
Plant GDSL Esterases/Lipases: Evolutionary, Physiological and Molecular Functions in Plant Development


Abstract
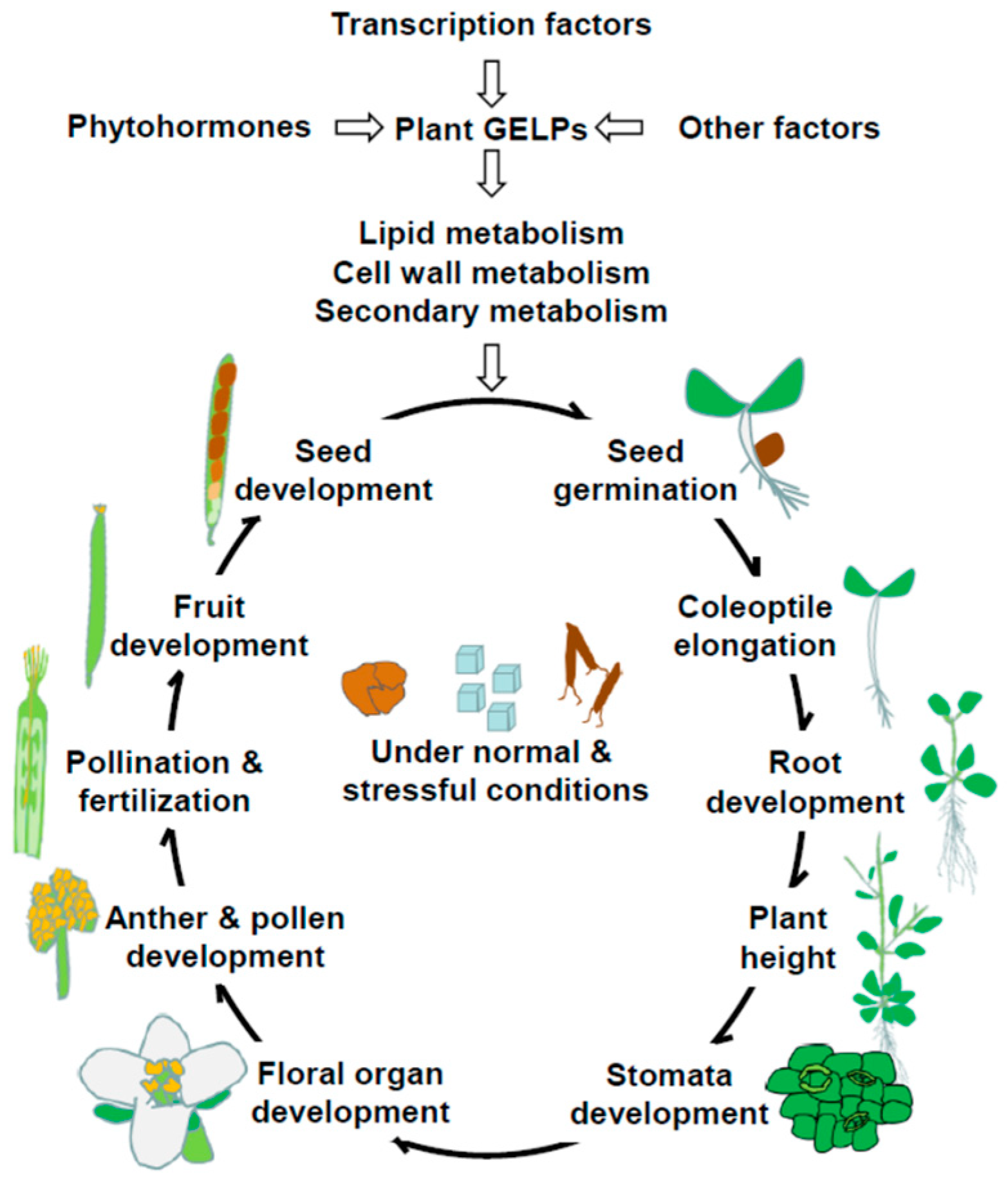
:1. Introduction
2. Roles of GELP in Plant Land Colonization
3. Roles of GELPs in Plant Development and Plant Metabolism
3.1. Vegetative Development
3.1.1. Seed Germination
3.1.2. Coleoptile Elongation
3.1.3. Lateral Root Development
3.1.4. Root Development
3.1.5. Plant Height
3.1.6. Stomata Development
3.2. Reproductive Development
3.2.1. Flower Development
3.2.2. Anther and Pollen Development
3.2.3. Pollen-Stigma Interaction and Pollinator Attraction
3.2.4. Fruit Development
3.2.5. Seed Development
3.3. Plant Metabolism
3.3.1. Lipid Metabolism
3.3.2. Cell Wall Metabolism
3.3.3. Secondary Metabolism
4. Roles of GELPs in Plant-Environment Interactions
4.1. Plant-Abiotic Stress Interaction
4.2. Plant-Biotic Stress Interaction
5. Possible Regulatory Factors and Networks
6. Concluding Remarks and Future Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ding, L.N.; Wang, W.J.; Cao, J.; Wang, Z.; Zhu, K.M.; Yang, Y.H.; Li, Y.L.; Tan, X.L. Advances in plant GDSL lipases: From sequences to functional mechanisms. Acta Physiol. Plant. 2019, 41, 151. [Google Scholar] [CrossRef]
- Volokita, M.; Rosilio-Brami, T.; Rivkin, N.; Zik, M. Combining comparative sequence and genomic data to ascertain phylogenetic relationships and explore the evolution of the large GDSL-lipase family in land plants. Mol. Biol. Evol. 2011, 28, 551–565. [Google Scholar] [CrossRef] [Green Version]
- Vujaklija, I.; Bielen, A.; Paradzik, T.; Bidin, S.; Goldstein, P.; Vujaklija, D. An effective approach for annotation of protein families with low sequence similarity and conserved motifs: Identifying GDSL hydrolases across the plant kingdom. BMC Bioinform. 2016, 17, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, C.P.; Huang, L.M.; Chen, L.O.; Chan, M.T.; Shaw, J.F. Genome-wide analysis of GDSL-type esterases/lipases in Arabidopsis. Plant Mol. Biol. 2017, 95, 181–197. [Google Scholar] [CrossRef]
- Chepyshko, H.; Lai, C.P.; Huang, L.M.; Liu, J.H.; Shaw, J.F. Multifunctionality and diversity of GDSL esterase/lipase gene family in rice (Oryza sativa L. japonica) genome: New insights from bioinformatics analysis. BMC Genom. 2012, 13, 309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, X.; Yi, H.; Han, C.T.; Nou, I.S.; Hur, Y. GDSL esterase/lipase genes in Brassica rapa L.: Genome-wide identification and expression analysis. Mol. Genet. Genom. 2016, 291, 531–542. [Google Scholar] [CrossRef]
- Ni, P.Y.; Ji, X.R.; Guo, D.L. Genome-wide identification, characterization, and expression analysis of GDSL-type esterases/lipases gene family in relation to grape berry ripening. Sci. Hortic. 2020, 264, 109162. [Google Scholar] [CrossRef]
- An, X.L.; Dong, Z.Y.; Tian, Y.H.; Xie, K.; Wu, S.W.; Zhu, T.T.; Zhang, D.F.; Zhou, Y.; Niu, C.F.; Ma, B.; et al. ZmMs30 encoding a novel GDSL lipase is essential for male fertility and valuable for hybrid breeding in maize. Mol. Plant 2019, 12, 343–359. [Google Scholar] [CrossRef] [Green Version]
- Su, H.G.; Zhang, X.H.; Wang, T.T.; Wei, W.L.; Wang, Y.X.; Chen, J.; Zhou, Y.B.; Chen, M.; Ma, Y.Z.; Xu, Z.S.; et al. Genome-wide identification, evolution, and expression of GDSL-type esterase/lipase gene family in soybean. Front. Plant Sci. 2020, 11, 726. [Google Scholar] [CrossRef]
- Cao, Y.; Han, Y.; Meng, D.; Abdullah, M.; Yu, J.; Li, D.; Jin, Q.; Lin, Y.; Cai, Y. Expansion and evolutionary patterns of GDSL-type esterases/lipases in Rosaceae genomes. Funct. Integr. Genom. 2018, 18, 673–684. [Google Scholar] [CrossRef]
- Niklas, K.J.; Cobb, E.D.; Matas, A.J. The evolution of hydrophobic cell wall biopolymers: From algae to angiosperms. J. Exp. Bot. 2017, 68, 5261–5269. [Google Scholar] [CrossRef] [Green Version]
- Philippe, G.; Sorensen, I.; Jiao, C.; Sun, X.; Fei, Z.; Domozych, D.S.; Rose, J.K. Cutin and suberin: Assembly and origins of specialized lipidic cell wall scaffolds. Curr. Opin. Plant Biol. 2020, 55, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Smyth, D.R. Wrinkles on sepals: Cuticular ridges form when cuticle production outpaces epidermal cell expansion. Mol. Plant 2017, 10, 540–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeats, T.H.; Huang, W.; Chatterjee, S.; Viart, H.M.; Clausen, M.H.; Stark, R.E.; Rose, J.K. Tomato Cutin Deficient 1 (CD1) and putative orthologs comprise an ancient family of cutin synthase-like (CUS) proteins that are conserved among land plants. Plant J. 2014, 77, 667–675. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Liu, Y.; Zhi, P.; Wang, X.; Xu, B.; Gong, Z.; Chang, C. Origins and evolution of cuticle biosynthetic machinery in land plants. Plant Physiol. 2020, 184, 1998–2010. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.Y.; Chen, R.J.; Dong, J.L.; Xu, Z.J.; Gao, X.L. Analysis of GDSL lipase (GLIP) family genes in rice (Oryza sativa). Plant Omics 2012, 5, 351–358. [Google Scholar]
- Dolui, A.K.; Vijayaraj, P. Functional omics identifies serine hydrolases that mobilize storage lipids during rice seed germination. Plant Physiol. 2020, 184, 693–708. [Google Scholar] [CrossRef] [PubMed]
- Clauss, K.; von Roepenack-Lahaye, E.; Bottcher, C.; Roth, M.R.; Welti, R.; Erban, A.; Kopka, J.; Scheel, D.; Milkowski, C.; Strack, D. Overexpression of sinapine esterase BnSCE3 in oilseed rape seeds triggers global changes in seed metabolism. Plant Physiol. 2011, 155, 1127–1145. [Google Scholar] [CrossRef] [Green Version]
- Rombola-Caldentey, B.; Rueda-Romero, P.; Iglesias-Fernandez, R.; Carbonero, P.; Onate-Sanchez, L. Arabidopsis DELLA and two HD-ZIP transcription factors regulate GA signaling in the epidermis through the L1 Box cis-element. Plant Cell 2014, 26, 2905–2919. [Google Scholar] [CrossRef] [Green Version]
- Riemann, M.; Gutjahr, C.; Korte, A.; Riemann, M.; Danger, B.; Muramatsu, T.; Bayer, U.; Waller, F.; Furuya, M.; Nick, P. GER1, a GDSL motif-encoding gene from rice is a novel early light- and jasmonate-induced gene. Plant Biol. 2007, 9, 32–40. [Google Scholar] [CrossRef] [Green Version]
- Ursache, R.; De Jesus Vieira Teixeira, C.; Denervaud Tendon, V.; Gully, K.; De Bellis, D.; Schmid-Siegert, E.; Grube Andersen, T.; Shekhar, V.; Calderon, S.; Pradervand, S.; et al. GDSL-domain proteins have key roles in suberin polymerization and degradation. Nat. Plants 2021, 7, 353–364. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.S.; Kim, B.K.; Kwon, S.J.; Jin, H.C.; Park, O.K. Arabidopsis GDSL lipase 2 plays a role in pathogen defense via negative regulation of auxin signaling. Biochem. Biophys. Res. Commun. 2009, 379, 1038–1042. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Shimada, T.; Kondo, M.; Tamai, A.; Mori, M.; Nishimura, M.; Hara-Nishimura, I. Ectopic expression of an esterase, which is a candidate for the unidentified plant cutinase, causes cuticular defects in Arabidopsis thaliana. Plant Cell Physiol. 2010, 51, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Ma, B.; Duan, K.X.; Li, X.K.; Lu, X.; Yin, C.C.; Tao, J.J.; Wei, W.; Zhang, W.K.; Xin, P.Y.; et al. The GDSL lipase MHZ11 modulates ethylene signaling in rice roots. Plant Cell 2020, 32, 1626–1643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savaldi-Goldstein, S.; Peto, C.; Chory, J. The epidermis both drives and restricts plant shoot growth. Nature 2007, 446, 199–202. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Woo, M.O.; Rihua, P.; Koh, H.J. The DROOPING LEAF (DR) gene encoding GDSL esterase is involved in silica deposition in rice (Oryza sativa L.). PLoS ONE 2020, 15, e0238887. [Google Scholar] [CrossRef] [PubMed]
- Park, J.J.; Jin, P.; Yoon, J.; Yang, J.I.; Jeong, H.J.; Ranathunge, K.; Schreiber, L.; Franke, R.; Lee, I.J.; An, G. Mutation in Wilted Dwarf and Lethal 1 (WDL1) causes abnormal cuticle formation and rapid water loss in rice. Plant Mol. Biol. 2010, 74, 91–103. [Google Scholar] [CrossRef]
- Tang, J.; Yang, X.P.; Xiao, C.L.; Li, J.Y.; Chen, Y.Q.; Li, R.Y.; Li, S.P.; Lu, S.Y.; Hu, H.H. GDSL lipase occluded stomatal pore 1 is required for wax biosynthesis and stomatal cuticular ledge formation. New Phytol. 2020, 228, 1880–1896. [Google Scholar] [CrossRef]
- Ishiguro, S.; Kawai-Oda, A.; Ueda, J.; Nishida, I.; Okada, K. The defective in anther dehiscence1 gene encodes a novel phospholipase A1 catalyzing the initial step of jasmonic acid biosynthesis, which synchronizes pollen maturation, anther dehiscence, and flower opening in Arabidopsis. Plant Cell 2001, 13, 2191–2209. [Google Scholar] [CrossRef] [Green Version]
- Philippe, G.; Gaillard, C.; Petit, J.; Geneix, N.; Dalgalarrondo, M.; Bres, C.; Mauxion, J.P.; Franke, R.; Rothan, C.; Schreiber, L.; et al. Ester cross-link profiling of the cutin polymer of wild-type and cutin synthase tomato mutants highlights different mechanisms of polymerization. Plant Physiol. 2016, 170, 807–820. [Google Scholar] [CrossRef] [Green Version]
- Hong, L.L.; Brown, J.; Segerson, N.A.; Rose, J.K.C.; Roeder, A.H.K. CUTIN SYNTHASE 2 maintains progressively Developing cuticular ridges in Arabidopsis sepals. Mol. Plant 2017, 10, 560–574. [Google Scholar] [CrossRef] [Green Version]
- Ran, X.R.; Chen, X.; Shi, L.; Ashraf, M.; Yan, F.Z.; Chen, Y.Q.; Xu, J.; Shi, J.X. Transcriptomic insights into the roles of HSP70-16 in sepal’s responses to developmental and mild heat stress signals. Environ. Exp. Bot. 2020, 179, 104225. [Google Scholar] [CrossRef]
- Chen, X.; Shi, L.; Chen, Y.Q.; Zhu, L.; Zhang, D.S.; Xiao, S.; Aharoni, A.; Shi, J.X.; Xu, J. Arabidopsis HSP70-16 is required for flower opening under normal or mild heat stress temperatures. Plant Cell Environ. 2019, 42, 1190–1204. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Shi, J.; Yang, X. Role of lipid metabolism in plant pollen exine development. In Lipids in Plant and Algae Development; Nakamura, Y., Li-Beisson, Y., Eds.; Springer: Cham, Switzerland, 2016; Volume 86, pp. 315–337. ISBN 978-3-319-25979-6. [Google Scholar]
- Shi, J.; Cui, M.; Yang, L.; Kim, Y.J.; Zhang, D. Genetic and biochemical mechanisms of pollen wall development. Trends Plant Sci. 2015, 20, 741–753. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Lou, Y.; Shi, Q.S.; Zhang, S.; Zhou, W.T.; Yang, J.; Zhang, C.; Yao, X.Z.; Xu, T.; Liu, J.L.; et al. Slowing development restores the fertility of thermo-sensitive male-sterile plant lines. Nat. Plants 2020, 6, 360–367. [Google Scholar] [CrossRef] [PubMed]
- Tsugama, D.; Fujino, K.; Liu, S.; Takano, T. A GDSL-type esterase/lipase gene, GELP77, is necessary for pollen dissociation and fertility in Arabidopsis. Biochem. Biophys. Res. Commun. 2020, 526, 1036–1041. [Google Scholar] [CrossRef]
- Xu, J.; Ding, Z.W.; Vizcay-Barrena, G.; Shi, J.X.; Liang, W.Q.; Yuan, Z.; Werck-Reichhart, D.; Schreiber, L.; Wilson, Z.A.; Zhang, D.B. ABORTED MICROSPORES acts as a master regulator of pollen wall formation in Arabidopsis. Plant Cell 2014, 26, 1544–1556. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Long, T.; Wang, Y.F.; Tong, X.H.; Tang, J.; Li, J.L.; Wang, H.M.; Tang, L.Q.; Li, Z.Y.; Shu, Y.Z.; et al. RMS2 encoding a GDSL lipase mediates lipid homeostasis in anthers to determine rice male fertility. Plant Physiol. 2020, 182, 2047–2064. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.H.; Wang, M.L.; Li, Y.Q.; Yan, W.; Chang, Z.Y.; Ni, H.L.; Chen, Z.F.; Wu, J.X.; Xu, C.J.; Deng, X.W.; et al. GDSL esterase/lipases OsGELP34 and OsGELP110/OsGELP115 are essential for rice pollen development. J. Integr. Plant Biol. 2020, 62, 1574–1593. [Google Scholar] [CrossRef] [Green Version]
- Yuan, G.Q.; Zou, T.; Zhang, X.; Liu, M.M.; Luo, T.T.; He, Z.Y.; Tao, Y.; Zhou, D.; Zhao, H.F.; Liu, S.J.; et al. A rice GDSL esterase/lipase protein (GELP) is required for anther and pollen development. Mol. Breed. 2020, 40, 90. [Google Scholar] [CrossRef]
- Huo, Y.Q.; Pei, Y.R.; Tian, Y.H.; Zhang, Z.G.; Li, K.; Liu, J.; Xiao, S.L.; Chen, H.B.; Liu, J. IRREGULAR POLLEN EXINE2 encodes a GDSL lipase essential for male fertility in maize. Plant Physiol. 2020, 184, 1438–1454. [Google Scholar] [CrossRef] [PubMed]
- Updegraff, E.P.; Zhao, F.; Preuss, D. The extracellular lipase EXL4 is required for efficient hydration of Arabidopsis pollen. Sex. Plant Reprod. 2009, 22, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Kram, B.W.; Bainbridge, E.A.; Perera, M.A.D.N.; Carter, C. Identification, cloning and characterization of a GDSL lipase secreted into the nectar of Jacaranda mimosifolia. Plant Mol. Biol. 2008, 68, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Tieman, D.; Liu, Z.M.; Chen, K.S.; Klee, H.J. Identification of a lipase gene with a role in tomato fruit short-chain fatty acid-derived flavor volatiles by genome-wide association. Plant J. 2020, 104, 631–644. [Google Scholar] [CrossRef]
- Girard, A.L.; Mounet, F.; Lemaire-Chamley, M.; Gaillard, C.; Elmorjani, K.; Vivancos, J.; Runavot, J.L.; Quemener, B.; Petit, J.; Germain, V.; et al. Tomato GDSL1 is required for cutin deposition in the fruit cuticle. Plant Cell 2012, 24, 3119–3134. [Google Scholar] [CrossRef] [Green Version]
- Petit, J.; Bres, C.; Just, D.; Garcia, V.; Mauxion, J.P.; Marion, D.; Bakan, B.; Joubes, J.; Domergue, F.; Rothan, C. Analyses of tomatoFfruit brightness mutants uncover both cutin-deficient and cutin-abundant mutants and a new hypomorphic allele of GDSL lipase. Plant Physiol. 2014, 164, 888–906. [Google Scholar] [CrossRef] [Green Version]
- Ma, R.D.; Yuan, H.L.; An, J.; Hao, X.Y.; Li, H.B. A Gossypium hirsutum GDSL lipase/hydrolase gene (GhGLIP) appears to be involved in promoting seed growth in Arabidopsis. PLoS ONE 2018, 13, e0195556. [Google Scholar] [CrossRef] [Green Version]
- Kondou, Y.; Nakazawa, M.; Kawashima, M.; Ichikawa, T.; Yoshizumi, T.; Suzuki, K.; Ishikawa, A.; Koshi, T.; Matsui, R.; Muto, S.; et al. RETARDED GROWTH OF EMBRYO1, a new basic helix-loop-helix protein, expresses in endosperm to control embryo growth. Plant Physiol. 2008, 147, 1924–1935. [Google Scholar] [CrossRef] [Green Version]
- Hong, J.K.; Choi, H.W.; Hwang, I.S.; Kim, D.S.; Kim, N.H.; Choi, D.S.; Kim, Y.J.; Hwang, B.K. Function of a novel GDSL-type pepper lipase gene, CaGLIP1, in disease susceptibility and abiotic stress tolerance. Planta 2008, 227, 539–558. [Google Scholar] [CrossRef]
- Huang, L.M.; Lai, C.P.; Chen, L.F.O.; Chan, M.T.; Shaw, J.F. Arabidopsis SFAR4 is a novel GDSL-type esterase involved in fatty acid degradation and glucose tolerance. Bot. Stud. 2015, 56, 33. [Google Scholar] [CrossRef] [Green Version]
- Gao, M.J.; Yi, X.; Yang, W.B.; Lam, S.M.; Tong, X.H.; Liu, J.Y.; Wang, X.; Li, Q.; Shui, G.H.; He, Z.H. GDSL lipases modulate immunity through lipid homeostasis in rice. PLoS Pathog. 2017, 13, e1006724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.; Froehlich, J.E.; Zienkiewicz, A.; Hersh, H.L.; Benning, C. A Plastid phosphatidylglycerol lipase contributes to the export of acyl groups from plastids for seed oil biosynthesis. Plant Cell 2017, 29, 1678–1696. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.J.; Gao, C.X.; Mentink-Vigier, F.; Tang, L.; Zhang, D.M.; Wang, S.G.; Cao, S.X.; Xu, Z.P.; Liu, X.L.; Wang, T.; et al. Arabinosyl deacetylase modulates the arabinoxylan acetylation profile and secondary wall formation. Plant Cell 2019, 31, 1113–1126. [Google Scholar] [CrossRef]
- Zhang, B.; Zhang, L.; Li, F.; Zhang, D.; Liu, X.; Wang, H.; Xu, Z.; Chu, C.; Zhou, Y. Control of secondary cell wall patterning involves xylan deacetylation by a GDSL esterase. Nat. Plants 2017, 3, 17017. [Google Scholar] [CrossRef] [PubMed]
- Yadav, V.K.; Yadav, V.K.; Pant, P.; Singh, S.P.; Maurya, R.; Sable, A.; Sawant, S.V. GhMYB1 regulates SCW stage-specific expression of the GhGDSL promoter in the fibres of Gossypium hirsutum L. Plant Biotechnol. J. 2017, 15, 1163–1174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeats, T.H.; Martin, L.B.; Viart, H.M.; Isaacson, T.; He, Y.; Zhao, L.; Matas, A.J.; Buda, G.J.; Domozych, D.S.; Clausen, M.H.; et al. The identification of cutin synthase: Formation of the plant polyester cutin. Nat. Chem. Biol. 2012, 8, 609–611. [Google Scholar] [CrossRef] [Green Version]
- Kikuta, Y.; Ueda, H.; Takahashi, M.; Mitsumori, T.; Yamada, G.; Sakamori, K.; Takeda, K.; Furutani, S.; Nakayama, K.; Katsuda, Y.; et al. Identification and characterization of a GDSL lipase-like protein that catalyzes the ester-forming reaction for pyrethrin biosynthesis in Tanacetum cinerariifolium- a new target for plant protection. Plant J. 2012, 71, 183–193. [Google Scholar] [CrossRef]
- Teutschbein, J.; Gross, W.; Nimtz, M.; Milkowski, C.; Hause, B.; Strack, D. Identification and localization of a lipase-like acyltransferase in phenylpropanoid metabolism of tomato (Solanum lycopersicum). J. Biol. Chem. 2010, 285, 38374–38381. [Google Scholar] [CrossRef] [Green Version]
- Watkins, J.L.; Li, M.; McQuinn, R.P.; Chan, K.X.; McFarlane, H.E.; Ermakova, M.; Furbank, R.T.; Mares, D.; Dong, C.M.; Chalmers, K.J.; et al. A GDSL esterase/lipase catalyzes the esterification of lutein in bread wheat. Plant Cell 2019, 31, 3092–3112. [Google Scholar] [CrossRef]
- Wang, K.; Guo, Q.; Froehlich, J.E.; Hersh, H.L.; Zienkiewicz, A.; Howe, G.A.; Benning, C. Two abscisic acid-responsive plastid lipase genes involved in jasmonic acid biosynthesis in Arabidopsis thaliana. Plant Cell 2018, 30, 1006–1022. [Google Scholar] [CrossRef] [Green Version]
- Oh, I.S.; Park, A.R.; Bae, M.S.; Kwon, S.J.; Kim, Y.S.; Lee, J.E.; Kang, N.Y.; Lee, S.M.; Cheong, H.; Park, O.K. Secretome analysis reveals an Arabidopsis lipase involved in defense against Alternaria brassicicola. Plant Cell 2005, 17, 2832–2847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.G.; Kwon, S.J.; Jang, Y.J.; Nam, M.H.; Chung, J.H.; Na, Y.C.; Guo, H.W.; Park, O.K. GDSL LIPASE1 modulates plant immunity through feedback regulation of ethylene signaling. Plant Physiol. 2013, 163, 1776–1791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hubner, S.; Bercovich, N.; Todesco, M.; Mandel, J.R.; Odenheimer, J.; Ziegler, E.; Lee, J.S.; Baute, G.J.; Owens, G.L.; Grassa, C.J.; et al. Sunflower pan-genome analysis shows that hybridization altered gene content and disease resistance. Nat. Plants 2019, 5, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Aharoni, A.; Dixit, S.; Jetter, R.; Thoenes, E.; van Arkel, G.; Pereira, A. The SHINE clade of AP2 domain transcription factors activates wax biosynthesis, alters cuticle properties, and confers drought tolerance when overexpressed in Arabidopsis. Plant Cell 2004, 16, 2463–2480. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.X.; Malitsky, S.; De Oliveira, S.; Branigan, C.; Franke, R.B.; Schreiber, L.; Aharoni, A. SHINE transcription factors act redundantly to pattern the archetypal surface of Arabidopsis flower organs. PLoS Genet. 2011, 7, e1001388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, J.X.; Adato, A.; Alkan, N.; He, Y.H.; Lashbrooke, J.; Matas, A.J.; Meir, S.; Malitsky, S.; Isaacson, T.; Prusky, D.; et al. The tomato SlSHINE3 transcription factor regulates fruit cuticle formation and epidermal patterning. New Phytol. 2013, 197, 468–480. [Google Scholar] [CrossRef]
- Jung, K.H.; Han, M.J.; Lee, Y.S.; Kim, Y.W.; Hwang, I.W.; Kim, M.J.; Kim, Y.K.; Nahm, B.H.; An, G.H. Rice Undeveloped Tapetum1 is a major regulator of early tapetum development. Plant Cell 2005, 17, 2705–2722. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Yuan, Z.; Vizcay-Barrena, G.; Yang, C.Y.; Liang, W.Q.; Zong, J.; Wilson, Z.A.; Zhang, D.B. PERSISTENT TAPETAL CELL1 encodes a PHD-finger protein that is required for tapetal cell death and pollen development in rice. Plant Physiol. 2011, 156, 615–630. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W. A GDSL lipase is required for anther and pollen development. Plant Physiol. 2020, 182, 1810–1811. [Google Scholar] [CrossRef] [Green Version]
- Miao, X.Y.; Qu, H.P.; Han, Y.L.; He, C.F.; Qiu, D.W.; Cheng, Z.W. The protein elicitor Hrip1 enhances resistance to insects and early bolting and flowering in Arabidopsis thaliana. PLoS ONE 2019, 14, e0216082. [Google Scholar] [CrossRef]
- Han, X.F.; Li, S.; Zhang, M.; Yang, L.Y.; Liu, Y.D.; Xu, J.; Zhang, S.Q. Regulation of GDSL lipase gene expression by the MPK3/MPK6 cascade and its downstream WRKY transcription factors in Arabidopsis immunity. Mol. Plant Microbe 2019, 32, 673–684. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Species | Numbers | Reference |
|---|---|---|
| Arabidopsis thaliana | 105 | [4] |
| Brassica rapa | 121 | [6] |
| Fragaria vesca | 96 | [10] |
| Glycine max | 194 | [2,9] |
| Malus domestica | 150 | [10] |
| Oryza sativa | 114 | [5] |
| Prunus avium | 70 | [10] |
| Prunus mume | 90 | [10] |
| Prunus persica | 97 | [10] |
| Pyrus bretschneideri | 94 | [10] |
| Vitis vinifera | 83 | [7] |
| Zea mays | 103 | [8] |
| Crop Species | Reported GELPs | Process of Development | Reference |
|---|---|---|---|
| Arabidopsis thaliana | CDEF1 | Lateral root development (auxin signaling-mediated); Pollen-stigma interaction and pollinator attraction; Lipid metabolism | [23] |
| CUS2 | Flower development | [31] | |
| DAD1 | Flower development; Anther and pollen development | [29] | |
| EXL4 | Pollen-stigma interaction and pollinator attraction; Lipid metabolism | [43] | |
| EXL6 | Anther and pollen development | [6,38] | |
| GELP15, GELP39 | Flower development | [32,33] | |
| GELP22, GELP38, GELP49, GELP51 and GELP96 GELP12, DELP55, GELP72, GELP73 and GELP81 | Lateral root development (auxin signaling-mediated) | [21] | |
| GELP77 | Anther and pollen development | [37] | |
| GER1 | Coleoptile elongation; Lateral root development (auxin signaling-mediated) | [20,22] | |
| GLIP1 | Plant-biotic stress interaction | [62] | |
| GLIP2 | Lipid metabolism; Plant-biotic stress interaction | [22] | |
| LIP1 | Seed germination | [19] | |
| OSP1 | Plant-abiotic stress interaction; Stomata development | [28] | |
| PLIP2, PLIP3 | Plant-abiotic stress interaction | [61] | |
| RVMS | Anther and pollen development | [36] | |
| SFAR4 | Lipid metabolism; Plant-abiotic stress interaction | [51] | |
| WDL1 | Plant height; Stomata development | [26] | |
| Brassica napus | BnSCE3, BnLIP2 | Seed germination; Seed development | [18] |
| Capsicum annuum | CaGLIP1 | Plant-abiotic/biotic stress interaction; Lipid metabolism | [50] |
| Glycine max | GmGELP28 | Plant-abiotic stress interaction | [9] |
| Gossypium hirsutum | GhGDSL | Cell wall metabolism | [56] |
| GhGLIP | Seed development | [48] | |
| Jacaranda mimosifolia | JNP1 | Pollen-stigma interaction and pollinator attraction; Lipid metabolism | [44] |
| Oraza sativa | BS1 | Cell wall metabolism | [55] |
| DARX1 | Cell wall metabolism | [54] | |
| DR | Plant height | [26] | |
| MHZ11 | Root development | [24] | |
| OSGELP34, OsGELP110, OsGELP115 | Anther and pollen development | [40,41] | |
| OsGLIP1, OsGLIP2 | Plant-biotic stress interaction; Lipid metabolism | [52] | |
| RMS2 | Anther and pollen development; Lipid metabolism; Secondary metabolism | [39] | |
| WDL1 | Plant-abiotic stress interaction | [27] | |
| Solanum lycopersicum | CD1 | Secondary metabolism; Flower development; Fruit development; Plant-abiotic stress interaction | [14,30,47,57] |
| CUS1 | Flower development; Fruit development | [14,30] | |
| GDSL1 | Fruit development; Plant-abiotic stress interaction; Secondary metabolism | [46] | |
| SlCGT | Secondary metabolism | [59] | |
| SlGDSL2/SlCD1 | Fruit development; Plant-abiotic stress interaction | [47,57] | |
| Sl-LIP8 | Pollen-stigma interaction and pollinator attraction; Lipid metabolism | [45] | |
| Tanacetum cinerariifolium | TcGLIP | Secondary metabolism | [58] |
| Zea mays | IPE2 | Anther and pollen development | [42] |
| ZmMs30 | Anther and pollen development; Lipid metabolism | [8] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, G.; Sun, W.; Chen, Z.; Shi, L.; Hong, J.; Shi, J. Plant GDSL Esterases/Lipases: Evolutionary, Physiological and Molecular Functions in Plant Development. Plants 2022, 11, 468. https://doi.org/10.3390/plants11040468
Shen G, Sun W, Chen Z, Shi L, Hong J, Shi J. Plant GDSL Esterases/Lipases: Evolutionary, Physiological and Molecular Functions in Plant Development. Plants. 2022; 11(4):468. https://doi.org/10.3390/plants11040468
Chicago/Turabian StyleShen, Gaodian, Wenli Sun, Zican Chen, Lei Shi, Jun Hong, and Jianxin Shi. 2022. "Plant GDSL Esterases/Lipases: Evolutionary, Physiological and Molecular Functions in Plant Development" Plants 11, no. 4: 468. https://doi.org/10.3390/plants11040468