
Effect of Phytoplasma Associated with Sesame Phyllody on Ultrastructural Modification, Physio-Biochemical Traits, Productivity and Oil Quality




Abstract
:1. Introduction
2. Results
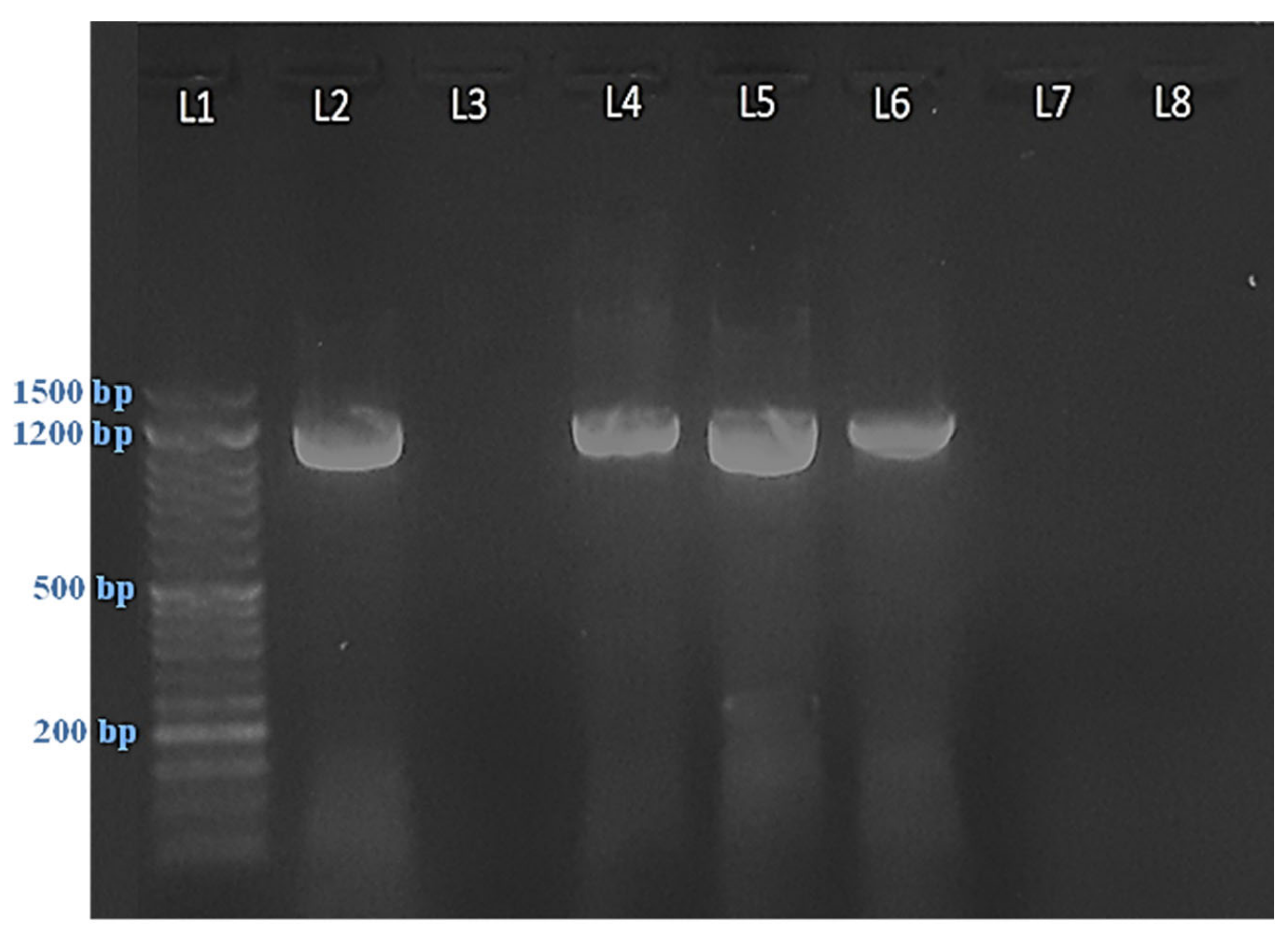
2.1. Symptomatology and Phytoplasma Detection
2.2. Sequence Analysis
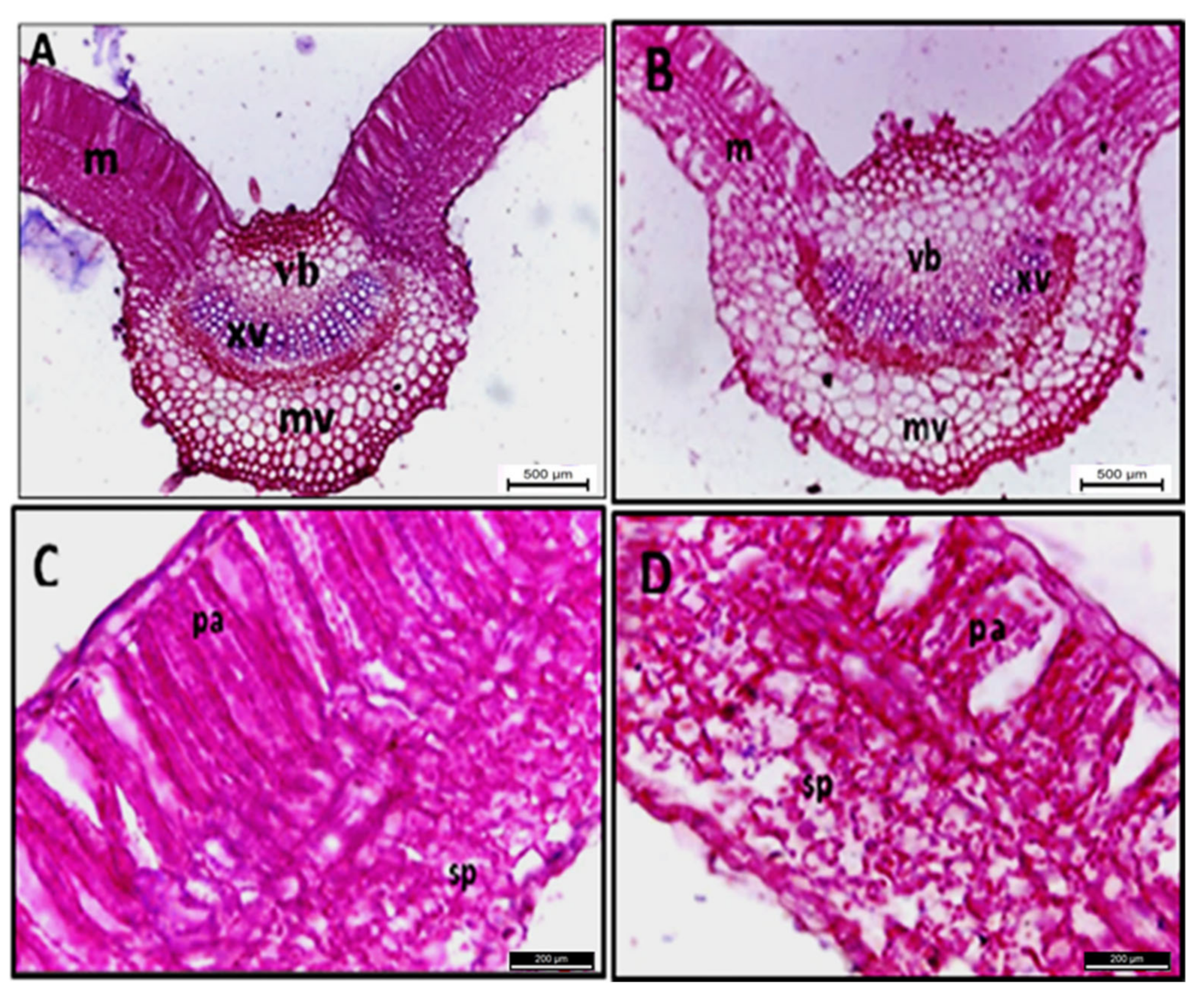
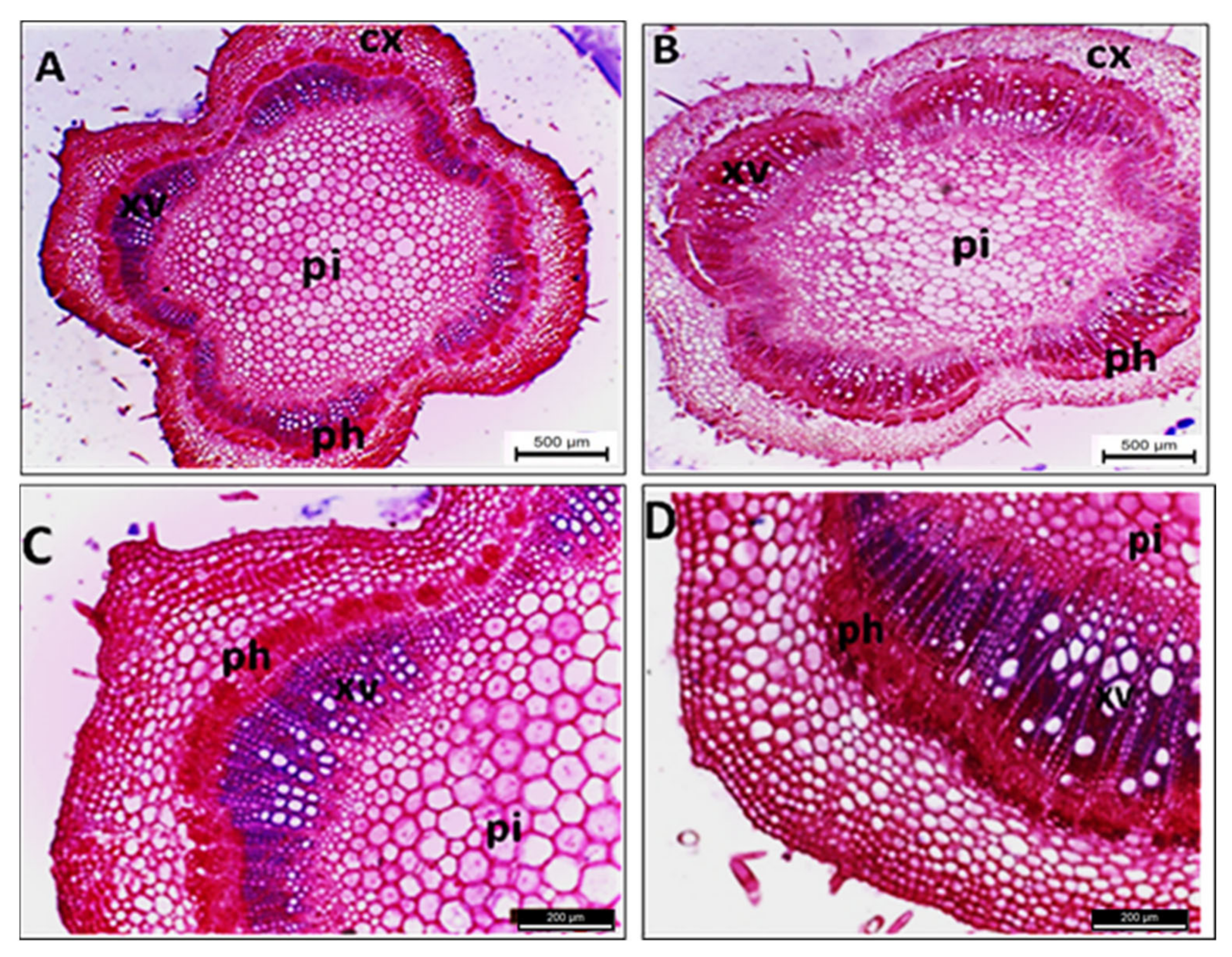
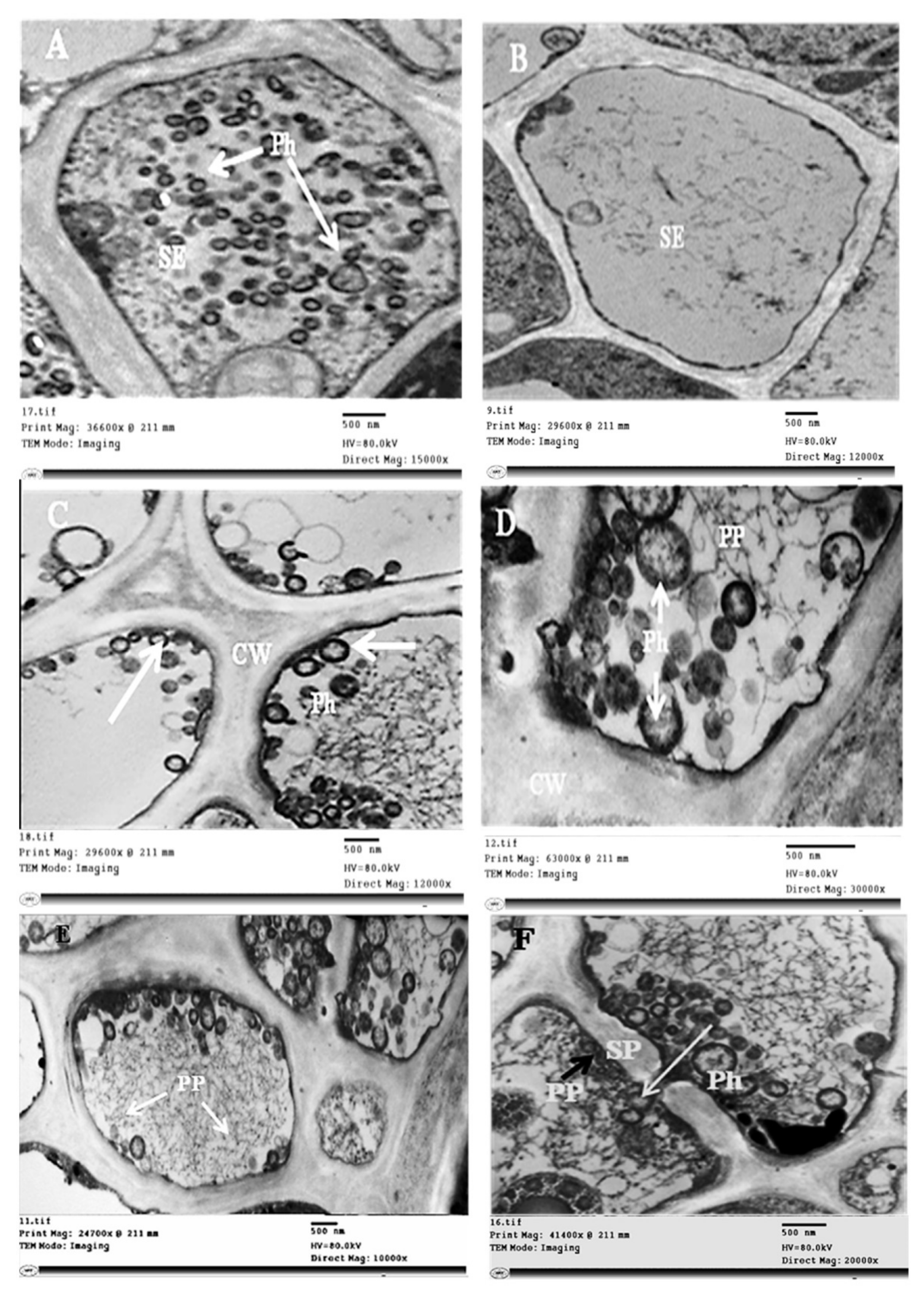
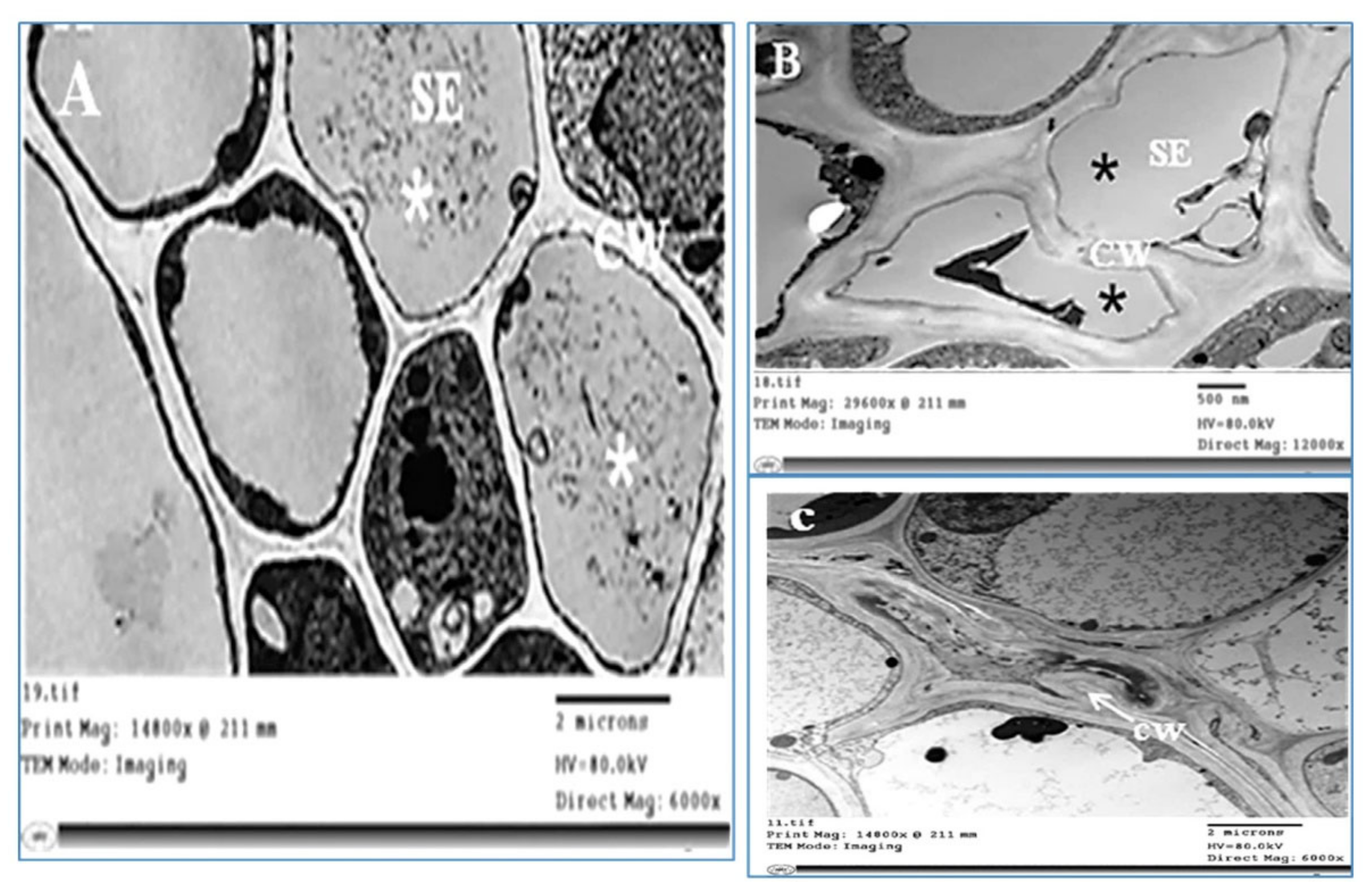
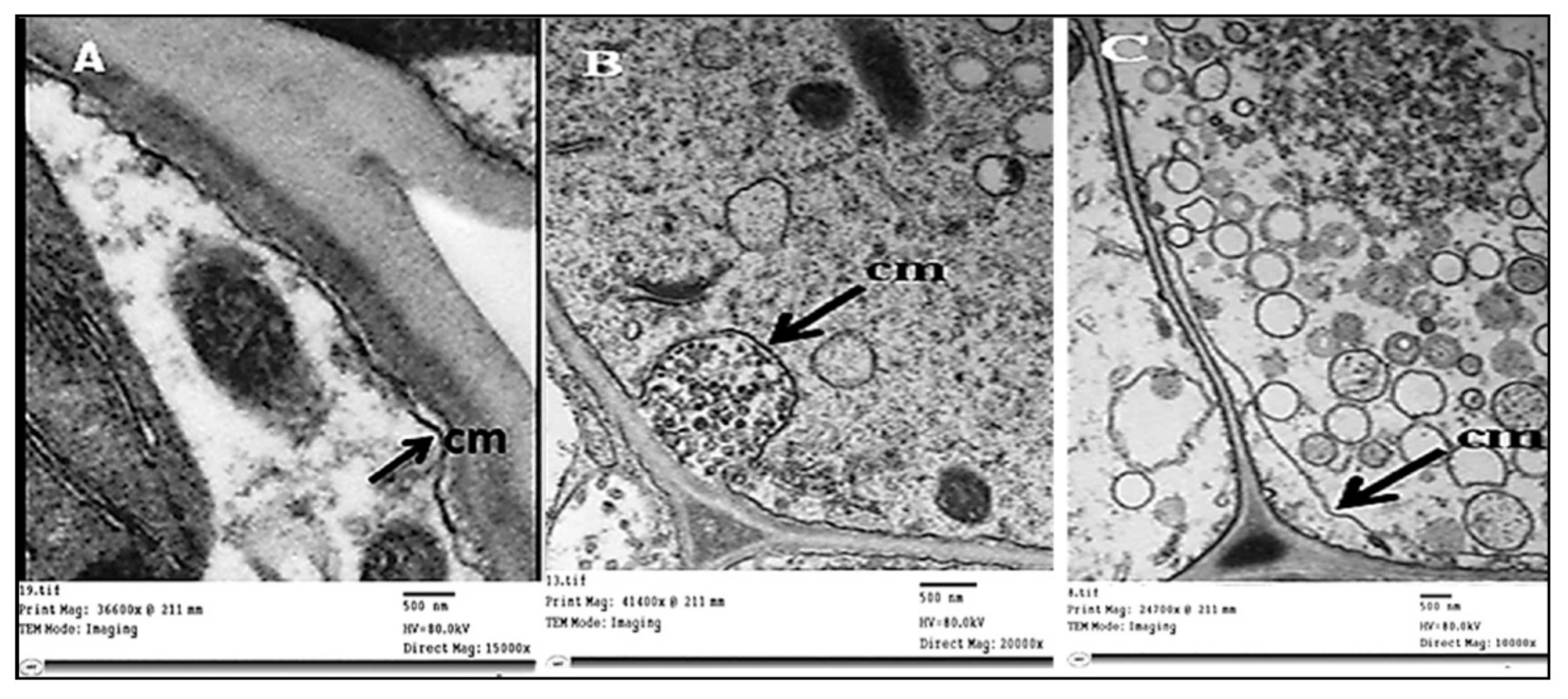
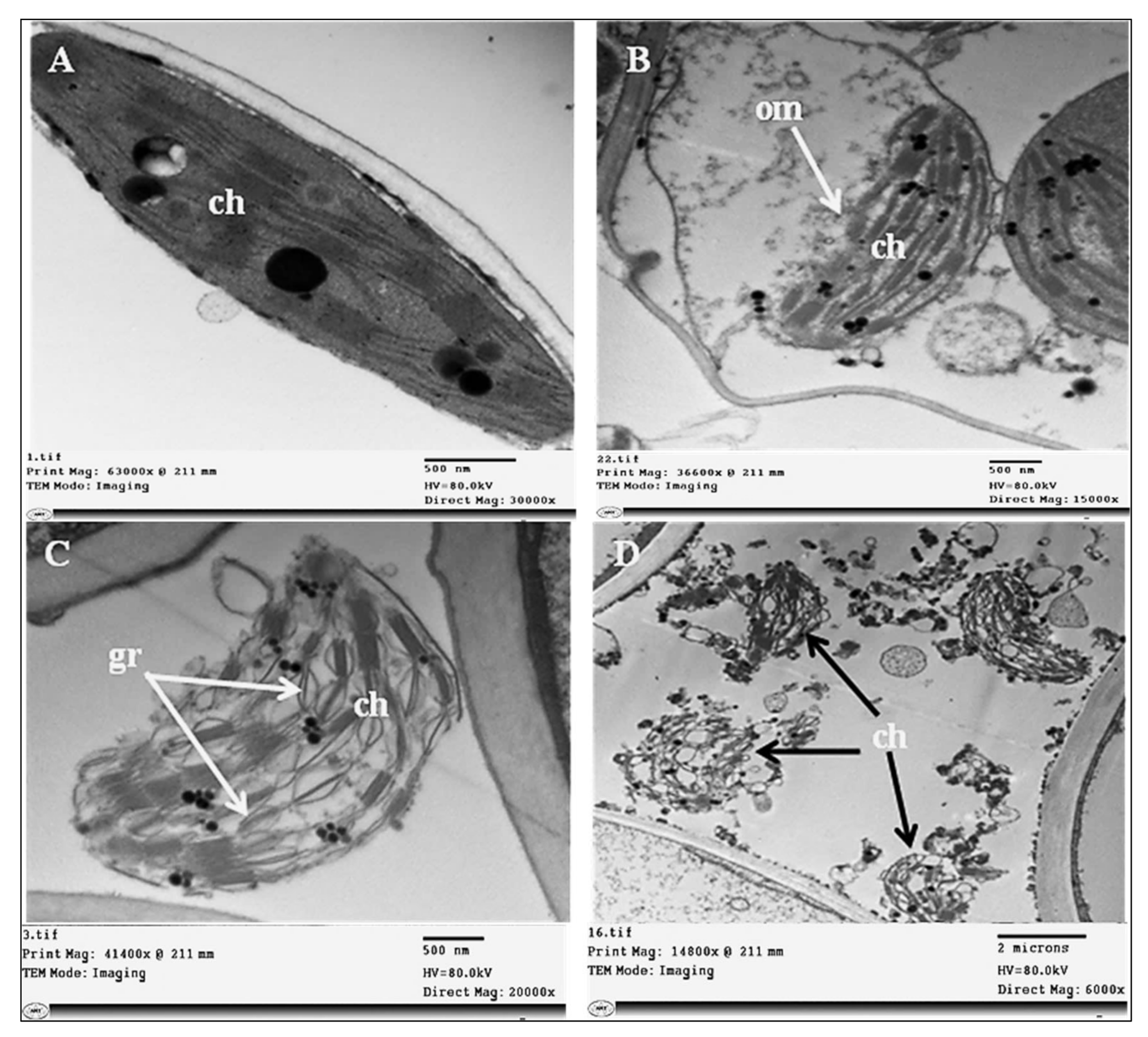
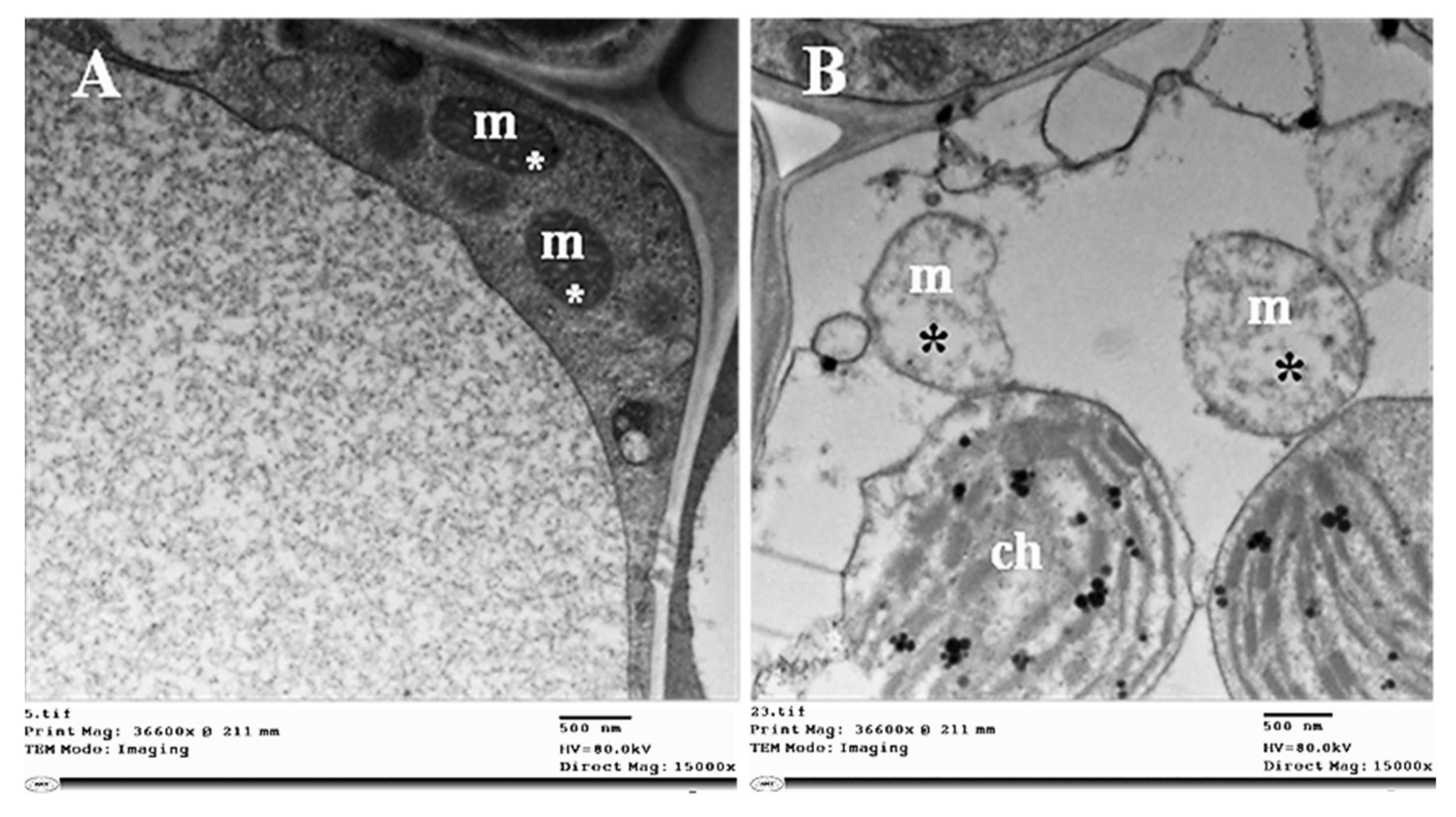
2.3. Histopathological Changes
2.4. Changes in Phytoplasma-Related Primary and Secondary Metabolites
2.5. Morpho-Physiological Responses in Phytoplasma-Infected Sesame Plants
2.6. Changes in Productivity, Seed Harvest Index, Seed Protein, and Seed Oil
2.7. Physio-Chemical Changes in Sesame Oil Properties
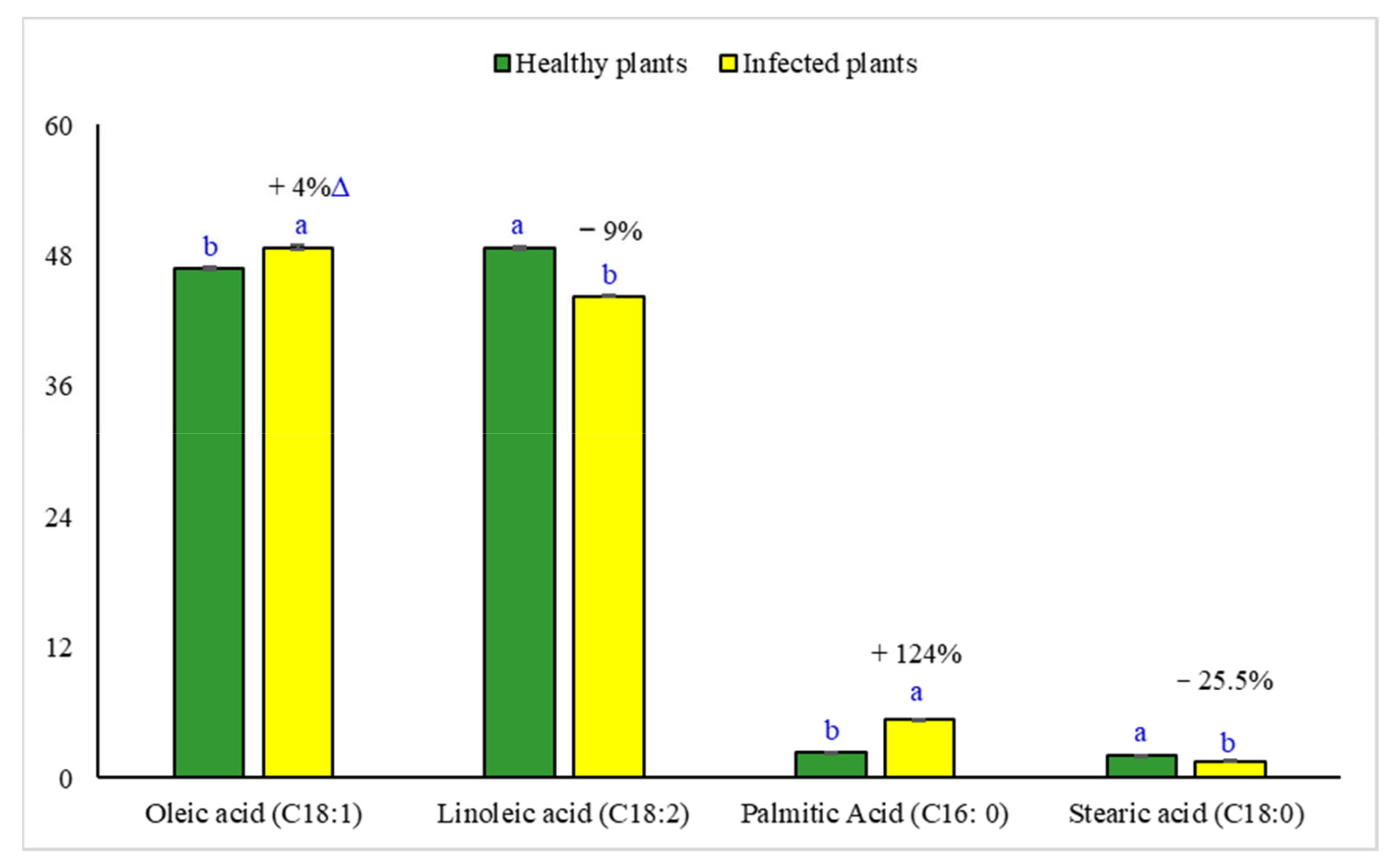
2.8. Changes in the Fatty Acid Composition of Sesame Oil
2.9. Correlation Analysis
3. Discussion
4. Materials and Methods
4.1. Site Description, Experimental Details, and Plant Material
4.2. Source of Samples
4.3. Nucleic Acid Extraction
4.4. Polymerase Chain Reaction (PCR)
4.5. PCR Cleanup
4.6. Sequencing and Analysis
4.7. Sampling and Measurements
4.7.1. Histopathological Changes
4.7.2. Light Microscopy
4.7.3. Transmission Electron Microscopy
4.7.4. Determination of Phytoplasma-Related Primary and Secondary Metabolites
4.7.5. Morpho-Physiological Responses
4.7.6. Yield and Its Components and Seed Harvest Index
4.7.7. Total Seed Protein, Seed Oil Content, and Oil Yield
4.7.8. Sesame Oil Extraction and Oil Physio-Chemical Properties Determination
4.7.9. Quantification of Sesame oil Fatty Acid Composition
4.7.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kabak, B.; Dobson, A.D. Mycotoxins in spices and herbs—An update. Crit. Rev. Food Sci. Nutr. 2017, 57, 18–34. [Google Scholar] [CrossRef] [PubMed]
- Jayaraj, P.; Narasimhulu, C.A.; Rajagopalan, S.; Parthasarathy, S.; Desikan, R. Sesamol: A powerful functional food ingredient from sesame oil for cardioprotection. Food Funct. 2020, 11, 1198–1210. [Google Scholar] [CrossRef] [PubMed]
- Uzun, B.; Arslan, C.; Furat, S. Variation in fatty acid composition, oil content and oil yield in a germplasm collection of sesame (Sesamum indicum L.). J. Am. Oil Chem. Soc. 2008, 85, 1135–1142. [Google Scholar] [CrossRef]
- Rizki, H.; Nabloussi, A.; Kzaiber, F.; Jbilou, M.; Latrache, H.; Hanine, H. Comparative assessment of bioactive components, antioxidant effects from 15 cultivars of sesame (Sesamum indicum L.) for different crop years. Int. J. Sci. Eng. Res. 2018, 9, 462–468. [Google Scholar]
- Nzikou, J.M.; Matos, L.; Kalou, G.B.; Ndangui, C.B.; Pambou-Tobi, N.P.G.; Kimbonguila, A.; Silou, T.; Linder, M.; Desobry, S. Chemical composition on the seeds and oil of sesame (Sesamum indicum L.) grown in Congo-Brazzaville. Adv. J. Food Sci. Technol. 2009, 1, 6–11. [Google Scholar]
- Food and Agriculture Organization of the United Nations (FAO). FAOSTAT Statistics; United Nations: Rome, Italy, 2020. [Google Scholar]
- MALR. Bulletin of Agricultural Statistics, Arab Republic of Egypt; Ministry of Agriculture and Land Reclamation, the Egyptian Economic Affairs Sector: Dokki, Egypt, 2020.
- FAS. Foreign Agriculture Service/USDA; Circular Series FOP 9–2017; Office of Global Analysis: Washington, DC, USA, 2017.
- Youssef, S.A.; Safwat, G.; Shalaby, A.B.A.; El-Beltagi, H.S. Effect of phytoplasma infection on plant hormones, enzymes and their role in infected sesame. Fresenius Environ. Bull. 2018, 27, 5727–5735. [Google Scholar]
- Akhtar, K.P.; Dickinson, M.; Sarwar, G.; Jamil, F.F.; Haq, M.A. First report on the association of a 16SrII phytoplasma with sesame phyllody in Pakistan. Plant Pathol. 2008, 57, 771. [Google Scholar] [CrossRef]
- Nagaraju; Muniyappa, V. Viral and phytoplasma diseases of sesame. In Diseases of Oilseed Crops; Saharan, G.S., Mehta, N., Sangwan, M.S., Eds.; Indus Publishing Company: New Delhi, India, 2005; pp. 304–318. [Google Scholar]
- Akhtar, K.P.; Sarwar, G.; Sarwar, N.; Elahi, M.T. Field evaluation of sesame germplasm against sesame phyllody disease. Pak. J. Bot. 2013, 45, 1085–1090. [Google Scholar]
- Salehi, M.; Esmailzadeh-Hosseini, S.A.; Salehi, E.; Bertaccini, A. Genetic diversity and vector transmission of phytoplasmas associated with sesame phyllody in Iran. Folia Microbiol. 2017, 62, 99–109. [Google Scholar] [CrossRef]
- Vasudeva, R.S.; Sahambi, H.S. Phyllody in sesamum (Sesamum orientale L.). Indian Phytopathol. 1955, 8, 124–129. [Google Scholar]
- Turkmenoglu, Z.; Ari, U. A disease-phyllody virus noted on sesame in the Aegean region. Plant Prot. Bull. 1959, 1, 12–17. [Google Scholar]
- Caglayan, K.; Gazel, M.; Škorić, D. Transmission of phytoplasmas by agronomic practices. In Phytoplasmas: Plant Pathogenic Bacteria—II; Bertaccini, A., Weintraub, P., Rao, G., Mori, N., Eds.; Springer: Singapore, 2019; pp. 149–163. [Google Scholar] [CrossRef]
- El-Banna, O.M.; Mikhail, M.S.; Farag, A.G.; Mohamed, A.M.S. Detection of phytoplasma in tomato and pepper plants by electron microscope and molecular biology-based methods. Egypt. J. Virol. 2007, 4, 93–111. [Google Scholar]
- Ahmed, E.A.; Shalaby, O.Y.; Dwidar, E.F.; Mokbel, S.A.; El-Attar, A.K. Occurrence, etiology and molecular characterization of phytoplasma diseases on Solanum lycopersicum crop in Egypt. Egypt. J. Virol. 2014, 11, 244–261. [Google Scholar]
- Akhtar, K.P.; Dickinson, M.; Sarwar, G.; Mushtak, A. Sesame phyllody disease: Its symptomatology, etiology and transmission in Pakistan. Turk. J. Agric. For. 2009, 33, 477–486. [Google Scholar] [CrossRef]
- El-Banna, O.M.; Mikhail, M.S.; El-Attar, A.K.; Aljamali, A.A.R. Molecular and electron microscope evidence for an association of phytoplasma with sesame phyllody in Egypt. Egypt. J. Phytopathol. 2013, 41, 1–14. [Google Scholar] [CrossRef]
- Ahmad, S.J.; Farid, N.; Ahmad, J.N. Metabolic and physiological changes induced in Sesamum indicum infected by phytoplasmas. Phytopathog. Mollicutes 2019, 9, 137–138. [Google Scholar] [CrossRef]
- Singh, A.; Verma, P.; Lakhanpaul, S. Exploring the methylation status of selected flowering genes in healthy and phyllody infected sesame plants. Phytopathog. Mollicutes 2020, 10, 36–42. [Google Scholar] [CrossRef]
- Xue, C.; Liu, Z.; Dai, L.; Bu, J.; Liu, M.; Zhao, Z.; Jiang, Z.; Gao, W.; Zhao, J. Changing host photosynthetic, carbohydrate, and energy metabolisms play important roles in Phytoplasma infection. Phytopathology 2018, 108, 1067–1077. [Google Scholar] [CrossRef] [Green Version]
- Maust, B.E.; Espadas, F.; Talavera, C.; Aguilar, M.; Santamaría, J.M.; Oropeza, C. Changes in carbohydrate metabolism in coconut palms infected with the lethal yellowing phytoplasma. Phytopathology 2003, 93, 976–981. [Google Scholar] [CrossRef] [Green Version]
- Tan, Y.; Wei, H.R.; Wang, J.W.; Zong, X.J.; Zhu, D.Z.; Liu, Q.Z. Phytoplasmas change the source-sink relationship of field-grown sweet cherry by disturbing leaf function. Physiol. Mol. Plant Pathol. 2015, 92, 22–27. [Google Scholar] [CrossRef]
- Devonshire, B.J. Visualization of phytoplasmas using electron microscopy. In Phytoplasma. Methods in Molecular Biology (Methods and Protocols); Dickinson, M., Hodgetts, J., Eds.; Humana Press: Totowa, NJ, USA, 2013; Volume 938. [Google Scholar] [CrossRef]
- El-Banna, O.M.; Toima, N.I.; Youssef, S.A.; Shalaby, A. Molecular and electron microscope evidence for an association of phytoplasma with Citrus Witches Broom disease. Int. J. Sci. Eng. Res. 2015, 6, 127–133. [Google Scholar]
- Youssef, S.A.; Sayed, Y.; Hassan, O.S.; Safwat, G.; Shalaby, A. Universal and specific 16S-23Sr RNA PCR primers for identification of phytoplasma associated with sesame in Egypt. Int. J. Adv. Res. Biol. Sci. 2017, 4, 191–200. [Google Scholar] [CrossRef]
- Gundersen, D.E.; Lee, I.M.; Rehner, S.A.; Davis, R.E.; Kingsbury, D.T. Phylogeny of mycoplasma like organisms (phytoplasmas): A basis for their classification. J. Bacteriol. 1994, 176, 5244–5254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mekdad, A.A.A.; Shaaban, A.; Rady, M.M.; Ali, E.F.; Hassan, F.A.S. Integrated application of K and Zn as an avenue to promote sugar beet yield, industrial sugar quality, and K-use efficiency in a salty and semi-arid agro-ecosystem. Agronomy 2021, 11, 780. [Google Scholar] [CrossRef]
- Shaaban, A.; Al-Elwany, O.A.A.I.; Abdou, N.M.; Hemida, K.A.; El-Sherif, A.M.A.; Abdel-Razek, M.A.; Semida, W.M.; Mohamed, G.F.; Abd El-Mageed, T.A. Filter Mud Enhanced Yield and Soil Properties of Water-Stressed Lupinus termis L. in Saline Calcareous Soil. J. Soil Sci. Plant Nutr. 2022. [Google Scholar] [CrossRef]
- Mekdad, A.A.A.; El-Sherif, A.M.A.; Rady, M.M.; Shaaban, A. Culture management and application of humic acid in favor of Helianthus annuus L. oil yield and nutritional homeostasis in a dry environment. J. Soil Sci. Plant Nutr. 2021. [Google Scholar] [CrossRef]
- Abd El-Mageed, T.A.; Rady, M.O.; Semida, W.M.; Shaaban, A.; Mekdad, A.A. Exogenous micronutrients modulate morpho-physiological attributes, yield, and sugar quality in two salt-stressed sugar beet cultivars. J. Soil Sci. Plant Nutr. 2021, 21, 1421–1436. [Google Scholar] [CrossRef]
- Mekdad, A.A.; El-Enin, M.M.A.; Rady, M.M.; Hassan, F.A.; Ali, E.F.; Shaaban, A. Impact of level of nitrogen fertilization and critical period for weed control in peanut (Arachis hypogaea L.). Agronomy 2021, 11, 909. [Google Scholar] [CrossRef]
- Singh, V.; Kumar, S.; Lakhanpaul, S. Differential distribution of phytoplasma during phyllody progression in sesame (Sesamum indicum L.) under field conditions—An important consideration for effective sampling of diseased tissue. Crop Prot. 2018, 110, 288–294. [Google Scholar] [CrossRef]
- Hemmati, C.; Nikooei, M.; Al-Subhi, A.M.; Al-Sadi, A.M. History and current status of phytoplasma diseases in the Middle East. Biology 2021, 10, 226. [Google Scholar] [CrossRef]
- Junior, E.D.J.G.; Segnana, L.G.; Kitajima, E.W.; Bedendo, I.P. Sesame phyllody associated with a 16SrI-B phytoplasma, a ‘Candidatus Phytoplasma asteris’-related strain, in Paraguay. Sci. Agric. 2019, 76, 47–50. [Google Scholar] [CrossRef]
- Baspinar, H.; Korkmaz, S.; Onelge, N.; Cinar, A.; Uygun, N.; Kersting, U. Studies on citrus stubborn disease pathogen and sesame phyllody in sesame and their related leafhopper vector. J. Turk. Phytopathol. 1993, 22, 1–8. [Google Scholar]
- Venkataravanappa, V.; Reddy, C.N.L.; Manjunath, M.; Chauhan, N.S.; Reddy, M.K. Detection, characterization and in-silico analysis of candidatus phytoplasma australasia associated with phyllody disease of sesame. Adv. Plants Agric. Res. 2017, 7, 00256. [Google Scholar] [CrossRef] [Green Version]
- Smart, C.D.; Schneider, B.; Blomquist, C.L.; Guerra, L.J.; Harrison, N.A.; Ahrens, U.; Lorenz, K.H.; Seemüller, E.; Kirkpatrick, B.C. Phytoplasma-specific PCR primers based on sequences of the 16S-23S rRNA spacer region. Appl. Environ. Microbiol. 1996, 62, 2988–2993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marzachì, C. Molecular diagnosis of phytoplasmas. Phytopathol. Mediterr. 2004, 43, 228–231. [Google Scholar]
- Kakizawa, S.; Yoneda, Y. The role of genome sequencing phytoplasma research. Phytopathog. Mollicutes 2015, 5, 19–24. [Google Scholar] [CrossRef]
- Omar, A.F.; Foissac, X. Occurrence and incidence of phytoplasmas of the 16SrII-D subgroup on solanaceous and cucurbit crop in Egypt. Eur. J. Plant Pathol. 2012, 133, 353–360. [Google Scholar] [CrossRef]
- El-Sisi, Y.; Omar, A.F.; Sidaros, S.A.; ElSharkawy, M.M. Characterization of 16SrII-D subgroup associated phytoplasmas in new host plants in Egypt. Arch. Phytopathol. Plant Prot. 2017, 50, 504–513. [Google Scholar] [CrossRef]
- Gad, S.M.; Kheder, A.A.; Awad, M.A. Detection and molecular identification of phytoplasma associated with Gazania in Egypt. J. Virol. Sci. 2019, 6, 12–23. [Google Scholar]
- El-Banna, O.M.; El-Deeb, S.H. Phytoplasma associated with mango malformation disease in Egypt. J. Phytopathol. 2007, 35, 141–153. [Google Scholar]
- Randall, J.J.; Bosland, P.W.; Hanson, S.F. Brote grande, a new phytoplasma associated diseases of chile peppers in the desert southwest. Plant Health Prog. 2011, 12, 18. [Google Scholar] [CrossRef]
- Ahmed, E.A.; Shalaby, O.Y.; Dwidar, E.F.; Mokbel, S.A.; El-Attar, A.K. Ultrastructural changes in tomato plant induced by phytoplasma infection and attempts for its elimination using tissue culture techniques. Egypt. J. Virol. 2016, 13, 34–51. [Google Scholar]
- Christensen, N.M.; Axelsen, K.B.; Nicolaisen, M.; Schulz, A. Phytoplasmas and their interactions with hosts. Trends Plant Sci. 2005, 10, 526–535. [Google Scholar] [CrossRef] [PubMed]
- Mou, H.Q.; Lu, J.; Zhu, S.F.; Lin, C.L.; Tian, G.Z.; Xu, X.; Zhao, W.J. Transcriptomic analysis of paulownia infected by paulownia witches’ broom phytoplasma. PLoS ONE 2013, 8, e77217. [Google Scholar] [CrossRef] [PubMed]
- Kesumawati, E.; Kimata, T.; Uemachi, T.; Hosokawa, M.; Yazawa, S. Correlation of phytoplasma concentration in hydrangea macrophylla with green-flowering stability. Sci. Hortic. 2006, 108, 74–78. [Google Scholar] [CrossRef]
- Kaminiska, M.; Sliwa, H.; Rudzinska, L.; Angwald, A. The association of phytoplasma with stunting, leaf necrosis and witches-broom symptoms in magnolia plants. J. Phytopathol. 2001, 149, 719–724. [Google Scholar] [CrossRef]
- Anstead, J.; Froelich, D.; Knoblauch, M.; Thompson, G. Arabidopsis P-protein filament formation requires both AtSEOR1 an AtSEOR2. Plant Cell Physiol. 2012, 53, 1033–1042. [Google Scholar] [CrossRef] [Green Version]
- Pagliari, L.; Buoso, S.; Santi, S.; Furch, A.C.U.; Martini, M.; Degola, F.; Loschi, A.; van Bel, A.J.E.; Musetti, R. Filamentous sieve element proteins are able to limit phloem mass flow but not phytoplasma spread. J. Exp. Bot. 2017, 13, 3673–3688. [Google Scholar] [CrossRef]
- Bernardini, C.; Pagliari, L.; de Rosa, V.; Almeida-Trapp, M.; Santi, S.; Martini, M.; Buoso, S.; Loschi, A.; Loi, N.; Chiesa, F.; et al. Pre-symptomatic modified phytohormone profile is associated with lower phytoplasma titres in an Arabidopsis seor1ko line. Sci. Rep. 2020, 10, 14770. [Google Scholar] [CrossRef]
- Buxa, S.V.; Degola, F.; Polizzotto, R.; Marco, F.D.; Loschi, A.; Kogel, K.H.; Musetti, R. Phytoplasma infection in tomato is associated with re-organization of plasma membrane, ER stacks, and actin filaments in sieve elements. Front Plant Sci. 2015, 6, 650. [Google Scholar] [CrossRef] [Green Version]
- Santi, S.; de Marco, F.; Polizzotto, R.; Grisan, S.; Musetti, R. Recovery from stolbur disease in grapevine involves changes in sugar transport and metabolism. Front. Plant Sci. 2013, 4, 171–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hameed, S.; Akhtar, K.P.; Hameed, A.; Gulzar, T.; Kiran, S.; Yousaf, S.; Abbas, G.; Asghar, M.J.; Sarwar, N. Biochemical changes in the leaves of mungbean (Vigna radiata) plants infected by phytoplasma. Turk. J. Biochem. 2017, 42, 591–599. [Google Scholar] [CrossRef]
- Thangjam, R.; Vastrad, A.S. Biochemical analysis of phytoplasma infected sesame plant transmitted by Orosius albicinctus distant. J. Entomol. Zool. Stud. 2018, 6, 153–155. [Google Scholar]
- Rasool, A.; Jahan, M.S.; Shazad, U.; Tariq, A.; Calica, P.N. Effect of phytoplasma infection on primary and secondary metabolites and antioxidative enzyme activities of sweet orange (Citrus sinenses L.). J. Plant Pathol. Microbiol. 2020, 11, 519. [Google Scholar] [CrossRef]
- Agrios, G.N. Plant Pathology, 4th ed.; Academic Press: San Diego, CA, USA, 1997. [Google Scholar]
- Tornero, P.; Chao, R.; Luthin, W.; Goff, S.; Dangl, J. Large-scale structure, function, analysis, of Arabidopsis RPM1 disease resistance protein. Plant Cell. 2002, 14, 435–450. [Google Scholar] [CrossRef] [Green Version]
- Junqueira, A.; Bedendo, I.; Pascholati, S. Biochemical changes in corn plants infected by the maize bushy stunt phytoplasma. Physiol. Mol. Plant Pathol. 2004, 65, 181–185. [Google Scholar] [CrossRef]
- Reveles-Torres, L.R.; Velásquez-Valle, R.; Salas-Muñoz, S.; Mauricio-Castillo, J.A.; Esqueda-Dávila, K.C.J.; Herrera, M.D. Candidatus phytoplasma trifolii (16Sr VI) infection modifies the polyphenols concentration in pepper (Capsicum annuum) plant tissues. J. Phytopathol. 2018, 166, 555–564. [Google Scholar] [CrossRef]
- Negro, C.; Sabella, E.; Nicolì, F.; Pierro, R.; Materazzi, A.; Panattoni, A.; Aprile, A.; Nutricati, E.; Vergine, M.; Miceli, A.; et al. Biochemical changes in leaves of vitis vinifera cv. Sangiovese infected by Bois Noir phytoplasma. Pathogens 2020, 9, 269. [Google Scholar] [CrossRef] [Green Version]
- Nicholson, R.L.; Hammerschmidt, R. Phenolic compounds and their role in disease resistance. Ann. Rev. Phytopathol. 1992, 30, 369–389. [Google Scholar] [CrossRef]
- Tan, Y.; Li, Q.; Zhao, Y.; Wei, H.; Wang, J.; Baker, C.J.; Liu, Q.; Wei, W. Integration of metabolomics and existing omics data reveals new insights into phytoplasma-induced metabolic reprogramming in host plants. PLoS ONE 2021, 16, e0246203. [Google Scholar] [CrossRef]
- De Oliveira, E.; Magalhães, P.C.; Gomide, R.L.; Vasconcelos, C.A.; Souza, I.R.P.; Oliveira, C.M.; Cruz, I.; Schaffert, R.E. Growth and nutrition of mollicute-infected maize. Plant Dis. 2002, 86, 945–949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broadley, M.; Brown, P.; Çakmak, I.; Rengel, Z.; Zhao, F. Function of nutrients: Micronutrients. In Marschner’s Mineral Nutrition of Higher Plants; Marschner, P., Ed.; Academic Press: London, UK, 2012; pp. 191–248. [Google Scholar] [CrossRef]
- Buoso, S.; Pagliari, L.; Musetti, R.; Martini, M.; Marroni, F.; Schmidt, W.; Santi, S. Candidatus Phytoplasma solani’ interferes with the distribution and uptake of iron in tomato. BMC Genom. 2019, 20, 1–21. [Google Scholar] [CrossRef]
- Bertamini, M.; Nedunchezhian, N.; Tomasi, F.; Grando, S. Phytoplasma [Stolbur subgroup (Bois Noir-BN)] infection inhibits photosynthetic pigments, ribulose-1, 5-biphosphate carboxylase and photosynthetic activities in field grown grapevine (Vitis vinifera L. cv. Chardonnay) leaves. Physiol. Mol. Plant Pathol. 2002, 61, 357–366. [Google Scholar] [CrossRef]
- Wu, Y.; Jin, X.; Liao, W.; Hu, L.; Dawuda, M.M.; Zhao, X.; Tang, Z.; Gong, T.; Yu, J. 5-Aminolevulinic acid (ALA) alleviated salinity stress in cucumber seedlings by enhancing chlorophyll synthesis pathway. Front. Plant Sci. 2018, 9, 635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, X.; Gai, Y.; Zheng, C.; Mu, Z. Comparative proteomic analysis provides new insights into mulberry dwarf responses in mulberry (Morus alba L.). Proteomics 2009, 9, 5328–5339. [Google Scholar] [CrossRef]
- Ding, Y.; Wei, W.; Wu, W.; Davis, R.E.; Jiang, Y.; Lee, I.M.; Hammond, R.W.; Shen, L.; Sheng, J.P.; Zhao, Y. Role of gibberellic acid in tomato defense against potato purple top phytoplasma infection. Ann. Appl. Biol. 2013, 162, 191–199. [Google Scholar] [CrossRef]
- Fan, G.; Dong, Y.; Deng, M.; Zhao, Z.; Niu, S.; Xu, E. Plant-pathogen interaction, circadian rhythm, and hormone-related gene expression provide indicators of phytoplasma infection in Paulownia fortunei. Int. J. Mol. Sci. 2014, 15, 23141–23162. [Google Scholar] [CrossRef]
- Ustun, R.; Yol, E.; Ikten, C.; Catal, M.; Uzun, B. Screening, selection and real-time qPCR validation for phytoplasma resistance in sesame (Sesamum indicum L.). Euphytica 2017, 213, 1–9. [Google Scholar] [CrossRef]
- Li, C.; Miao, H.; Wei, L.; Zhang, T.; Han, X.; Zhang, H. Association mapping of seed oil and protein content in Sesamum indicum L. using SSR markers. PLoS ONE 2014, 9, e105757. [Google Scholar] [CrossRef] [Green Version]
- Bellaloui, N.; Abbas, H.K.; Ebelhar, M.W.; Mengistu, A.; Mulvaney, M.J.; Accinelli, C.; Shier, W.T. Effect of increased nitrogen application rates and environment on protein, oil, fatty acids, and minerals in sesame (Sesamum indicum) seed grown under Mississippi Delta conditions. Food Nutr. Sci. 2018, 9, 1115. [Google Scholar] [CrossRef] [Green Version]
- Hossain, Z.; Johnson, E.N.; Wang, L.; Blackshaw, R.E.; Gan, Y. Comparative analysis of oil and protein content and seed yield of five Brassicaceae oilseeds on the Canadian prairie. Ind. Crops Prod. 2019, 136, 77–86. [Google Scholar] [CrossRef]
- Kambhampati, S.; Aznar-Moreno, J.A.; Hostetler, C.; Caso, T.; Bailey, S.R.; Hubbard, A.H.; Durrett, T.P.; Allen, D.K. On the inverse correlation of protein and oil: Examining the effects of altered central carbon metabolism on seed composition using soybean fast neutron mutants. Metabolites 2020, 10, 18. [Google Scholar] [CrossRef] [Green Version]
- Xue, C.; Liu, Z.; Wang, L.; Li, H.; Gao, W.; Liu, M.; Zhao, Z.; Zhao, J. The antioxidant defense system in Chinese jujube is triggered to cope with phytoplasma invasion. Tree Physiol. 2020, 40, 1437–1449. [Google Scholar] [CrossRef] [PubMed]
- Aslam, F.; Iqbal, S.; Nasir, M.; Anjum, A.A. White sesame seed oil mitigates blood glucose level, reduces oxidative stress, and improves biomarkers of hepatic and renal function in participants with type 2 diabetes mellitus. J. Am. Coll. Nutr. 2019, 38, 235–246. [Google Scholar] [CrossRef]
- Gholinezhad, E.; Darvishzadeh, R. Influence of arbuscular mycorrhiza fungi and drought stress on fatty acids profile of sesame (Sesamum indicum L.). Field Crops Res. 2021, 262, 108035. [Google Scholar] [CrossRef]
- Nazari, M.; Mirlohi, A.; Majidi, M.M. Effects of drought stress on oil characteristics of Carthamus Species. J. Am. Oil Chem. Soc. 2017, 94, 247–256. [Google Scholar] [CrossRef]
- Ponce, V.; Pandey, R.; Ercan, S. Characterization of drought across climatic spectrum. J. Hydrol. Eng. 2000, 5, 222–224. [Google Scholar] [CrossRef]
- Soil Survey Staff USDA. Soil Taxonomy. A Basic System of Soil Classification for Making Sand Interpreting Soil Surveys, 2nd ed.; Handbook 436; Natural Resources Conservation Service, U.S. Department of Agriculture: Washington, DC, USA, 1999.
- Dellaporta, S.L.; Wood, J.; Hicks, J.B. A plant DNA minipreparation: Version II. Plant Mol. Biol. Rep. 1983, 1, 19–21. [Google Scholar] [CrossRef]
- Casati, P.; Quaglino, F.; Tedeschi, R.; Spiga, F.M.; Alma, A.; Spadone, P.; Bianco, P.A. Identification and molecular characterisation of ‘Candidatus Phytoplasma mali’ isolates in North-Western Italy. J. Phytopathol. 2010, 158, 81–87. [Google Scholar] [CrossRef]
- Franova, J.; Ludvíková, H.; Paprstein, F. Detection and characterization of phytoplasmas infecting apple trees in Czech Republic during 2010. J. Bull. Insectol. 2011, 64, 51–52. [Google Scholar]
- Nassar, M.; El-Sahhar, K. Botanical Preparations and Microscopy (Microtechnique); Academic Bookshop: Giza, Egypt, 1998; p. 219. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Jia, Z.S.; Tang, M.C.; Wu, J.M. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Saxena, M.; Saxena, J.; Nema, R.; Singh, D.; Gupta, A. Phytochemistry of medicinal plants. J. Pharmacogn. Phytochem. 2013, 1, 168–182. [Google Scholar]
- Nagpurkar, M.; Patil, N.M. Qualitative and Quantitative Phytochemical Studies in Different Parts of Sesamum indicum L. In Medicinal Plants: Biodiversity, Sustainable Utilization and Conservation; Springer: Singapore, 2020; pp. 583–595. [Google Scholar]
- Jones, M.M.; Turner, N.C. Osmotic adjustment in leaves of sorghum in response to water deficits. Plant Physiol. 1978, 61, 122–126. [Google Scholar] [CrossRef] [Green Version]
- Sinclair, T.R. Historical changes in harvest index and crop nitrogen accumulation. Crop Sci. 1998, 38, 638–643. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of International, Method; Association of Official Analytical Chemists: Washington, DC, USA, 1995. [Google Scholar]
- AOCS. American Oil Chemist’s Society, Method Ag 1–65; Official Methods and Recommended Practices of the American Oil Chemist’s Society; AOCS: Washington, DC, USA, 1993.
- AOAC. Official Methods of Analysis of International; Association of Official Analytical Chemists: Washington, DC, USA, 2005. [Google Scholar]
- Joslyn, M.A. Methods in Food Analysis Applied to Plant Products; Academic Press: New York, NY, USA, 1950. [Google Scholar]
- AOCS. American Oil Chemist’s Society, Method Ca 5a-40; Official Methods and Recommended Practices of the American Oil Chemist’s Society; AOCS: Washington, DC, USA, 1998.
- AOCS. American Oil Chemist’s Society, Method Cd 3d-63; Official Methods and Recommended Practices of the American Oil Chemist’s Society; AOCS: Washington, DC, USA, 1999.
- AOCS. American Oil Chemist’s Society, Method Cd 8b-90; Official Methods and Recommended Practices of the American Oil Chemist’s Society; AOCS: Washington, DC, USA, 2011.
- Levene, H. Robust tests of equality of variances. In Contributions to Probability and Statistics, Essays in Honor of Harold Hoteling; Olkin, I., Ghurye, S.G., Hoeffding, W., Madow, W.G., Mann, H.B., Eds.; Stanford University Press: Stanford, CA, USA, 1960; pp. 278–292. [Google Scholar]
- Razali, N.; Wah, Y.B. Power comparisons of Shapiro-Wilk, Kolmogorov-Smirnov, Lilliefors and Anderson-Darling tests. J. Stat. Modeling Anal. 2011, 2, 21–33. [Google Scholar]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters (μm) | Healthy Sesame | Infected Sesame | Change (%) ∆ |
|---|---|---|---|
| Upper epidermis thickness | 13.74 | 10.42 | −24.2 |
| Lower epidermis thickness | 14.59 | 9.82 | −32.7 |
| Midvein width | 757.13 | 864.00 | +14.1 |
| Midvein length | 569.86 | 707.56 | +24.2 |
| Blade thickness | 256.84 | 225.39 | −12.2 |
| Palisade tissue thickness | 103.02 | 77.26 | −25.0 |
| Spongy tissue thickness | 135.65 | 121.64 | −10.3 |
| Vascular bundle width | 516.70 | 549.22 | +6.3 |
| Vascular bundle length | 135.04 | 177.35 | +31.3 |
| Protoxylem vessels height | 14.06 | 18.94 | +34.7 |
| Protoxylem vessels width | 18.48 | 21.26 | +15.1 |
| Metaxylem vessels height | 24.21 | 23.59 | −2.5 |
| Metaxylem vessels width | 27.43 | 26.20 | −4.5 |
| Parameters (μm) | Healthy Sesame | Infected Sesame | Change (%) ∆ |
|---|---|---|---|
| Stem diameter | 2788.56 | 3876.16 | +39.0 |
| Cortex thickness | 217.34 | 283.38 | +30.4 |
| Number of cortex layers | 10.00 | 12.00 | +20.0 |
| Vascular cylinder thickness | 2260.59 | 3190.25 | +41.1 |
| Vascular tissues thickness | 339.71 | 1205.23 | +254.8 |
| Phloem zone thickness | 79.33 | 265.09 | +234.1 |
| Xylem zone thickness | 228.87 | 1087.74 | +375.3 |
| Pith diameter | 1487.40 | 1737.42 | +16.8 |
| Parameter | Healthy Plants | Infected Plants | Change (%) ∆ | p-Value |
|---|---|---|---|---|
| Total soluble proteins (mg g−1 DW) | 2.24 ± 0.04 b | 2.90 ± 0.12 a | +29.5 | 0.005 ** |
| Total phenolic content (mg GAEs g−1 DW) | 5.02 ± 0.04 b | 13.35 ± 0.09 a | +165.9 | <0.001 ** |
| Total flavonoid content (mg QUEs g−1 DW) | 0.99 ± 0.01 b | 1.63 ± 0.05 a | +64.7 | <0.001 ** |
| Total tannins (mg TAEs g−1 DW) | 0.24 ± 0.01 b | 0.28 ± 0.01 a | +16.7 | 0.036 * |
| Total alkaloids (mg g−1 DW) | 1.04 ± 0.02 b | 5.84 ± 0.04 a | +461.5 | <0.001 ** |
| Parameter | Healthy Plants | Infected Plants | Change (%) ∆ | p-Value |
|---|---|---|---|---|
| Main stem height (cm) | 180.33 ± 6.1 a | 104.33 ± 2.3 b | −42.1 | <0.001 ** |
| 1st-order branches number plant−1 | 4.50 ± 0.5 b | 6.58 ± 0.2 a | +46.2 | 0.014 * |
| Leaves number plant−1 | 109.00 ± 2.6 a | 70.43 ± 2.3 b | −35.4 | <0.001 ** |
| Main stem diameter (cm) | 1.63 ± 0.03 a | 1.29 ± 0.02 b | −20.9 | <0.001 ** |
| Leaf area per plant (dm2) | 67.92 ± 1.6 a | 24.09 ± 0.1 b | −64.5 | 0.001 ** |
| Plant dry weight (g) | 114.21 ± 2.0 a | 66.63 ± 1.8 b | −41.7 | <0.001 ** |
| Leaf greenness index | 48.51 ± 0.9 a | 26.17 ± 0.7 b | −46.1 | <0.001 ** |
| Relative water content (%) | 77.43 ± 1.2 a | 68.60 ± 1.0 b | −11.4 | 0.004 ** |
| Parameter | Healthy Plants | Infected Plants | Change (%) ∆ | p-Value |
|---|---|---|---|---|
| Seed yield plant−1 (g) | 17.61 ± 0.35 a | 10.91 ± 0.29 b | −38.0 | <0.001 ** |
| Seed index (g) | 3.61 ± 0.05 a | 3.25 ± 0.04 b | −10.0 | 0.005 ** |
| Seed yield (kg ha−1) | 1467.0 ± 26.5 a | 910.4 ± 24.0 b | −37.9 | <0.001 ** |
| Oil yield (kg ha−1) | 756.2 ± 16.9 a | 435.2 ± 10.9 b | −42.5 | <0.001 ** |
| Biological yield (t ha−1) | 6.18 ± 0.37 a | 7.30 ± 0.65 a | +18.1 | 0.206 ns |
| Seed harvest index | 23.95 ± 0.20 a | 12.65 ± 0.78 b | −47.2 | <0.001 ** |
| Seed protein content (%) | 19.32 ± 0.10 b | 21.27 ± 0.24 a | +10.1 | 0.002 ** |
| Seed oil content (%) | 51.54 ± 0.28 a | 47.81 ± 0.40 b | −7.2 | 0.002 ** |
| Parameter | Healthy Plants | Infected Plants | Change (%) ∆ | p-Value |
|---|---|---|---|---|
| Refractive index | 1.474 ± 0.00 a | 1.474 ± 0.00 a | 0.00 | >0.9999 ns |
| Flow rate | 5.43 ± 0.02 a | 5.47 ± 0.06 a | +0.74 | 0.500 ns |
| Acid value (mg KOH g−1) | 3.18 ± 0.21 a | 3.28 ± 0.61 a | +3.14 | 0.887 ns |
| Acidity (%) | 1.60 ± 0.10 a | 1.65 ± 0.31 a | +3.12 | 0.887 ns |
| Peroxide value (meq O2 kg−1 oil) | 9.57 ± 0.53 b | 13.70 ± 0.63 a | +43.2 | 0.007 ** |
| Month | Temperature (°C) | Relative Humidity (%) | Wind Speed (m s−1) | Precipitation (mm) | Solar Radiation(MJ m−2 d−1) | |
|---|---|---|---|---|---|---|
| Day | Night | |||||
| May | 34.2 | 17.2 | 36.1 | 3.73 | 0.00 | 28.7 |
| June | 36.9 | 19.9 | 31.9 | 3.70 | 0.30 | 31.6 |
| July | 38.2 | 22.0 | 34.4 | 3.68 | 0.00 | 32.7 |
| August | 38.7 | 22.6 | 36.2 | 3.42 | 0.00 | 30.7 |
| September | 38.1 | 22.4 | 40.3 | 3.74 | 0.00 | 28.3 |
| Primer | Sequence | Size of the PCR Product |
|---|---|---|
| P1 | AAGAGTTTGATCCTGGCTCAGGATT | 1.8 kb |
| P7 | CGTCCTTCATCGGCTCTT | |
| R16F2n | GAAACGACTGCTAAGACTGG | 1.2 kb |
| R16R2 | TGACGGGCGGTGTGTACAAACCCCG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmed, E.A.; Farrag, A.A.; Kheder, A.A.; Shaaban, A. Effect of Phytoplasma Associated with Sesame Phyllody on Ultrastructural Modification, Physio-Biochemical Traits, Productivity and Oil Quality. Plants 2022, 11, 477. https://doi.org/10.3390/plants11040477
Ahmed EA, Farrag AA, Kheder AA, Shaaban A. Effect of Phytoplasma Associated with Sesame Phyllody on Ultrastructural Modification, Physio-Biochemical Traits, Productivity and Oil Quality. Plants. 2022; 11(4):477. https://doi.org/10.3390/plants11040477
Chicago/Turabian StyleAhmed, Eman A., Amro A. Farrag, Ahmed A. Kheder, and Ahmed Shaaban. 2022. "Effect of Phytoplasma Associated with Sesame Phyllody on Ultrastructural Modification, Physio-Biochemical Traits, Productivity and Oil Quality" Plants 11, no. 4: 477. https://doi.org/10.3390/plants11040477