; Bigot, S.; Marinez, J.P.; Lutts, S. Tomato fruit development and metabolism. Front. Plant Sci. 2019, 10, 1554. [Google Scholar] [CrossRef] [Green Version]
- D’Ambrosio, C.; Arena, S.; Rocco, M.; Verrillo, F.; Novi, G.; Viscosi, V.; Marra, M.; Scaloni, A. Proteomic analysis of apricot fruit during ripening. J. Proteom. 2013, 78, 39–57. [Google Scholar] [CrossRef] [Green Version]
- Salzano, A.M.; Renzone, G.; Sobolev, A.P.; Carbone, V.; Petriccione, M.; Capitani, D.; Vitale, M.; Novi, G.; Zambrano, N.; Pasquariello, M.S.; et al. Unveiling kiwifruit metabolite and protein changes in the course of postharvest cold storage. Front. Plant Sci. 2019, 10, 71. [Google Scholar] [CrossRef] [Green Version]
- Mouzo, D.; Bernal, J.; López-Pedrouso, M.; Franco, D.; Zapata, C. Advances in the biology of seed and vegetative storage proteins based on two-dimensional electrophoresis coupled to mass spectrometry. Molecules 2018, 23, 2462. [Google Scholar] [CrossRef] [Green Version]
- Kok, S.Y.; Namasivayam, P.; Ee, G.C.L.; Ong-Abdullah, M. Comparative proteomic analysis of oil palm (Elaeis guineensis Jacq.) during early fruit development. J. Proteom. 2021, 232, 104052. [Google Scholar] [CrossRef]
- Deytieux, C.; Geny, L.; Lapaillerie, D.; Claverol, S.; Bonneu, M.; Donèche, B. Proteome analysis of grape skins during ripening. J. Exp. Bot. 2007, 58, 1851–1862. [Google Scholar] [CrossRef] [PubMed]
- Tian, B.; Harrison, R.; Morton, J.; Jaspers, M. Influence of skin contact and different extractants on the extraction of proteins and phenolic substances in Sauvignon Blanc grape skin. Aust. J. Grape Wine Res. 2020, 26, 180–187. [Google Scholar] [CrossRef]
- Sarry, J.E.; Sommerer, N.; Sauvage, F.X.; Bergoin, A.; Rossignol, M.; Albagnac, G.; Romieu, C. Grape berry biochemistry revisited upon proteomic analysis of the mesocarp. Proteomic 2004, 4, 201–215. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Lüscher, J.; Plank, C.M.; Brillante, L.; Cooper, M.L.; Smith, R.J.; Al-Rwahnih, M.; Yu, R.; Oberholster, A.; Girardello, R.; Kurtural, S.K. Grapevine red blotch virus may reduce carbon translocation leading to impaired grape berry ripening. J. Agric. Food Chem. 2019, 67, 2437–2448. [Google Scholar] [CrossRef]
- Faurobert, M.; Mihr, C.; Bertin, N.; Pawlowski, T.; Negroni, L.; Sommerer, N.; Causse, M. Major proteome variations associated with cherry tomato pericarp development and ripening. Plant Physiol. 2007, 143, 1327–1346. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Xu, Z.; Ding, A.; Kong, Y. Genome-wide identification and expression profiling analysis of the xyloglucan endotransglucosylase/hydrolase gene family in tobacco (Nicotiana tabacum L.). Genes 2018, 9, 273. [Google Scholar] [CrossRef] [Green Version]
- Mazzeo, M.F.; Cacace, G.; Iovieno, P.; Massarelli, I.; Grillo, S.; Siciliano, R.A. Response mechanisms induced by exposure to high temperature in anthers from thermo-tolerant and thermo-sensitive tomato plants: A proteomic perspective. PLoS ONE 2018, 13, e0201027. [Google Scholar] [CrossRef] [Green Version]
- Efrat, A.; Hoffmann, F.; Kriegel, K.; Schultz, C.; Wenk, C. Geometric algorithms for the analysis of 2D-electrophoresis gels. J. Comput. Biol. 2002, 9, 299–315. [Google Scholar] [CrossRef]
- Carrari, F.; Fernie, A.R. Metabolic regulation underlying tomato fruit development. J. Exp. Bot. 2006, 57, 1883–1897. [Google Scholar] [CrossRef]
- Pontiggia, D.; Spinelli, F.; Fabbri, C.; Licursi, V.; Negri, R.; Lorenzo, G.D.; Mattei, B. Changes in the microsomal proteome of tomato fruit during ripening. Sci. Rep. 2019, 9, 14350. [Google Scholar] [CrossRef]
- Alba, R.; Payton, P.; Fei, Z.J.; McQuinn, R.; Debbie, P.; Martin, G.B.; Tanksley, S.D.; Giovannoni, J.J. Transcriptome and selected metabolite analyses reveal multiple points of ethylene control during tomato fruit development. Plant Cell 2005, 17, 2954–2965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocco, M.; Ambrosio, C.D.; Arena, S.; Faurobert, M.; Scaloni, A.; Marra, M. Proteomic analysis of tomato fruits from two ecotypes during ripening. Proteomics 2006, 6, 3781–3791. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Ge, C.; Yang, M.; Long, Y.; Li, M.; Zhang, Y.; Chen, Q.; Sun, B.; Wang, Y.; Wang, X.; et al. Cytosolic/plastid glyceraldehyde-3-phosphate dehydrogenase is a negative regulator of strawberry fruit ripening. Gens 2020, 11, 580. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.; Song, J.; Campbell-Palmer, L.; Thompson, K.; Li, L.; Walker, B.; Cui, Y.; Li, X. A proteomic investigation of apple fruit during ripening and in response to ethylene treatment. J. Proteom. 2013, 93, 276–294. [Google Scholar] [CrossRef] [PubMed]
- Sharpe, R.; Gustafson, L.; Hewitt, S.; Kilian, B.; Grabb, K.J.; Hendrickson, C.; Jiwan, D.; Andrew, P.; Dhingra, A. Concomitant phytonutrient and transcriptome analysis of mature fruit and leaf tissues of tomato (Solanum lycopersicum L. cv. Oregon Spring) grown using organic and conventional fertilizer. PLoS ONE 2020, 15, e0227429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sorrequieta, A.; Ferraro, G.; Boggio, S.B.; Valle, E.M. Free amino acid production during tomato fruit ripening: A focus on L-glutamate. Amino Acids 2010, 38, 1523–1532. [Google Scholar] [CrossRef]
- Zhao, X.; Yuan, X.; Chen, S.; Fu, D.Q.; Jiang, C.Z. Metabolomic and transcriptomic analyses reveal that a MADS-box transcription factor TDR4 regulates tomato fruit quality. Front. Plant Sci. 2019, 10, 792. [Google Scholar] [CrossRef] [Green Version]
- Ji, D.; Cui, X.; Qin, G.; Chen, T.; Tian, S. SlFERL interacts with S-adenosylmethionine synthetase to regulate fruit ripening. Plant Physiol. 2020, 184, 2168–2181. [Google Scholar] [CrossRef]
- Yuan, Y.; Xu, X.; Gong, Z.; Tang, Y.; Wu, M.; Yan, F.; Zhang, X.; Zhang, Q.; Yang, F.; Hu, X.; et al. Auxin response factor 6A regulates photosynthesis, sugar accumulation, and fruit development in tomato. Hortic. Res. 2019, 6, 85. [Google Scholar] [CrossRef] [Green Version]
- Tao, X.; Wu, Q.; Li, J.; Wang, D.; Nassarawa, S.S.; Ying, T. Ethylene biosynthesis and signal transduction are enhanced during accelerated ripening of postharvest tomato treated with exogenous methyl jasmonate. Sci. Hortic. 2021, 281, 109965. [Google Scholar] [CrossRef]
- Gao, H.; Zhu, H.; Chen, Y.S.; Lu, C.; Zhu, B.; Luo, Y. Lycopene accumulation affects the biosynthesis of some carotenoid-related volatiles independent of ethylene in tomato. J. Integr. Plant Biol. 2008, 50, 991–996. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Jiang, L.; Zhang, L.; Kang, R.; Yu, Z. Dynamic changes in proteins during apple (Malus x Domestica) fruit ripening and storage. Hortic. Res. 2014, 1, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luengwilai, K.; Saltveit, M.; Beckles, D.M. Metabolite content of harvested Micro-Tom tomato (Solanum lycopersicum L.) fruit is altered by chilling and protective heat-shock treatments as shown by GC–MS metabolic profiling. Postharvest Biol. Technol. 2012, 63, 116–122. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, H.; Wang, H.; Li, B.; Yi, Y.; Kong, F.; Liu, J.; Zhang, H. Constitutive expression of a tomato small heat shock protein gene LeHSP21 improves tolerance to high-temperature stress by enhancing antioxidation capacity in tobacco. Plant Mol. Biol. Rep. 2016, 34, 399–409. [Google Scholar] [CrossRef]
- Sanchez-Bel, P.; Egea, I.; Sanchez-Ballesta, M.T.; Sevillano, L.; Bolarin, M.D.C.; Flores, F.B. Proteome changes in tomato fruits prior to visible symptoms of chilling injury are linked to defensive mechanisms, uncoupling of photosynthetic processes and protein degradation machinery. Plant Cell Physiol. 2012, 53, 470–484. [Google Scholar] [CrossRef] [Green Version]
- Wahyudi, A.; Ariyani, D.; Ma, G.; Inaba, R.; Fukasawa, C.; Nakano, R.; Motohashi, R. Functional analyses of lipocalin proteins in tomato. Plant Biotechnol. 2018, 35, 303–312. [Google Scholar] [CrossRef] [Green Version]
- Tuan, P.A.; Lee, J.; Park, C.H.; Kim, J.K.; Noh, Y.H.; Kim, Y.B.; Kim, H.R.; Park, S.U. Carotenoid biosynthesis in oriental melon (Cucumis melo L. var. makuwa). Foods 2019, 8, 77. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar. (Fruit Color) | Ripening Stage z | Spot y | Matched Spot x | Matching Ratio (%) |
|---|---|---|---|---|
| Sugaryellow (Yellow) | Green | 351 | 269 | 76.6 |
| Breaker | 308 | 242 | 75.6 | |
| Turning | 401 | 317 | 79.1 | |
| Mature | 372 | 364 | 97.8 | |
| Sinheuksu (Black) | Green | 256 | 198 | 77.3 |
| Breaker | 287 | 286 | 99.7 | |
| Turning | 333 | 211 | 63.7 | |
| Mature | 311 | 201 | 64.6 | |
| Rapsodie (Red) | Green | 398 | 356 | 89.4 |
| Breaker | 399 | 313 | 78.4 | |
| Turning | 334 | 244 | 73.1 | |
| Mature | 257 | 198 | 77.0 |
| Pot No. z | Protein Identification y | ID (NCBI) | Mr/pI | MP x |
|---|---|---|---|---|
| Metabolism | ||||
| 187 | Glutamine synthase | Q42874 | 18,735/6.13 | 5 |
| 189 | Glutamine synthase | Q42874 | 18,735/6.13 | 5 |
| 200 | S-adenosylmethionine synthase 1 | P43280 | 43,730/5.52 | 11 |
| 204 | S-adenosylmethionine synthase 2 | P43281 | 43,511/5.41 | 11 |
| Energy | ||||
| 157 | Glyceraldehyde-3-phosphate dehydrogenase | O04891 | 32,097/5.93 | 10 |
| 159 | Glyceraldehyde-3-phosphate dehydrogenase | O04891 | 32,097/5.93 | 16 |
| 162 | Glyceraldehyde-3-phosphate dehydrogenase | O04891 | 32,097/5.93 | 16 |
| 168 | Glyceraldehyde-3-phosphate dehydrogenase | gi|525314339 | 32,100/5.93 | 7 |
| 264 | Glyceraldehyde-3-phosphate dehydrogenase | Q7Y0S2 | 28,957/5.70 | 8 |
| 193 | Phosphoglycerate kinase | K4CHY3 | 42,263/5.78 | 9 |
| 197 | Phosphoglycerate kinase, cytosolic-like | gi|460396822 | 39,627/6.49 | 8 |
| 202 | Phosphoglycerate kinase | K4CHY4 | 50,592/7.66 | 13 |
| Protein synthesis | ||||
| 279 | Heat shock protein 70 isoform 2 | H1ZXA8 | 71,142/5.08 | 19 |
| 280 | Heat shock protein 70 isoform 3 | H1ZXA9 | 71,744/5.14 | 19 |
| 281 | Heat shock protein 70 kDa protein 2-like | gi|460408280 | 71,141/5.08 | 18 |
| 282 | Heat shock protein 70 | A8W7B5 | 74,415/5.41 | 18 |
| 283 | Heat shock protein 70 | A8W7B5 | 74,415/5.41 | 16 |
| 284 | Heat shock protein 70 | A8W7B5 | 74,415/5.41 | 13 |
| Interaction with the cellular environment | ||||
| 10 | Temperature-induced lipocalin | Q38JD4 | 21,509/6.16 | 10 |
| 61 | Temperature-induced lipocalin | Q38JE1 | 21,303/5.96 | 5 |
| 65 | Temperature-induced lipocalin | Q38JE1 | 21,303/5.96 | 7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, H.-G.; Park, D.-Y.; Kang, N.-J. The Fruit Proteome Response to the Ripening Stages in Three Tomato Genotypes. Plants 2022, 11, 553. https://doi.org/10.3390/plants11040553
Choi H-G, Park D-Y, Kang N-J. The Fruit Proteome Response to the Ripening Stages in Three Tomato Genotypes. Plants. 2022; 11(4):553. https://doi.org/10.3390/plants11040553
Chicago/Turabian StyleChoi, Hyo-Gil, Dong-Young Park, and Nam-Jun Kang. 2022. "The Fruit Proteome Response to the Ripening Stages in Three Tomato Genotypes" Plants 11, no. 4: 553. https://doi.org/10.3390/plants11040553