
The Biology of the Genus Ceiba, a Potential Source for Sustainable Production of Natural Fiber


Abstract
:1. Introduction
2. Characteristics of the Genus Ceiba
3. Ecology of Ceiba Species
3.1. The Ecosystem and the Challenges Faced by Ceiba Species Due to Habitat Loss and Degradation
3.2. Conservation and Ecosystem Restoration Potential of Ceiba Species
3.2.1. Ceiba aesculifolia
3.2.2. Ceiba pentandra
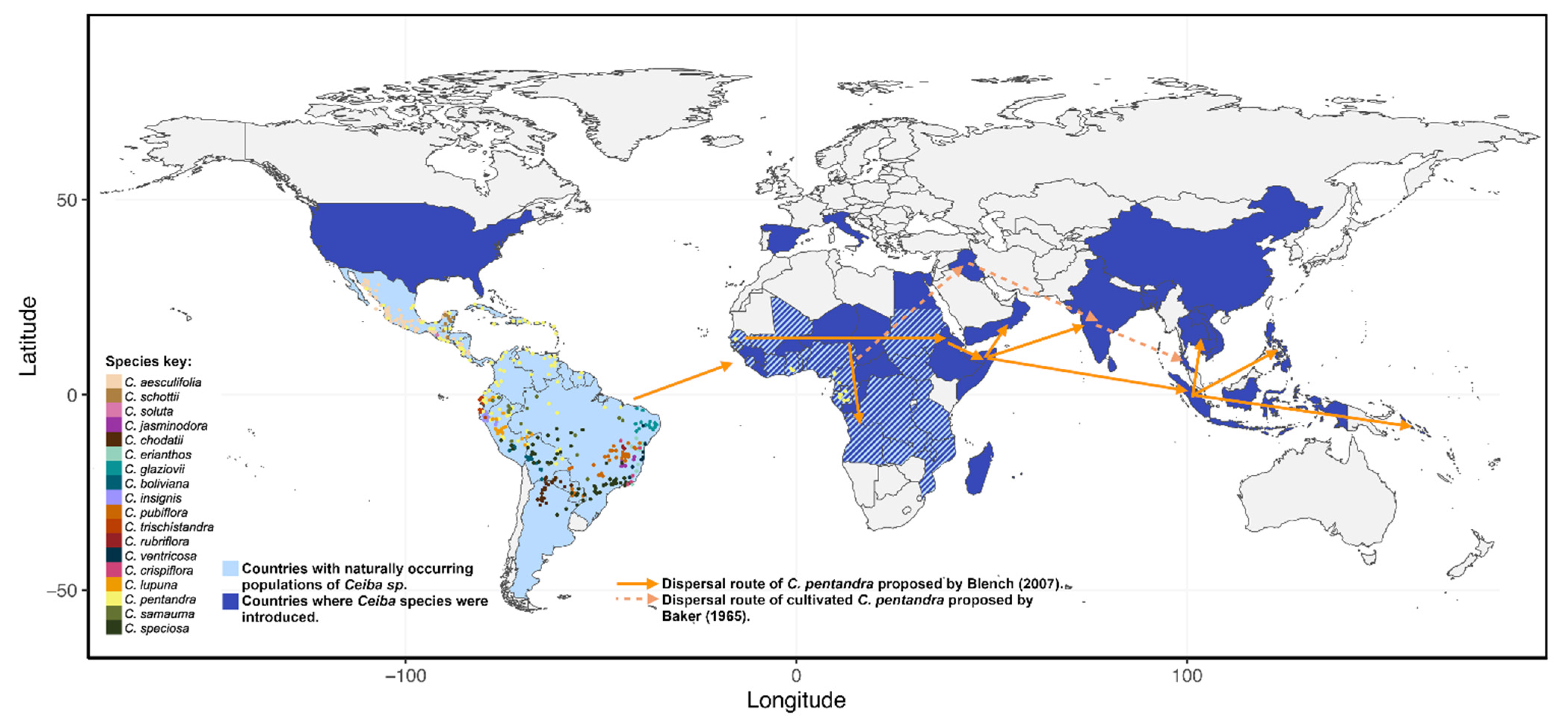
4. A Brief History on the Origin of the Clade Ceiba and the Arrival of Cultivated C. pentandra to Asia
5. Kapok Fiber Characteristics and Uses
6. Other Exploitable Resources from Ceiba Species
7. High Throughput Technologies and Molecular Approaches towards Plant-Resource Management
8. The Future of the Genus Ceiba and Perspectives
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Townsend, T.; Sette, J. Natural Fibres and the World Economy. In Natural Fibres: Advances in Science and Technology towards Industrial Applications; Fangueiro, R., Rana, S., Eds.; Springer: Dordrecht, The Netherland, 2016; Volume 12, pp. 381–390. [Google Scholar] [CrossRef]
- Chen, H.L.; Burns, L.D. Environmental analysis of textile products. CTRJ 2006, 24, 248–261. [Google Scholar] [CrossRef]
- Gedik, G.; Avinc, O. Hemp fiber as a sustainable raw material source for textile industry: Can we use its potential for more eco-friendly production? In Sustainability in the Textile and Apparel Industries; Muthu, S.S., Gardetti, M.A., Eds.; Springer Nature: Chaum, Switzerland, 2020; pp. 87–109. [Google Scholar] [CrossRef]
- Carvalho-Sobrinho, J.G.; Alverson, W.S.; Alcantara, S.; Queiroz, L.P.; Mota, A.C.; Baum, D.A. Revisiting the phylogeny of Bombacoideae (Malvaceae): Novel relationships, morphologically cohesive clades, and a new tribal classification based on multilocus phylogenetic analyses. Mol. Phylogenet. Evol. 2016, 101, 56–74. [Google Scholar] [CrossRef]
- Zeven, A.C. Kapok tree, Ceiba pentandra Gaertn. In Outlines of Perennial Crop Breeding in the Tropics; Ferwerda, F.P., Wit, F., Eds.; Landbouwhogeschool: Wageningen, The Netherlands, 1969; pp. 269–287. [Google Scholar]
- Zheng, Y.; Wang, J.; Wang, A. Recent advances in the potential applications of hollow kapok fiber-based functional materials. Cellulose 2021, 28, 5262–5292. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. FAOSTAT Statistical Database. Rome, Italy. Available online: https://www.fao.org/faostat/en/#home (accessed on 5 January 2022).
- Blench, R.M. The intertwined history of the silk-cotton and baobab. In Fields of Change: Progress in African Archaeobotany; Cappers, R.T.J., Ed.; Barkhuis: Eelde, The Netherlands, 2007; Volume 5, pp. 1–19. [Google Scholar]
- Barrera-Vázquez, A. La ceiba-cocodrilo. Anales INAH 1975, 7, 187–208. [Google Scholar]
- López-Austin, A. Las cuatro columnas. Arqueol. Mex. 2019, 83, 20–29. Available online: https://arqueologiamexicana.mx (accessed on 19 May 2020).
- Comisión Nacional de Áreas Naturales Protegidas. De Mitos y Leyendas, la Ceiba. Gobierno de México, 2021. Available online: https://www.gob.mx/conanp (accessed on 8 August 2021).
- Hurtado, V. Datos Interesantes Sobre Algunos de los Símbolos Patrios de Guatemala. The Foundation for Latin American Anthropological Research (FLAAR Mesoamérica), 2021. Available online: https://flaar-mesoamerica.org (accessed on 16 May 2020).
- Pezzini, F.F.; Dexter, K.G.; de Carvalho-Sobrinho, J.G.; Kidner, C.A.; Nicholls, J.A.; De Queiroz, L.P.; Pennington, R.T. Phylogeny and biogeography of Ceiba Mill. (Malvaceae, Bombacoideae). Front. Biogeogr. 2021, 13, 1–18. [Google Scholar] [CrossRef]
- Gibbs, P.; Semir, J. A taxonomic revision of the genus Ceiba Mill. (Bombacaceae). Anales Jard. Bot. Madrid 2003, 60, 259–300. [Google Scholar] [CrossRef]
- De Carvalho-Sobrinho, J.G.; de Queiroz, L.P. Ceiba rubriflora (Malvaceae: Bombacoideae), a new species from Bahia, Brazil. Kew Bull. 2008, 63, 649–653. [Google Scholar] [CrossRef]
- Niembro-Rocas, A.; Vázquez-Torres, M.; Sánchez-Sánchez, O. Árboles de Veracruz: 100 Especies Para la Reforestación Estratégica; Universidad Veracruzana: Mexico City, Mexico, 2010. [Google Scholar]
- Duno de Stefano, R.; Carnevali Fernández-Concha, G.; Ramírez Morillo, I.M.; Tapia Muñoz, J.L.; Can Itzá, L.L.; Hernández-Aguilar, S.; Embray, T. Flora de la Península de Yucatán. 2010. Revised on January 2022 Herbario CICY, Flora de la Península de Yucatán. Available online: https://www.cicy.mx/sitios/flora%20digital/ficha_virtual.php?especie=1746 (accessed on 10 January 2022).
- Bullock, S.H.; Mooney, H.A.; Medina, E. Seasonally Dry Tropical Forests; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- Dirzo, R.; Young, H.S.; Mooney, H.A.; Ceballos, G. Seasonally Dry Tropical Forests: Ecology and Conservation; Island Press: Washington, DC, USA, 2011; pp. xi–xiii. [Google Scholar] [CrossRef]
- Linares-Palomino, R.; Oliveira-Filho, A.T.; Pennington, R.T. Neotropical Seasonally Dry Forests: Diversity, Endemism, and Biogeography of Woody Plants. In Seasonally Dry Tropical Forests; Dirzo, R., Young, H.S., Mooney, H.A., Ceballos, G., Eds.; Island Press: Washington, DC, USA, 2011; pp. 3–21. [Google Scholar] [CrossRef]
- Lopezaraiza-Mikel, M.; Quesada, M.; Álvarez-Añorve, M.; Ávila-Cabadilla, L.; Martén-Rodríguez, S.; Calvo-Alvaado, J.; do Espírito-Santo, M.M.; Fernandes, G.W.; Sánchez-Azofeifa, A.; Aguilar-Aguilar, M.J.; et al. Phenological patterns of tropical dry forests along latitudinal and successional gradients in the neotropics. In Tropical Dry Forests in the Americas; Sanchez-Azofeifa, A., Powers, J.S., Fernandes, G.W., Quesada, M., Eds.; CRC Press: Boca Raton, FL, USA, 2013; pp. 119–146. [Google Scholar]
- Maas, M.; Burgos, A. Water dynamics and the ecosystem level in seasonally dry tropical forests Seasonally Dry Tropical Forests: Ecology and Conservation. In Seasonally Dry Tropical Forests; Dirzo, R., Young, H.S., Mooney, H.A., Ceballos, G., Eds.; Island Press: Washington, DC, USA, 2011; pp. 141–156. [Google Scholar] [CrossRef]
- Herrerías-Diego, Y.; Quesada, M.; Stoner, K.E.; Lobo, J.A. Effects of forest fragmentation on phenological patterns and reproductive success of the tropical dry forest tree Ceiba aesculifolia. Conserv. Biol. 2006, 20, 1111–1120. [Google Scholar] [CrossRef]
- Valle-Díaz, O.; Blanco-García, A.; Bonfil, C.; Paz, H.; Lindig-Cisneros, R. Altitudinal range shift detected through seedling survival of Ceiba aesculifolia in an area under the influence of an urban heat island. Forest Ecol. Manag. 2009, 258, 1511–1515. [Google Scholar] [CrossRef]
- Brondani, R.P.V.; Gaiotto, F.A.; Missiaggia, A.A.; Kirst, M.; Gribel, R.; Grattapaglia, D. Microsatellite markers for Ceiba pentandra (Bombacaceae), an endangered tree species of the Amazon Forest. Mol. Ecol. Notes 2003, 3, 177–179. [Google Scholar] [CrossRef] [Green Version]
- IUCN 2022. The International Union for Conservation of Nature and Natural Resources Red List of Threatened Species. Version 2021-3. Available online: https://www.iucnredlist.org/ (accessed on 16 January 2022).
- Cusack, D.F.; Karpman, J.; Ashdown, D.; Cao, Q.; Ciochina, M.; Halterman, S.; Lydon, S.; Neupane, A. Global change effects on humid tropical forests: Evidence for biogeochemical and biodiversity shifts at an ecosystem scale. Rev. Geophys. 2016, 54, 523–610. [Google Scholar] [CrossRef] [Green Version]
- Velázquez-Rosas, N.; Ruiz-Guerra, B.; Sánchez-Coronado, M.E.; Gamboa de Buen, A.; Orozco-Segovia, A. Morphological variation in fruits and seeds of Ceiba aesculifolia and its relationship with germination and seedling biomass. Bot. Sci. 2017, 95, 81–91. [Google Scholar] [CrossRef] [Green Version]
- Martínez-González, I.; Sánchez-Velázquez, L.R.; Ruiz-Guerra, B.; del Rosario Pineda-López, M.; Velázquez-Rosas, N. The role of seed size in the emergence and survival of seedlings in contrasting environments: The case of Ceiba aesculifolia. New For. 2021, 52, 493–507. [Google Scholar] [CrossRef]
- Vargas-Rodriguez, Y.L.; Vázquez-García, J.A.; Williamson, G.B. Environmental correlates of tree and seedling–sapling distributions in a Mexican tropical dry forest. Plant Ecol. 2005, 180, 117–134. [Google Scholar] [CrossRef]
- Pineda-García, F.; Paz, H.; Meinzer, F.C. Drought resistance in early and late secondary successional species from a tropical dry forest: The interplay between xylem resistance to embolism, sapwood water storage and leaf shedding. Plant Cell Environ. 2013, 36, 405–418. [Google Scholar] [CrossRef]
- Mendoza, P.A.M.; Moya, E.G.; Rivera, J.R.A.; Xolocotzi, E.H. Regeneración natural de especies arbóreas en una selva mediana subperennifolia perturbada por extracción forestal. Acta Bot. Mex. 1995, 32, 11–23. [Google Scholar] [CrossRef] [Green Version]
- Encino-Ruiz, L.; Lindig-Cisneros, R.; Gómez-Romero, M.; Blanco-García, A. Desempeño de tres especies arbóreas del bosque tropical caducifolio en un ensayo de restauración ecológica. Bot. Sci. 2013, 91, 107–114. [Google Scholar] [CrossRef] [Green Version]
- Martínez-González, I.; Velázquez-Rosas, N.; del Rosario Pineda-López, M.; Ruiz-Guerra, B.; Sánchez-Velásquez, L.R. The role of seed size in the performance of Ceiba aesculifolia seedlings subjected to foliar damage. Acta Oecol. 2021, 112, 103772. [Google Scholar] [CrossRef]
- Olvera-Mendoza, E.I.; Lara-Cabrera, S.I.; Sáenz-Romero, C.; Lindig-Cisneros, R. AFLP polymorphism in restored provenances of Ceiba aesculifolia within an urban heat island. Phyton Inter. J. Exp. Bot. 2016, 85, 169–175. [Google Scholar]
- Gómez-Maqueo, X.; Soriano, D.; Velázquez-Rosas, N.; Alvarado-López, S.; Jiménez-Durán, K.; del Mar Garciadiego, M.; Gamboa-deBuen, A. The seed water content as a time-independent physiological trait during germination in wild tree species such as Ceiba aesculifolia. Sci. Rep. 2020, 10, 10429. [Google Scholar] [CrossRef] [PubMed]
- Hending, D.; Randrianarison, H.; Holderied, M.; McCabe, G.; Cotton, S. The kapok tree (Ceiba pentandra (L.) Gaertn, Malvaceae) as a food source for native vertebrate species during times of resource scarcity and its potential for reforestation in Madagascar. Austral Ecol. 2021, 46, 1440–1444. [Google Scholar] [CrossRef]
- Immanuel, R.R.; Ganapathy, M. Growth and physiological attributes of Ceiba pentandra (L.) Gaertn. seeds and seedlings under salt stress. J. Agric. Biol. Sci. 2007, 2, 12–16. [Google Scholar]
- Immanuel, R.R.; Ganapathy, M. Agro-techniques for afforestation of degraded coastal agricultural lands with silk cotton (Ceiba pentendra (L.) Gaertn.). J. Pharmacog. Phytochem. 2019, 8, 1587–1590. [Google Scholar]
- The University of Arizona, Campus Arboretum. 2021. Available online: https://apps.cals.arizona.edu/ (accessed on 26 November 2021).
- Guillot, D.; Laguna, E.; Puche, C.; Meer, P. Ceiba speciosa ‘Valencian Beauty’. Bouteloua 2016, 23, 3–13. [Google Scholar]
- Dick, C.W.; Bermingham, E.; Lemes, M.R.; Gribel, R. Extreme long-distance dispersal of the lowland tropical rainforest tree Ceiba pentandra L.(Malvaceae) in Africa and the Neotropics. Mol. Ecol. 2007, 16, 3039–3049. [Google Scholar] [CrossRef] [PubMed]
- Rahmah, A.U.; Abdullah, M.A. Evaluation of Malaysian Ceiba pentandra (L.) Gaertn. for oily water filtration using factorial design. Desalination 2011, 266, 51–55. [Google Scholar] [CrossRef]
- Silitonga, A.S.; Ong, H.C.; Mahlia, T.M.I.; Masjuki, H.H.; Chong, W.T. Characterization and production of Ceiba pentandra biodiesel and its blends. Fuel 2013, 108, 855–858. [Google Scholar] [CrossRef]
- Zhang, X.; Fu, W.; Duan, C.; Xiao, H.; Shi, M.; Zhao, N.; Xu, J. Superhydrophobicity determines the buoyancy performance of kapok fiber aggregates. Appl. Surf. Sci. 2013, 266, 225–229. [Google Scholar] [CrossRef]
- Graham, B.P.; Haigler, C.H. Microtubules exert early, partial, and variable control of cotton fiber diameter. Planta 2021, 253, 47. [Google Scholar] [CrossRef]
- Carranza-Nuñez, U.; Ramiro Vasquez-Garcia, S.; Flores-Ramirez, N.; Ahmed Abdel-Gawwad, H.; Luis Rico, J.; Arizbe Santiago, A.; Vargas, J.; Cruz-de-León, J. Physicochemical characterization of natural fibers obtained from seed pods of Ceiba aesculifolia. BioResources 2021, 16, 4200–4211. [Google Scholar] [CrossRef]
- Anderson, D.B.; Kerr, T. Growth and structure of cotton fiber. Ind. Eng. Chem. 1938, 30, 48–54. [Google Scholar] [CrossRef]
- Yan, J.; Xu, G.; Wang, F. A study on the quality of kapok blended yarns through different processing methods. J. Tex. Ins. 2013, 104, 675–681. [Google Scholar] [CrossRef]
- Patwary, S. Clothing and Textile Sustainability: Current State of Environmental Challenges and the Ways Forward. Text. Leather Rev. 2020, 3, 158–173. [Google Scholar] [CrossRef]
- Sartika, D.; Syamsu, K.; Warsiki, E.; Fahma, F.; Arnata, I.W. Nanocrystalline cellulose from kapok fiber (Ceiba pentandra) and its reinforcement effect on alginate hydrogel bead. Starch-Stärke 2021, 79, 2100033. [Google Scholar] [CrossRef]
- Leal, M.R.; Flores-Sahagun, T.H.S.; Franco, T.S.; Muniz, G.I. Ceiba speciosa St. Hill fruit fiber as a potential source for nanocellulose production and reinforcement of polyvinyl acetate composites. Polym. Compos. 2021, 42, 397–411. [Google Scholar] [CrossRef]
- Hori, K.; Flavier, M.E.; Kuga, S.; Lam, T.B.T.; Iiyama, K. Excellent oil absorbent kapok Ceiba pentandra (L.) Gaertn. fiber: Fiber structure, chemical characteristics, and application. J. Wood Sci. 2000, 46, 401–404. [Google Scholar] [CrossRef]
- Thilagavathi, G.; Karan, C.P.; Thenmozhi, R. Development and investigations of kapok fiber based needle punched nonwoven as eco-friendly oil sorbent. J. Nat. Fibers 2018, 17, 18–27. [Google Scholar] [CrossRef]
- Nkouam, G.B.; Adjoh, G.A.; Tchankou Leudeu, C.B.; Kouebou, C.; Tchiegang, C.; Kapseu, C. Physico-chemical properties of fruits, seed and oil of kapok (Ceiba pentandra Gaertn.) tree of different provenances from the northern part of Cameroon. Int. J. Agric. Innov. Res. 2017, 6, 275–278. [Google Scholar]
- Berry, S.K. The characteristics of the kapok (Ceiba pentadra Gaertn.) seed oil. Pertanika J. Trop. Agric. Sci. 1979, 21, 1–4. [Google Scholar]
- Rosselli, S.; Tundis, R.; Bruno, M.; Leporini, M.; Falco, T.; Gagliano Candela, R.; Badalamenti, N.; Loizzo, M.R. Ceiba speciosa (A. St.-Hil.) seeds oil: Fatty acids profiling by GC-MS and NMR and bioactivity. Molecules 2020, 25, 1037. [Google Scholar] [CrossRef] [Green Version]
- Avendaño, A.; Casas, A.; Dávila, P.; Lira, R. Use forms, management and commercialization of “pochote” Ceiba aesculifolia (HB & K.) Britten & Baker f. subsp. parvifolia (Rose) PE Gibbs & Semir (Bombacaceae) in the Tehuacán Valley, Central Mexico. J. Arid Environ. 2006, 67, 15–35. [Google Scholar] [CrossRef]
- Avendaño, A.; Casas, A.; Dávila, P.; Lira, R. In situ management and patterns of morphological variation of Ceiba aesculifolia subsp. parvifolia (Bombacaceae) in the Tehuacán-Cuicatlán Valley. Econ. Bot. 2009, 63, 138–151. [Google Scholar] [CrossRef]
- Suastegui-Baylón, L.; Salazar, R.; Maldonado-Astudillo, Y.I.; Ramírez-Sucre, M.O.; Arámbula-Villa, G.; Flores-Casamayor, V.; Jiménez-Hernández, J. Physical, chemical and rheological characterization of tuber and starch from Ceiba aesculifolia subsp. parvifolia. Molecule 2021, 26, 2097. [Google Scholar] [CrossRef] [PubMed]
- Marks, R.A.; Hotaling, S.; Frandsen, P.B.; VanBuren, R. Representation and participation across 20 years of plant genome sequencing. Nat. Plants 2021, 7, 1571–1578. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Wang, H.; Liu, C.; Chu, H.; Dai, D.; Song, S.; Yu, L.; Han, L.; Fu, Y.; Tian, B.; et al. De novo genome assembly of the red silk cotton tree (Bombax ceiba). GigaScience 2018, 7, giy051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Y.; Wang, H.; Liu, C.; Chu, H.; Yan, Y.; Tang, L. Complete chloroplast genome sequence of the red silk cotton tree (Bombax ceiba). Mitochondrial DNA Part B 2018, 3, 315–316. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Wang, H.; Liu, C.; Chu, H.; Yan, Y.; Tang, L. The complete mitochondrial genome of Bombax ceiba. Mitochondrial DNA Part B 2018, 3, 313–314. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Fan, W.; Guo, X.; Wu, K.; Zhou, S.; Chen, Z.; Li, D.; Wang, K.; Zhu, Y.; Zhou, Y. MaGenDB: A functional genomics hub for Malvaceae plants. Nucleic Acids Res. 2020, 48, D1076–D1084. [Google Scholar] [CrossRef]
- Huang, S.; Zhu, Q.; Huang, G.; Han, B.; Zhou, Q.; Dai, J. The chloroplast genome of silk floss tree (Ceiba speciosa). Mitochondrial DNA Part B 2019, 4, 3632–3633. [Google Scholar] [CrossRef] [Green Version]
- Nakabayashi, K.; Okamoto, M.; Koshiba, T.; Kamiya, Y.; Nambara, E. Genome-wide profiling of stored mRNA in Arabidopsis thaliana seed germination: Epigenetic and genetic regulation of transcription in seed. Plant J. 2005, 41, 697–709. [Google Scholar] [CrossRef] [PubMed]
- Howell, K.A.; Narsai, R.; Carroll, A.; Ivanova, A.; Lohse, M.; Usadel, B.; Miller, A.H.; Whelan, J. Mapping metabolic and transcript temporal switches during germination in rice highlights specific transcription factors and the role of RNA instability in the germination process. Plant Physiol. 2009, 149, 961–980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, G.; Huang, J.Q.; Chen, X.Y.; Zhu, Y.X. Recent advances and future perspectives in cotton research. Annu. Rev. Plant Biol. 2021, 72, 437–462. [Google Scholar] [CrossRef]
- Wu, H.; Tian, Y.; Wan, Q.; Fang, L.; Guan, X.; Chen, J.; Hu, Y.; Ye, W.; Zhang, H.; Guo, W.; et al. Genetics and evolution of MIXTA genes regulating cotton lint fiber development. New Phytol. 2018, 217, 883–895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-Maqueo, X.M.; Gamboa-deBuen, A.; (Universidad Nacional Autónoma de México, Ciudad de México, México). Personal communication, 2022.
- Arellanes-Cancino, Y.; Romero-Sosa, M.Á.; Vega, E.; Maza-Villalobos, S.; Casas-Fernández, A. Ecological bases for sustainable management of Pochote (Ceiba aesculifolia subsp. parvifolia) through demographic analysis. Econ. Bot. 2018, 72, 20–37. [Google Scholar] [CrossRef]
- Royal Botanic Gardens Kew. Plants of the World Online Database. Available online: https://powo.science.kew.org/ (accessed on 4 January 2022).
- Tropicos.org. Missouri Botanical Garden. Version 3.3.1.. Available online: https://tropicos.org (accessed on 5 January 2022).




{kind=link}
{kind=link}
{kind=link}
| Species | Height | Flower Features | Trunk | Habitat | Distribution | Flowering Time | Pollinators |
|---|---|---|---|---|---|---|---|
| C. aesculifolia (Kunth) Britten and Baker f. | tree, 8–15 m [16] | cream, ivory | slender trunk with spines | SDTF | Mexico and Central America | March–July (September–January) | bats |
| C. boliviana Britten and Baker f. | tree, 10 m | pink with intense dark red striations | usually ventricose with spines | SDTF | Bolivia, Peru | March–April(January) | possibly bats |
| C. chodatii (Hassl.) Ravenna | tree, 12 m | ivory to pale yellow, sometimes with crimson flecks | ventricose, usually with spines | SDTF | Argentina, Bolivia and Paraguay | February–May | possibly sphingid moths |
| C. crispiflora (Kunth) Ravenna | tree, over 10 m | dark pink magenta with few striations distally, yellowish at the base | usually with spines | SDTF and humid forests | Brazil | February–March | possibly diurnal butterflies |
| C. erianthos (Cav.) K. Schum. | tree, 10 m | white with sparse carmine striations distally, becoming uniform towards the base | presents spines | SDTF | Brazil | March–July | bats |
| C. glaziovii (Kuntze) K. Schum. | tree, 10–15 m | white, sometimes with magenta striations towards the base | ventricose, with spines | SDTF | Brazil | July-September | possibly bats |
| C. insignis (Kunth) P.E. Gibbs and Semir | tree, 10 m | white to light pink with a yellowish base, occasionally with red striations | ventricose, usually with spines | SDTF | Ecuador and Peru | May–July (October) | possibly bats |
| C. jasminodora (A. St.-Hil.) K. Schum. | treelet, 1–2 m | cream, markedly reflexed | spiny branches | SDTF rocky outcrops [13] | Brazil | April–July | possibly moths |
| C. lupuna P.E. Gibbs and Semir | giant tree, up to 50 m | deep red distally, pale yellow with red speckles towards the base | usually with spines | humid forests | Ecuador, Peru and Brazil | May–June | unknown |
| C. pentandra (L.) Gaertn. | giant tree, up to 60 m; the savannah ecotype about 10 m; cultivated types 25 m [5] | white to distinctive light pink | presents spines and large buttresses (cultivated types can lack both) [5] | SDTF and humid forests [13] | pantropical, introduced in Asia and Europe | August–September | bats, possibly bees |
| C. pubiflora (A. St.-Hil.) | Tree, over 20 m | pale pink with sparse dark flecks, or pink-lilac with carmine striations | sometimes ventricose, with spines | SDTF | Paraguay, Argentina and Brazil | February–May | possibly hummingbirds |
| C. rubriflora Carv.-Sobr. and L.P. Queiroz | tree, 20 m | deep red | ventricose, with spines | SDTF, calcareous outcrops | Brazil | July–August | unknown |
| C. samauma (Mart.) K. Schum. | tree, 15 m | white but with dense golden brown trichomes | may present buttresses, spiny branches | SDTF and humid forests | Ecuador, Peru, Brazil and Bolivia | December–March (May) | unknown |
| C. schottii Britten and Baker f. | tree, 8 m | white | presents spines | SDTF, mangroves and flood zones [17] | Mexico and Central America | June–October [17] | diurnal butterflies |
| C. soluta (Donn. Sm.) Ravenna | Not reported | white | presents spines | SDTF | Guatemala | February | unknown |
| C. speciosa (A. St.-Hil.) Ravenna | tree, 10–20 m | dark pink magenta distally, base white to yellow, usually with dark striations | ventricose, usually with spines | SDTF and humid forests | Argentina, Bolivia and Brazil | January–May | possibly diurnal butterflies |
| C. trischistandra (A. Gray) Bakh. | tree, 15–30 m | white, externally tinted green, somewhat reflexed | presents spines | SDTF | Ecuador | April-July | unknown |
| C. ventricosa (Nees and Mart.) Ravenna | tree, 10 m or more | white to cream with dark redish flecks towards the base | ventricose, usually with spines | SDTF | Brazil | February-April | possibly bats |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gómez-Maqueo, X.; Gamboa-deBuen, A. The Biology of the Genus Ceiba, a Potential Source for Sustainable Production of Natural Fiber. Plants 2022, 11, 521. https://doi.org/10.3390/plants11040521
Gómez-Maqueo X, Gamboa-deBuen A. The Biology of the Genus Ceiba, a Potential Source for Sustainable Production of Natural Fiber. Plants. 2022; 11(4):521. https://doi.org/10.3390/plants11040521
Chicago/Turabian StyleGómez-Maqueo, Ximena, and Alicia Gamboa-deBuen. 2022. "The Biology of the Genus Ceiba, a Potential Source for Sustainable Production of Natural Fiber" Plants 11, no. 4: 521. https://doi.org/10.3390/plants11040521