
The Passage of H2O2 from Chloroplasts to Their Associated Nucleus during Retrograde Signalling: Reflections on the Role of the Nuclear Envelope


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
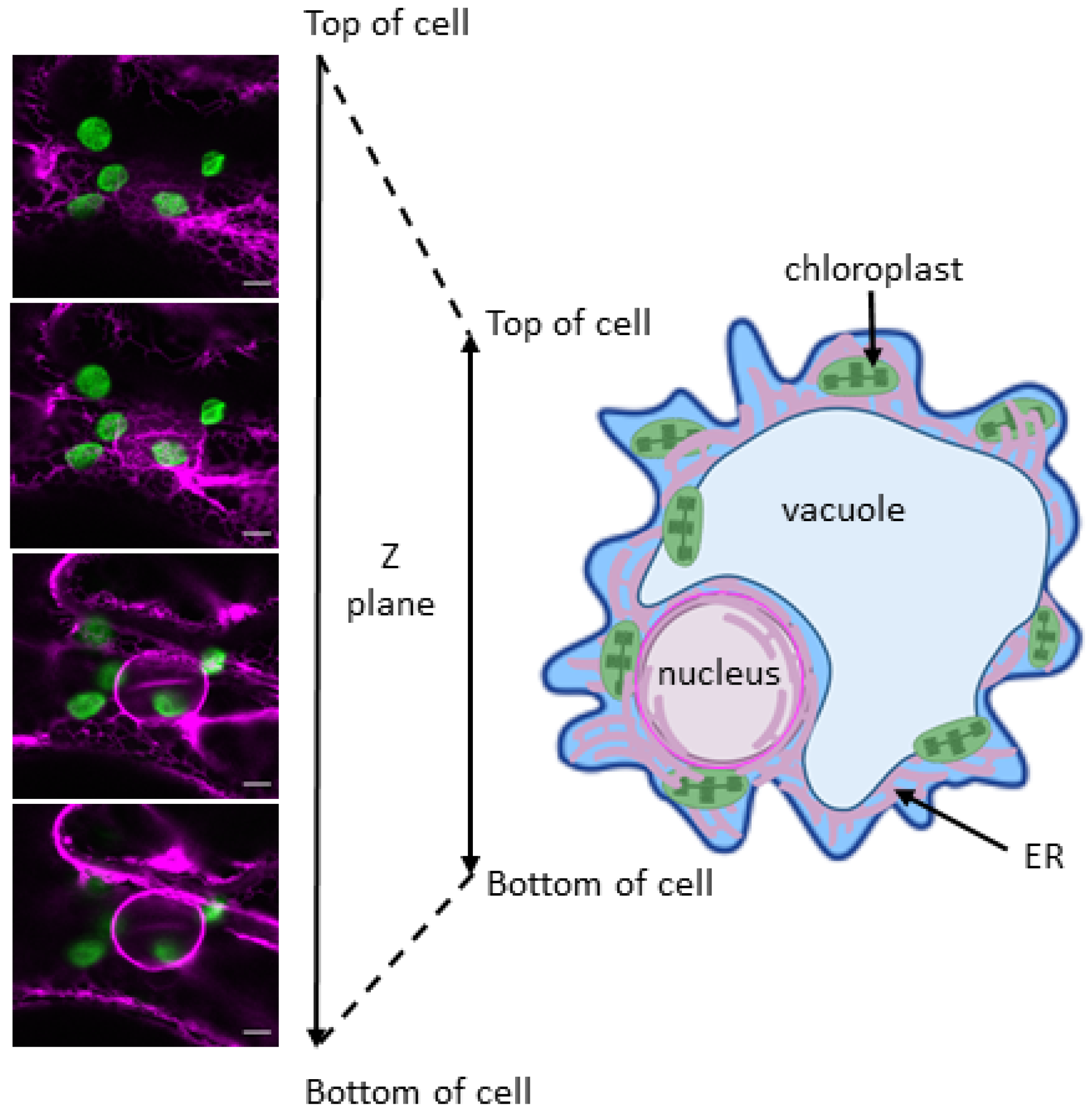
2. Stromules
3. Nature of the Linkages—The Nuclear Envelope
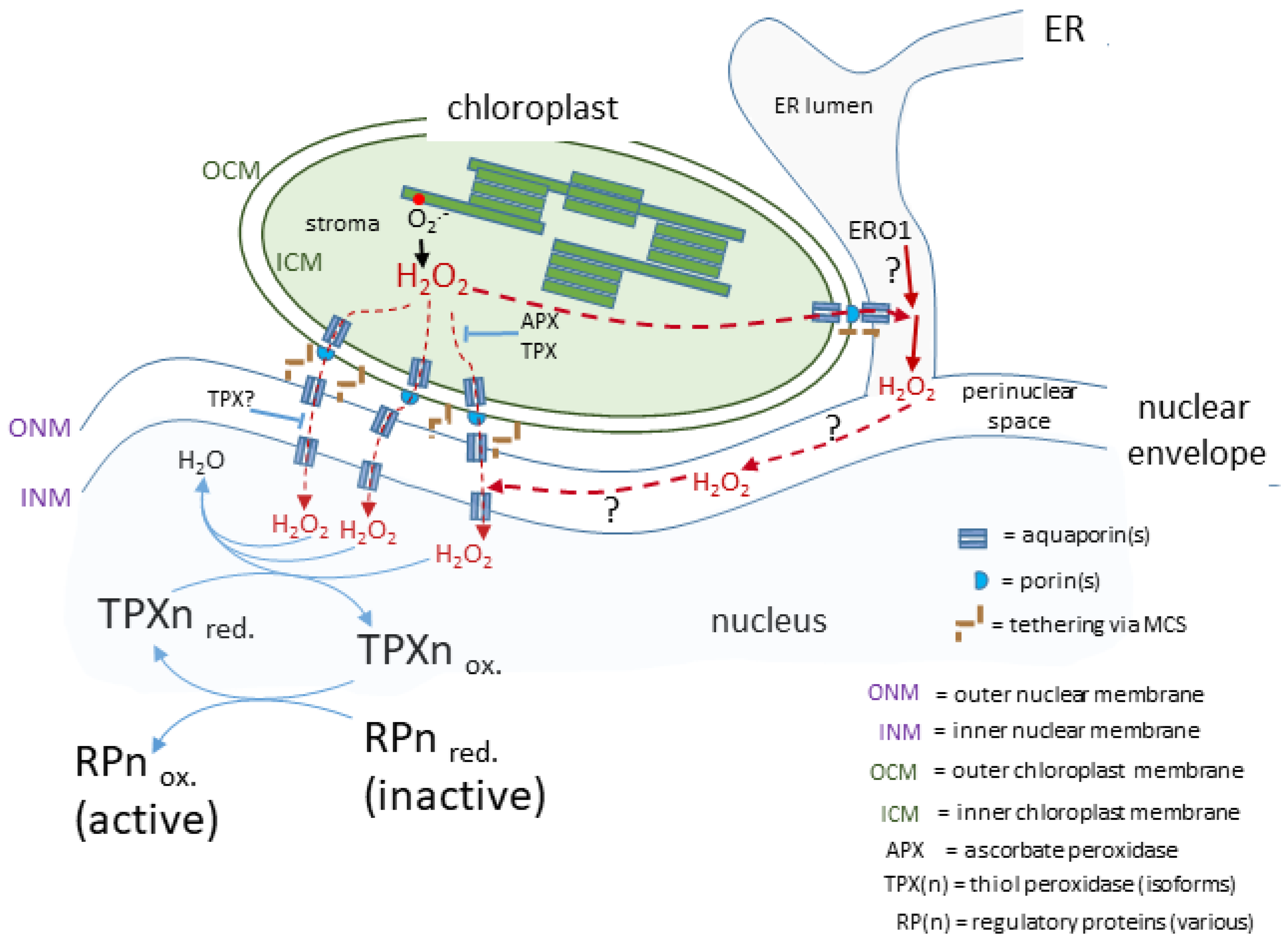
4. H2O2, Aquaporins and the Route to the Nucleus
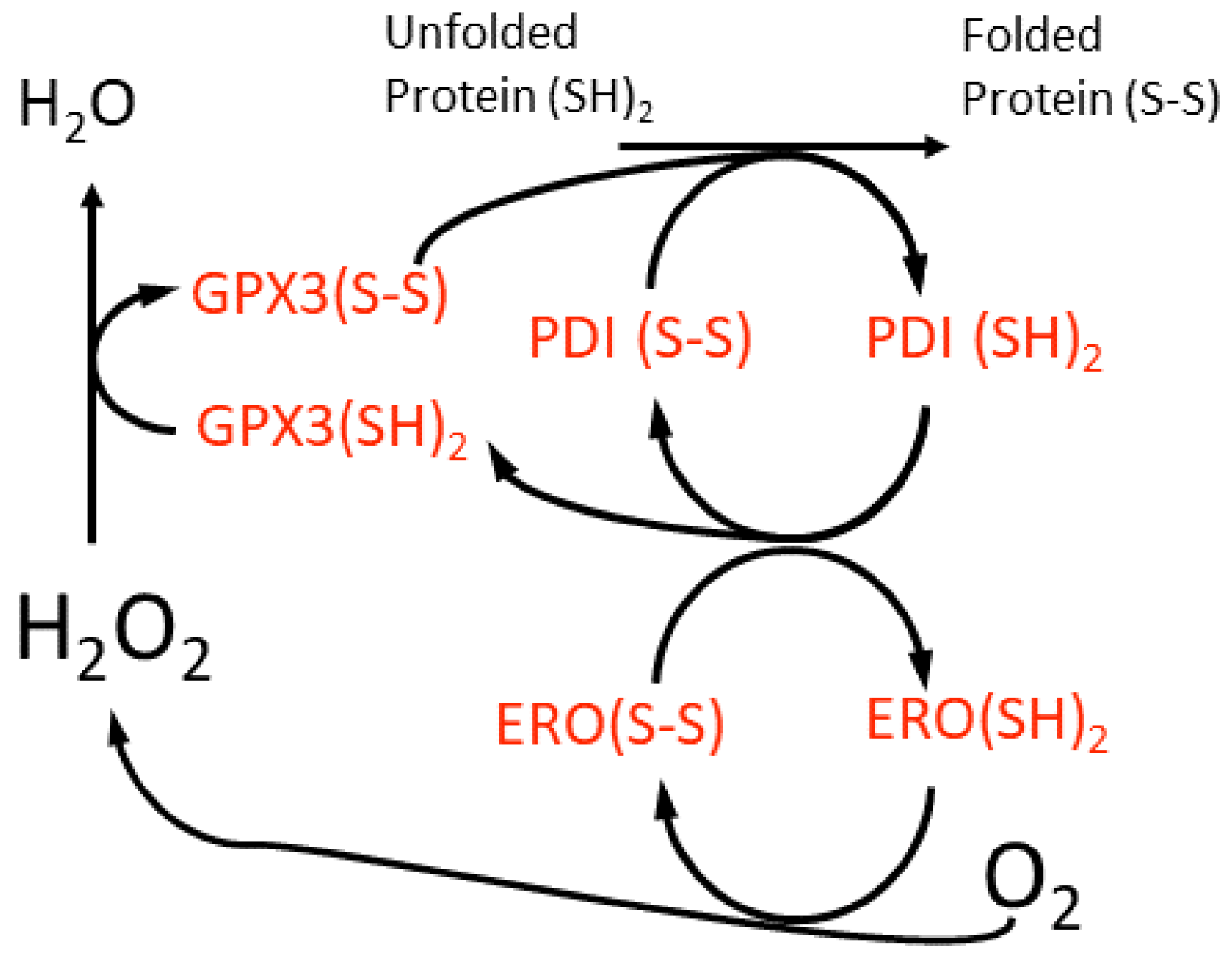
5. H2O2 in the Perinuclear Space and ER Lumen and Its Impact on Retrograde Signalling
6. Suppression of the UPR by High Light Intensities
7. Conclusions and Possibilities
Author Contributions
Funding
Conflicts of Interest
References
- Rossel, J.B.; Wilson, P.B.; Hussain, D.; Woo, N.S.; Gordon, M.J.; Mewett, O.P.; Howell, K.A.; Whelan, J.; Kazan, K.; Pogson, B.J. Systemic and Intracellular Responses to Photooxidative Stress in Arabidopsis. Plant Cell 2007, 19, 4091–4110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galvez-Valdivieso, G.; Fryer, M.J.; Lawson, T.; Slattery, K.; Truman, W.; Smirnoff, N.; Asami, T.; Davies, W.J.; Jones, A.M.; Baker, N.R.; et al. The High Light Response in Arabidopsis Involves ABA Signaling between Vascular and Bundle Sheath Cells. Plant Cell 2009, 21, 2143–2162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biswas, M.S.; Mano, J. Lipid Peroxide-Derived Short-Chain Carbonyls Mediate Hydrogen Peroxide-Induced and Salt-Induced Programmed Cell Death in Plants. Plant Physiol. 2015, 168, 885–898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Exposito-Rodriguez, M.; Laissue, P.P.; Yvon-Durocher, G.; Smirnoff, N.; Mullineaux, P.M. Photosynthesis-Dependent H2O2 Transfer from Chloroplasts to Nuclei Provides a High-Light Signalling Mechanism. Nat. Commun. 2017, 8, 49. [Google Scholar] [CrossRef] [Green Version]
- Calero-Muñoz, N.; Exposito-Rodriguez, M.; Collado-Arenal, A.M.; Rodríguez-Serrano, M.; Laureano-Marín, A.M.; Estrella Santamaría, M.; Gotor, C.; Díaz, I.; Mullineaux, P.M.; Romero-Puertas, M.C.; et al. Cadmium Induces Reactive Oxygen Species-Dependent Pexophagy in Arabidopsis Leaves. Plant Cell Environ. 2019, 42, 2696–2714. [Google Scholar] [CrossRef]
- Devireddy, A.R.; Zandalinas, S.I.; Fichman, Y.; Mittler, R. Integration of Reactive Oxygen Species and Hormone Signalling during Abiotic Stress. Plant J. 2021, 105, 459–476. [Google Scholar] [CrossRef]
- Xiong, H.; Hua, L.; Reyna-Llorens, I.; Shi, Y.; Chen, K.-M.; Smirnoff, N.; Kromdijk, J.; Hibberd, J.M. Photosynthesis-Independent Production of Reactive Oxygen Species in the Rice Bundle Sheath during High Light Is Mediated by NADPH Oxidase. Proc. Natl. Acad. Sci. USA 2021, 118, e2022702118. [Google Scholar] [CrossRef]
- Pesaresi, P.; Schneider, A.; Kleine, T.; Leister, D. Interorganellar Communication. Curr. Opin. Plant Biol. 2007, 10, 600–606. [Google Scholar] [CrossRef]
- Kangasjärvi, S.; Neukermanns, J.; Li, S.; Aro, E.-M.; Noctor, G. Photosynthesis, Photorespiration, and Light Signalling in Defence Responses. J. Exp. Bot. 2012, 63, 1619–1636. [Google Scholar] [CrossRef] [Green Version]
- Estavillo, G.M.; Crisp, P.A.; Pornsiriwong, W.; Wirtz, M.; Collinge, D.; Carrie, C.; Giraud, E.; Whelan, J.; David, P.; Javot, H.; et al. Evidence for a Sal1-Pap Chloroplast Retrograde Pathway that Functions in Drought and High Light Signaling in Arabidopsis. Plant Cell 2011, 23, 3992–4012. [Google Scholar] [CrossRef] [Green Version]
- Bobik, K.; Burch-Smith, T.M. Chloroplast Signalling within, between and beyond Cells. Front. Plant Sci. 2015, 6, 781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Souza, A.; Wang, J.; Dehesh, K. Retrograde Signals: Integrators of Interorganellar Communication and Orchestrators of Plant Development. Annu. Rev. Plant Biol. 2017, 68, 85–108. [Google Scholar] [CrossRef] [PubMed]
- Kerchev, P.I.; Van Breusegem, F. Improving Oxidative Stress Resilience in Plants. Plant J. 2021, 109, 359–372. [Google Scholar] [CrossRef] [PubMed]
- Selga, T.; Selga, M.; Gobins, V.; Ozolina, A. Plastid-Nuclear Complexes: Permanent Structures in Photosynthesizing of Vascular Plants. Environ. Exp. Biol. 2010, 8, 85–92. [Google Scholar]
- Mullineaux, P.M.; Exposito-Rodriguez, M.; Laissue, P.P.; Smirnoff, N.; Park, E. Spatial Chloroplast-to-Nucleus Signalling Involving Plastid–Nuclear Complexes and Stromules. Phil. Trans. R. Soc. B 2020, 375, 20190405. [Google Scholar] [CrossRef]
- Caplan, J.L.; Kumar, A.S.; Park, E.; Padmanabhan, M.S.; Hoban, K.; Modla, S.; Czymmek, K.; Dinesh-Kumar, S.P. Chloroplast Stromules Function during Innate Immunity. Dev. Cell 2015, 34, 45–57. [Google Scholar] [CrossRef] [Green Version]
- Ding, X.; Jimenez-Gongora, T.; Krenz, B.; Lozana-Duran, R. Chloroplast Clustering Around the Nucleus Is a General Response to Pathogen Perception in Nicotiana benthamiana. Mol. Plant Pathol. 2019, 20, 1298–1306. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, Y.; Zalabák, D.; Kubalová, I.; Králová, M.; Brenner, W.G.; Aida, M. Interpreting Cytokinin Action as Anterograde Signalling and beyond. Front. Plant Sci. 2021, 12, 641257. [Google Scholar] [CrossRef]
- Sewelam, N.; Jaspert, N.; Van Der Kelen, K.; Tognetti, V.B.; Schmitz, J.; Frerigmann, H.; Stahl, E.; Zeier, J.; Van Breusegem, F.; Maurino, V.G. Spatial H2O2 Signaling Specificity: H2O2 from Chloroplasts and Peroxisomes Modulates the Plant Transcriptome Differentially. Mol. Plant 2014, 7, 1191–1210. [Google Scholar] [CrossRef] [Green Version]
- Willems, P.; Mhamdi, A.; Stael, S.; Storme, V.; Kerchev, P.; Noctor, G.; Gevaert, K.; Van Breusegem, F. The ROS Wheel: Refining ROS Transcriptional Footprints. Plant Physiol. 2016, 171, 1720–1733. [Google Scholar] [CrossRef] [Green Version]
- Mignolet-Spruyt, L.; Xu, E.; Idänheimo, N.; Hoeberichts, F.A.; Mühlenbock, P.; Brosché, M.; Van Breusegem, F.; Kangasjärvi, J. Spreading the News: Subcellular and Organellar Reactive Oxygen Species Production and Signalling. J. Exp. Bot. 2016, 67, 3831–3844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, F.; Chi, Y.; Jiang, Z.; Xu, Y.; Xie, L.; Huang, F.; Wan, D.; Ni, J.; Yuan, F.; Wu, X.; et al. Hydrogen Peroxide Sensor HPCA1 Is an LRR Receptor Kinase in Arabidopsis. Nature 2020, 578, 577–581. [Google Scholar] [CrossRef] [PubMed]
- Mullineaux, P.M.; Karpinski, S.; Baker, N.R. Spatial Dependence for Hydrogen Peroxide–Directed Signaling in Light-Stressed Plants. Plant Physiol. 2006, 141, 346–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, K.X.; Mabbitt, P.D.; Phua, S.Y.; Mueller, J.W.; Nisar, N.; Gigolashvili, T.; Stroeher, E.; Grassl, J.; Arlt, W.; Estavillo, G.M.; et al. Sensing of Chloroplast Oxidative Stress by SAL1. Proc. Natl. Acad. Sci. USA 2016, 113, 4567–4576. [Google Scholar] [CrossRef] [Green Version]
- Vasseghi, M.-J.; Chibani, K.; Telman, W.; Liebthal, M.F.; Gerken, M.; Schnitzer, H.; Mueller, S.M.; Dietz, K.-J. The Chloroplast 2-Cysteine Peroxiredoxin Functions as Thioredoxin Reductase in Redox Regulation of Chloroplast Metabolism. eLife 2018, 7, e38194. [Google Scholar] [CrossRef]
- Savage, Z.; Duggan, C.; Toufexi, A.; Pandey, P.; Liang, Y.; Segretin, M.E.; Yuen, L.H.; Gaboriau, D.C.A.; Leary, A.Y.; Tumtas, Y.; et al. Chloroplasts Alter Their Morphology and Accumulate at the Pathogen Interface during Infection by Phytophthora infestans. Plant J. 2021, 107, 1771–1787. [Google Scholar] [CrossRef]
- de Torres Zabala, M.; Littlejohn, G.; Siddharth, J.; Studholme, D.; Bailey, T.; Lawson, T.; Tillich, M.; Licht, D.; Bölter, B.; Delfino, L.; et al. Chloroplasts Play a Central Role in Plant Defence and Are Targeted by Pathogen Effectors. Nat. Plants 2015, 1, 15074. [Google Scholar] [CrossRef]
- Costa, A.; Drago, I.; Behera, S.; Zottini, M.; Pizzo, P.; Schroeder, J.I.; Pozzan, T.; Schiavo, F.L. H2O2 in Plant Peroxisomes: An In Vivo Analysis Uncovers a Ca2+-Dependent Scavenging System. Plant J. 2010, 62, 760–772. [Google Scholar] [CrossRef] [Green Version]
- Smirnoff, N.; Arnaud, D. Hydrogen Peroxide Metabolism and Functions in Plants. New Phytol. 2019, 221, 1197–1214. [Google Scholar] [CrossRef]
- Hu, C.-H.; Wang, P.-Q.; Zhang, P.-P.; Nie, X.-M.; Li, B.-B.; Tai, L.; Liu, W.-T.; Li, W.-Q.; Chen, K.-M. NADPH Oxidases: The Vital Performers and Center Hubs during Plant Growth and Signalling. Cells 2020, 9, 437. [Google Scholar] [CrossRef] [Green Version]
- Mubarakshina, M.M.; Ivanov, B.N.; Naydov, I.A.; Hillier, W.; Badger, M.R.; Krieger-Liszkay, A. Production and Diffusion of Chloroplastic H2O2 and Its Implication to Signalling. J. Exp. Bot. 2010, 61, 3577–3587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, A.S.; Park, E.; Nedo, A.; Alqarni, A.; Ren, L.; Hoban, K.; Modla, S.; McDonald, J.H.; Kambhamettu, C.; Dinesh-Kumar, S.P.; et al. Stromule Extension along Microtubules Coordinated with Actin-Mediated Anchoring Guides Perinuclear Chloroplast Movement during Innate Immunity. eLife 2018, 7, e23625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhai, Y.; Yuan, Q.; Qiu, S.; Li, S.; Li, M.; Zheng, H.; Wu, G.; Lu, W.; Peng, J.; Rao, S.; et al. Turnip Mosaic Virus Impairs Perinuclear Chloroplast Clustering to Facilitate Viral Infection. Plant Cell Environ. 2021, 44, 3681–3699. [Google Scholar] [CrossRef] [PubMed]
- Hanson, M.R.; Hines, K.M. Stromules: Probing Form and Function. Plant Physiol. 2018, 176, 128–137. [Google Scholar] [CrossRef] [Green Version]
- Breeze, E.; Vale, V.; McLellan, H.; Godiard, L.; Grant, M.; Frigerio, L. The Plant Endoplasmic Reticulum Is both Receptive and Responsive to Pathogen Effectors. bioRxiv 2020, 9, 142141. [Google Scholar]
- Prunuske, A.J.; Ullman, K.S. The Nuclear Envelope: Form and Reformation. Curr. Opin. Cell Biol. 2006, 18, 108–116. [Google Scholar] [CrossRef] [Green Version]
- Andersson, M.X.; Goksör, M.; Sandelius, A.S. Optical Manipulation Reveals Strong Attracting Forces at Membrane Contact Sites between Endoplasmic Reticulum and Chloroplasts. J. Biol. Chem. 2007, 282, 1170–1174. [Google Scholar] [CrossRef]
- Scorrano, L.; De Matteis, M.A.; Emr, S.; Giordano, F.; Hajnóczky, G.; Kornmann, B.; Lackner, L.L.; Levine, T.P.; Pellegrini, L.; Reinisch, K.; et al. Coming Together to Define Membrane Contact Sites. Nat. Commun. 2019, 10, 1287. [Google Scholar]
- Baillie, A.L.; Falz, A.-L.; Müller-Schüssele, S.J.; Sparkes, I. It Started with a Kiss: Monitoring Organelle Interactions and Identifying Membrane Contact Site Components in Plants. Front. Plant Sci. 2020, 11, 517. [Google Scholar] [CrossRef]
- Gibbs, S.P. Nuclear Envelope-Chloroplast Relationships in Algae. J. Cell Sci. 1962, 14, 433–444. [Google Scholar] [CrossRef] [Green Version]
- Crotty, W.J.; Ledbetter, M.C. Membrane Continuities Involving Chloroplasts and Other Organelles in Plant Cells. Science 1973, 182, 839–842. [Google Scholar] [CrossRef] [PubMed]
- McLean, B.; Whatley, J.M.; Juniper, B.E. Continuity of Chloroplast and Endoplasmic Reticulum Membranes in Chara and Equisetum. New Phytol. 1988, 109, 59–65. [Google Scholar] [CrossRef]
- Whately, J.M.; McLean, B.; Juniper, B.E. Continuity of Chloroplast and Endoplasmic Reticulum Membranes in Phaseolus vulgaris. New Phytol. 1991, 117, 209–217. [Google Scholar] [CrossRef]
- Tan, X.; Wang, Q.; Tian, B.; Zhang, H.; Lu, D.; Zhou, J. A Brassica napus Lipase Locates at the Membrane Contact Sites Involved in Chloroplast Development. PLoS ONE 2011, 6, e26831. [Google Scholar] [CrossRef]
- Mehrshahi, P.; Stefano, G.; Andaloro, J.M.; Brandizzi, F.; Froehlich, J.E.; DellaPenna, D. Transorganellar Complementation Redefines the Biochemical Continuity of Endoplasmic Reticulum and Chloroplasts. Proc. Natl. Acad. Sci. USA 2013, 110, 12126–12131. [Google Scholar] [CrossRef] [Green Version]
- Andersson, M.X.; Goksör, M.; Sandelius, A.S. Physical Attachment between Chloroplasts and Endoplasmic Reticulum Revealed by Optical Manipulation. Plant Signal. Behav. 2007, 2, 185–187. [Google Scholar] [CrossRef] [Green Version]
- Perico, C.; Sparkes, I. Plant Organelle Dynamics: Cytoskeletal Control and Membrane Contact Sites. New Phytol. 2018, 220, 381–394. [Google Scholar] [CrossRef] [Green Version]
- Briggs, W.R.; Beck, C.F.; Cashmore, A.R.; Christie, J.M.; Hughes, J.; Jarillo, J.A.; Kagawa, T.; Kanegae, H.; Liscum, E.; Nagatani, A.; et al. The Phototropin Family of Photoreceptors. Plant Cell 2001, 13, 993–997. [Google Scholar] [CrossRef] [Green Version]
- Wen, F.; Xing, D.; Zhang, L. Hydrogen Peroxide Is Involved in High Blue Light-Induced Chloroplast Avoidance Movements in Arabidopsis. J. Exp. Bot. 2008, 59, 2891–2901. [Google Scholar] [CrossRef] [Green Version]
- Kong, S.-G.; Shimada, A.; Kijima, S.T.; Hirose, K.; Katoh, K.; Ahn, J.; Higa, T.; Takano, A.; Nakamura, Y.; Suetsugu, N.; et al. CHLOROPLAST UNUSUAL POSITIONING 1 Is a New Type of Actin Nucleation Factor in Plants. bioRxiv 2020, 14, 905984. [Google Scholar]
- Suetsugu, N.; Yamada, N.; Kagawa, T.; Yonekura, H.; Uyeda, T.Q.P.; Kadota, A.; Wada, M. Two Kinesin-Like Proteins Mediate Actin-Based Chloroplast Movement in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2010, 107, 8860–8865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suetsugu, N.; Higa, T.; Gotoh, E.; Wada, M. Light-Induced Movements of Chloroplasts and Nuclei Are Regulated in both Cp-Actin-Filament-Dependent and -Independent Manners in Arabidopsis thaliana. PLoS ONE 2016, 11, e0157429. [Google Scholar]
- Oikawa, K.; Kasahara, M.; Kiyosue, T.; Kagawa, T.; Suetsugu, N.; Takahashi, F.; Kanegae, T.; Niwa, Y.; Kadota, A.; Wada, M. CHLOROPLAST UNUSUAL POSITIONING1 Is Essential for Proper Chloroplast Positioning. Plant Cell 2003, 15, 2805–2815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higa, T.; Suetsugu, N.; Kong, S.; Wada, M. Actin-Dependent Plastid Movement is Required for Motive Force Generation in Directional Nuclear Movement in Plants. Proc. Natl. Acad. Sci. USA 2014, 111, 4327–4331. [Google Scholar] [CrossRef] [Green Version]
- Glynn, J.M.; Yang, Y.; Vitha, S.; Schmitz, A.J.; Hemmes, M.; Miyagishima, S.; Osteryoung, K.W. PARC6, a Novel Chloroplast Division Factor, Influences FtsZ Assembly and Is Required for Recruitment of PDV1 during Chloroplast Division in Arabidopsis. Plant J. 2009, 59, 700–711. [Google Scholar] [CrossRef]
- Itoh, R.D.; Ishikawa, H.; Nakajima KPMoriyama, S.; Fujiwara, M.T. Isolation and Analysis of a Stromule-Overproducing Arabidopsis Mutant Suggest the Role of PARC6 in Plastid Morphology Maintenance in the Leaf Epidermis. Physiol. Plant. 2018, 162, 474–494. [Google Scholar] [CrossRef]
- Oikawa, K.; Yamasato, A.; Kong, S.-G.; Kasahara, M.; Nakai, M.; Takahashi, F.; Ogura, Y.; Kagawa, T.; Wada, M. Chloroplast Outer Envelope Protein CHUP1 Is Essential for Chloroplast Anchorage to the Plasma Membrane and Chloroplast Movement. Plant Physiol. 2008, 148, 829–842. [Google Scholar] [CrossRef] [Green Version]
- Schmidt von Braun, S.; Schleiff, E. The Chloroplast Outer Membrane Protein CHUP1 Interacts with Actin and Profilin. Planta 2008, 227, 1151–1159. [Google Scholar] [CrossRef]
- Lehmann, P.; Bohnsack, M.T.; Schleiff, E. The Functional Domains of the Chloroplast Unusual Positioning Protein 1. Plant Sci. 2011, 180, 650–654. [Google Scholar] [CrossRef]
- van Gisbergen, P.A.C.; Bezanilla, M. Plant Formins: Membrane Anchors for Actin Polymerization. Trends Cell Biol. 2013, 23, 227–233. [Google Scholar] [CrossRef]
- Kong, S.G.; Wada, M. New Insights into Dynamic Actin-Based Chloroplast Photo-Relocation Movement. Mol. Plant 2011, 4, 771–781. [Google Scholar] [CrossRef] [PubMed]
- Meier, I.; Brkljacic, J. The Arabidopsis Nuclear Pore and Nuclear Envelope. Arab. Book 2010, 8, e0139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groves, N.R.; Biel, A.M.; Newman-Griffis, A.H.; Meier, I. Dynamic Changes in Plant Nuclear Organization in Response to Environmental and Developmental Signals. Plant Physiol. 2018, 176, 230–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, Y. The Nuclear Pore Complex: A Strategic Platform for Regulating Cell Signalling. New Phytol. 2017, 219, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Krupinska, K.; Blanco, N.E.; Oetke, S.; Zottini, M. Genome Communication in Plants Mediated by Organelle-Nucleus-Located Proteins. Phil. Trans. R. Soc. B 2020, 375, 20190397. [Google Scholar] [CrossRef] [PubMed]
- Bienert, G.P.; Chaumont, F. Aquaporin-Facilitated Transmembrane Diffusion of Hydrogen Peroxide. Biochim. Biophys. Acta 2014, 1840, 1596–1604. [Google Scholar] [CrossRef]
- Orrico, F.; Lopez, A.C.; Saliwonczyk, D.; Acosta, C.; Rodrgiuez-Grecco, I.; Mouro-Chanteloup, I.; Ostuni, M.A.; Denicola, A.; Thomson, L.; Möller, M.N. The Permeability of Human Red Blood Cell Membranes to Hydrogen Peroxide Is Independent of Aquaporins. J. Biol. Chem. 2021, 298, 101503. [Google Scholar] [CrossRef]
- Appenzeller-Herzog, C.; Bánhegyi, G.; Bogeski, I.; Davies, K.J.A.; Delaunay-Moisan, A.; Forman, H.J.; Görlach, A.; Kietzmann, T.; Laurindo, F.; Margittai, E.; et al. Transit of H2O2 across the Endoplasmic Reticulum Membrane Is not Sluggish. Free. Rad. Biol. Med. 2016, 94, 157–160. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Schoebel, S.; Schmitz, F.; Dong, H.; Hedfalk, K. Characterization of Aquaporin-Driven Hydrogen Peroxide Transport. BBA Biomembr. 2020, 1862, 183065. [Google Scholar] [CrossRef]
- Borisova Mubarakshina, M.M.; Kozuleva, M.A.; Rudenko, N.N.; Naydov, I.A.; Klenina, I.B.; Ivanov, B.N. Photosynthetic Electron Flow to Oxygen and Diffusion of Hydrogen Peroxide through the Chloroplast Envelope via Aquaporins. Biochim. Biophys. Acta 2012, 1817, 1314–1321. [Google Scholar] [CrossRef] [Green Version]
- Maurel, C.; Bousiac, Y.; Luu, D.-T.; Santoni, V.; Shahzad, Z.; Verdoucq, L. Aquaporins in Plants. Physiol. Rev. 2015, 95, 1321–1358. [Google Scholar] [CrossRef] [PubMed]
- Simm, S.; Papasotiriou, D.G.; Ibrahim, M.; Leisepang, M.S.; Muller, B.; Schorge, T.; Karas, M.; Mirus, O.; Sommer, M.S.; Schleiff, E. Defining the Core Proteome of the Chloroplast Envelope Membranes. Front. Plant Sci. 2013, 4, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, L.; Lu, Z.; Ding, L.; Guo, J.; Wang, M.; Ling, N.; Guo, S.; Shen, Q. Role of Aquaporins in Determining Carbon and Nitogen Status in Higher Plants. Int. J. Mol. Sci. 2018, 19, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beebo, A.; Zia, A.; Kinzel, C.R.; Herdean, A.; Bouhidel, K.; Kirchoff, H.; Scoefs, B.; Spetea, C. TIP Family Aquaporins Play Role in Chloroplast Osmoregulation and Photosynthesis. bioRxiv 2020, 18, 297978. [Google Scholar]
- Booth, D.M.; Enyedi, B.; Geiszt, M.; Varnai, P.; Hajnóczky, G. Redox Nanodomains Are Induced by and Control Calcium Signalling at the ER-Mitochondrial Interface. Mol. Cell 2016, 63, 240–248. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Berchner-Pfannschmidt, U.; Möller, U.; Brecht, M.; Wotzlaw, C.; Acker, H.; Jungermann, K.; Kietzmann, T. A Fenton Reaction at the Endoplasmic Reticulum Is Involved in the Redox Control of Hypoxia-inducible Gene Expression. Proc. Natl. Acad. Sci. USA 2004, 101, 4302–4307. [Google Scholar] [CrossRef] [Green Version]
- Melo, E.P.; Lopes, C.; Gollwitzer, P.; Lortz, S.; Lenzen, S.; Mehmeti, I.; Kaminski, C.F.; Ron, D.; Avezov, E. TriPer, an Optical Probe Tuned to the Endoplasmic Reticulum Tracks Changes in Luminal H2O2. BMC Biol. 2017, 15, 24. [Google Scholar] [CrossRef] [Green Version]
- Yobue, E.D.; Sitia, R.; Simmen, T. Redox Crosstalk at Endoplasmic Reticulum (ER) Membrane Contact Sites (MCS) Uses Toxic Waste to Deliver Messages. Cell Death Dis. 2018, 9, 331. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Li, J. Communications between Endoplasmic Reticulum and Other Organelles during Abiotic Stress Response in Plants. Front. Plant Sci. 2019, 10, 749. [Google Scholar] [CrossRef]
- Birk, J.; Meyer, M.; Aller, I.; Hansen, H.G.; Odermatt, A.; Dick, T.P.; Meyer, A.J.; Appenzeller-Herzog, C. Endoplasmic Reticulum: Reduced and Oxidized Glutathione Revisited. J. Cell Sci. 2013, 126, 1604–1617. [Google Scholar] [CrossRef] [Green Version]
- Ugalde, J.M.; Aller, I.; Kudrjasova, L.; Schmidt, R.R.; Schlöβer, M.; Homagk, M.; Fuchs, P.; Lichtenauer, S.; Schwarzländer, M.; Műller-Schűssele, S.J.; et al. Endoplasmic Reticulum Oxidoreductin (ERO) Provides Resilience against Reductive Stress and Hypoxic Conditions by Mediating Luminal Redox Dynamics. bioRxiv 2021, 13, 472397. [Google Scholar]
- Araki, K.; Iemura, S.; Kamiya, Y.; Ron, D.; Kato, K.; Natsume, T.; Nagata, K. Ero1α and PDIs Constitute a Hierarchical Electron Transfer Network of Endoplasmic Reticulum Oxidoreductases. J. Cell Biol. 2013, 202, 861–874. [Google Scholar] [CrossRef] [PubMed]
- Konno, T.; Melo, E.P.; Lopes, C.; Mehmeti, I.; Lenzen, S.; Ron, D.; Avezov, E. ERO1-Independent Production of H2O2 within the Endoplasmic Reticulum Fuels Prdx4-Mediated Oxidative Protein Folding. J. Cell Biol. 2015, 211, 253–259. [Google Scholar] [CrossRef] [Green Version]
- Meyer, A.J.; Dreyer, A.; Ugalde, J.M.; Feitosa-Araujo, E.; Dietz, K.-J.; Schwärzlander, M. Shifting Paradigms and Novel Players in Cys-Based Redox Regulation and ROS Signalling in Plants–And Where to Go Next. Biol. Chem. 2020, 402, 399–423. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.-S.; Yang, H.; Ding, L.; Song, Z.-T.; Ma, H.; Chang, F.; Liu, J.-X. Tissue-Specific Transcriptomics Reveals an Important Role of the Unfolded Protein Response in Maintaining Fertility Upon Heat Stress in Arabidopsis. Plant Cell 2017, 29, 1007–1023. [Google Scholar] [CrossRef]
- Albihlal, W.S.; Obomighie, I.; Blein, T.; Persad, R.; Chernukhin, I.; Crespi, M.; Bechtold, U.; Mullineaux, P.M. Arabidopsis HEAT SHOCK TRANSCRIPTION FACTORA1b Regulates Multiple Developmental Genes under Benign and Stress Conditions. J. Exp. Bot. 2018, 69, 2847–2862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beaugelin, I.; Chevalier, A.; D’Alessandro, S.; Ksas, B.; Havaux, M. Endoplasmic Reticulum-Mediated Unfolded Protein Response Is an Integral Part of Singlet Oxygen Signalling in Plants. Plant J. 2020, 102, 1266–1280. [Google Scholar] [CrossRef] [PubMed]
- Mullineaux, P.M.; Exposito-Rodriguez, M.; Laissue, P.P.; Smirnoff, N. ROS-Dependent Signalling Pathways in Plants and Algae Exposed to High Light: Comparisons with Other Eukaryotes. Free Rad. Biol. Med. 2018, 122, 52–64. [Google Scholar] [CrossRef]
- Nawkar, G.M.; Kang, C.H.; Maibam, P.; Park, J.H.; Jung, Y.J.; Chae, H.B.; Chi, Y.H.; Jung, I.J.; Kim, W.Y.; Yun, D.-J.; et al. HY5, a Positive Regulator of Light Signalling, Negatively Controls the Unfolded Protein Response in Arabidopsis. Proc. Natl. Acad. Sci. USA 2016, 114, 2084–2089. [Google Scholar] [CrossRef] [Green Version]
- Alvarez-Fernandez, R.; Penfold, C.A.; Galvez-Valdivieso, G.; Exposito-Rodriguez, M.; Stallard, E.J.; Bowden, L.; Moore, J.D.; Mead, A.; Davey, P.A.; Matthews, J.S.A.; et al. Time Series Transcriptomics Reveals a BBX32- and Photoreceptor-Directed Acclimation of Photosynthesis to High Light in Mature Arabidopsis Leaves. Plant J. 2021, 107, 1363–1386. [Google Scholar] [CrossRef]
- Fryer, M.J.; Ball, L.; Oxborough, K.; Karpinski, S.; Mullineaux, P.M.; Baker, N.R. Control of Ascorbate Peroxidase 2 Expression by Hydrogen Peroxide and Leaf Water Status during Excess Light Stress Reveals a Functional Organisation of Arabidopsis Leaves. Plant J. 2003, 33, 691–705. [Google Scholar] [CrossRef] [PubMed]
- Műhlenbock, P.; Szechyńska-Hebda, M.; Płaszczyca, M.; Baudo, M.; Mateo, A.; Mullineaux, P.M.; Parker, J.E.; Karpińska, B.; Karpiński, S. Chloroplast Signaling and LESION SIMULATING DISEASE1 Regulate Crosstalk between Light Acclimation and Immunity in Arabidopsis. Plant Cell 2008, 20, 2339–2356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramel, F.; Birtic, S.; Ginies, C.; Soubigou-Taconnat, L.; Triantaphylides, C.; Havaux, M. Carotenoid Oxidation Products Are Stress Signals that Mediate Gene Responses to Singlet Oxygen in Plants. Proc. Natl. Acad. Sci. USA 2012, 109, 5535–5540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fomenko, D.E.; Koc, A.; Agisheva, N.; Jacobsen, M.; Kaya, A.; Malinouski, M.; Rutherford, J.C.; Siu, K.-L.; Jin, D.-Y.; Winge, D.R.; et al. Thiol Peroxidases Mediate Specific Genome-Wide Regulation of Gene Expression in Response to Hydrogen Peroxide. Proc. Natl. Acad. Sci. USA 2011, 108, 2729–2734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chibani, K.; Wingsle, G.; Jacquot, J.-P.; Gelhaye, E.; Rouhier, N. Comparative Genomic Study of the Thioredoxin Family in Photosynthetic Organisms with Emphasis on Populus trichocarpa. Mol. Plant 2009, 2, 308–322. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Breeze, E.; Mullineaux, P.M. The Passage of H2O2 from Chloroplasts to Their Associated Nucleus during Retrograde Signalling: Reflections on the Role of the Nuclear Envelope. Plants 2022, 11, 552. https://doi.org/10.3390/plants11040552
Breeze E, Mullineaux PM. The Passage of H2O2 from Chloroplasts to Their Associated Nucleus during Retrograde Signalling: Reflections on the Role of the Nuclear Envelope. Plants. 2022; 11(4):552. https://doi.org/10.3390/plants11040552
Chicago/Turabian StyleBreeze, Emily, and Philip M. Mullineaux. 2022. "The Passage of H2O2 from Chloroplasts to Their Associated Nucleus during Retrograde Signalling: Reflections on the Role of the Nuclear Envelope" Plants 11, no. 4: 552. https://doi.org/10.3390/plants11040552