
Toxicity, Deterrent and Repellent Activities of Four Essential Oils on Aphis punicae (Hemiptera: Aphididae)

 ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Essential Oil Extraction
2.2. Insects
2.3. GC–MS Analysis of Essential Oils
2.4. Contact Toxicity
2.5. Deterrent Test
2.6. Repellent Test
2.7. Statistical Analysis
3. Results
3.1. Yields and Chemical Composition of Essential Oils
3.2. Effect of Contact Toxicity
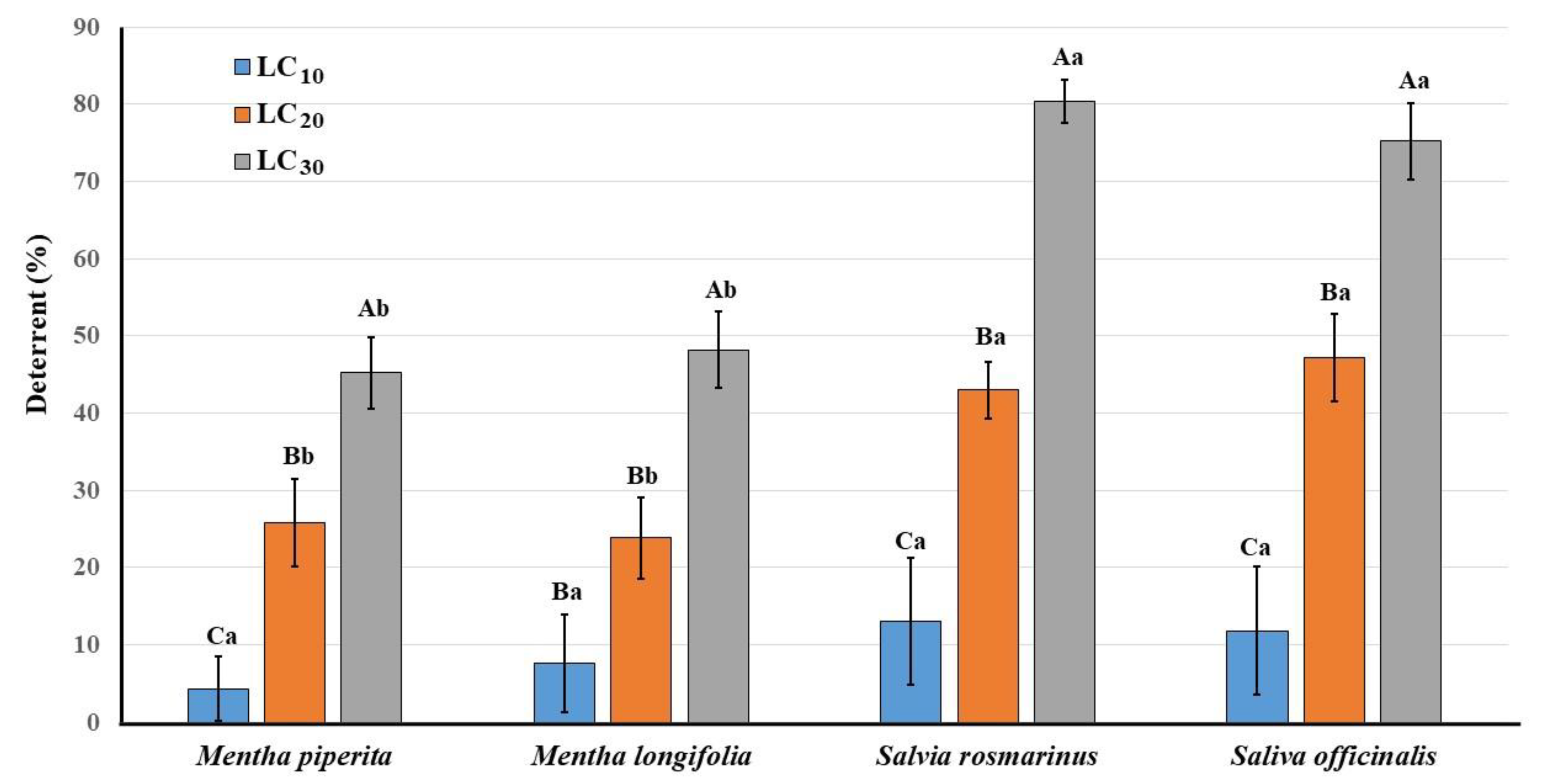
3.3. Deterrent Activity
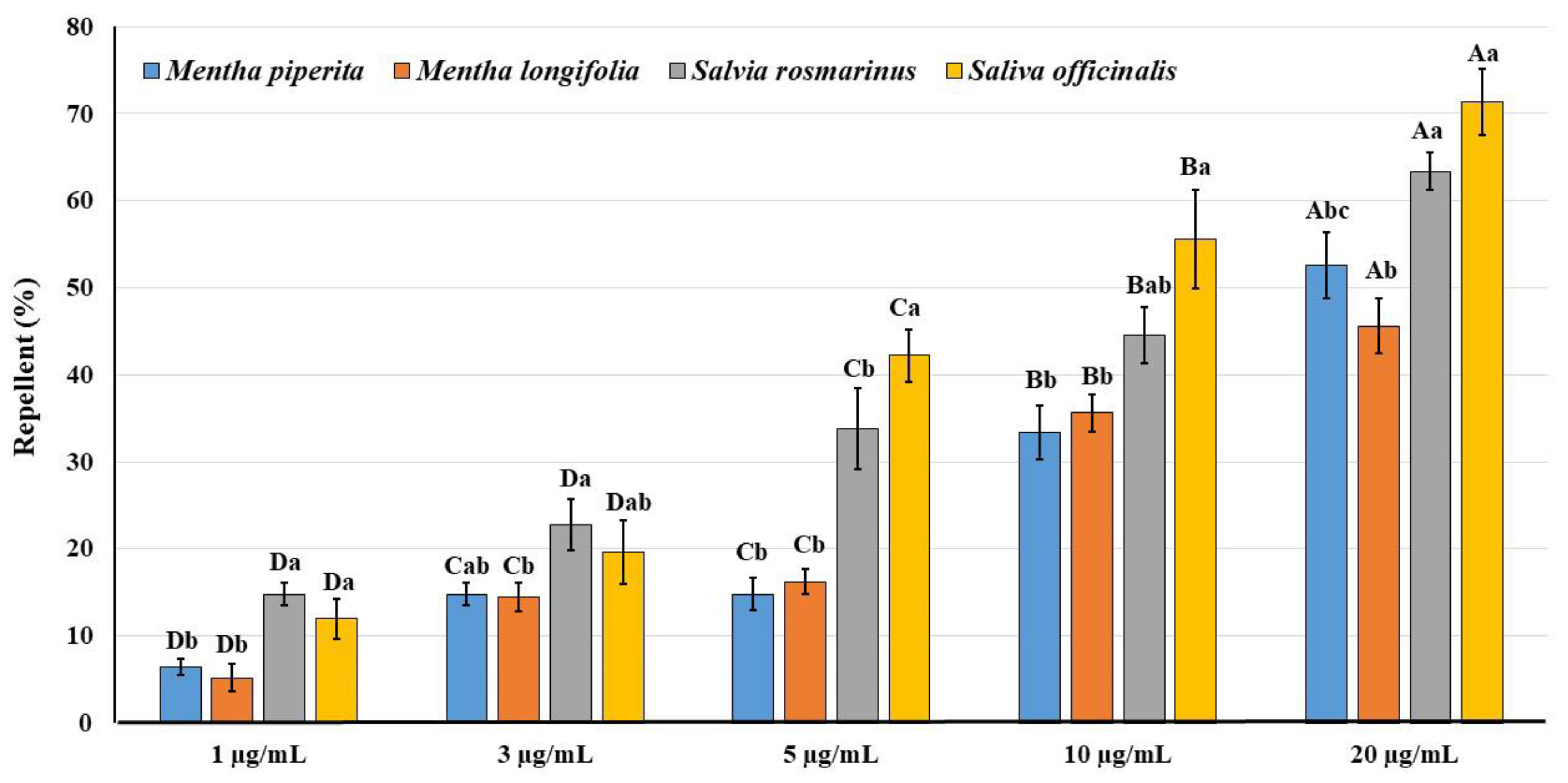
3.4. Repellent Activity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Werrie, P.Y.; Durenne, B.; Delaplace, P.; Fauconnier, M.L. Phytotoxicity of essential oils: Opportunities and constraints for the development of biopesticides. A review. Foods 2020, 9, 1291. [Google Scholar] [CrossRef]
- Roy, D.N.; Goswami, R.; Pal, A. The insect repellents: A silent environmental chemical toxicant to the health. Environ. Toxicol. Pharmacol. 2017, 50, 91–102. [Google Scholar] [CrossRef]
- Costa, L.G. Organophosphorus compounds at 80: Some old and new issues. Toxicol. Sci. 2018, 162, 24–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benelli, G.; Pavela, R.; Maggi, F.; Petrelli, R.; Nicoletti, M. Commentary: Making green pesticides greener? The potential of plant products for nanosynthesis and pest control. J. Cluster. Sci. 2017, 28, 3–10. [Google Scholar] [CrossRef]
- Isman, M.B. Botanical insecticides in the twenty-first century-fulfilling their promise? Annu. Rev. Èntomol. 2019, 65, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikbal, C.; Pavela, R. Essential oils as active ingredients of botanical insecticides against aphids. J. Pest. Sci. 2019, 92, 971–986. [Google Scholar] [CrossRef]
- Regnault-Roger, C.; Vincent, C.; Arnason, J.T. Essential oils in insect control: Low-risk products in a highstakes world. Annu. Rev. Entomol. 2012, 57, 405–424. [Google Scholar] [CrossRef] [PubMed]
- Hikal, W.; Baeshen, R.S.; Ahl, H.A. Botanical insecticide as simple extractives for pest control. Cogent Biol. 2017, 3, 1404274. [Google Scholar] [CrossRef]
- Isman, M.B. Botanical insecticides, deterrents, and repellents in modern agriculture and an increasingly regulated world. Annu. Rev. Entomol. 2006, 51, 45–66. [Google Scholar] [CrossRef] [Green Version]
- Bohinc, T.; Horvat, A.; Ocvirk, M.; Košir, I.J.; Rutnik, K.; Trdan, S. The first evidence of the insecticidal potential of plant powders from invasive alien plants against rice weevil under laboratory conditions. Appl. Sci. 2020, 10, 7828. [Google Scholar] [CrossRef]
- Farajzadeh, M.A.; Khoshmaram, L.; Nabil, A.A.A. Determination of pyrethroid pesticides residues in vegetable oils using liquid-liquid extraction and dispersive liquid-liquid microextraction followed by gas chromatography-flame ionization detection. J. Food Compos. Anal. 2014, 34, 128–135. [Google Scholar] [CrossRef]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological effects of essential oils: A review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef] [PubMed]
- Abd-Ella, A.A. Effect of several insecticides on pomegranate aphid, Aphis punicae (Passerini) (Homoptera: Aphididae) and its predators under field conditions. EPPO Bull. 2015, 45, 90–96. [Google Scholar] [CrossRef]
- Sreedevi, K.; Verghese, A. Seasonal incidence of aphid, Aphis punicae Passerini (Homoptera: Aphididae) and its relationship with abiotic factors. Pest. Manag. Hortic. Ecosys. 2007, 13, 165–171. [Google Scholar]
- Hawry, M.A.; Skalicka-Woźniak, K.; Świeboda, R.; Niemiec, M.; Stępak, K.; Waksmundzka-Hajnos, M.; Hawry, A.; Szymczak, G. GC-MS fingerprints of mint essential oils. Open Chem. 2015, 13, 1326–1332. [Google Scholar] [CrossRef]
- Hussain, A.I.; Anwar, F.; Nigam, P.S.; Ashraf, M.; Gilanif, A.H. Seasonal variation in content, chemical composition and antimicrobial and cytotoxic activities of essential oils from four Mentha species. J. Sci. Food Agric. 2010, 90, 1827–1836. [Google Scholar] [CrossRef]
- Walker, J.B.; Sytsma, K.J. Staminal evolution in the genus Salvia (Lamiaceae): Molecular phylogenetic evidence for multiple origins of the staminal lever. Ann. Bot. 2007, 100, 375–391. [Google Scholar] [CrossRef]
- De Macedo, L.M.; Santos, É.M.D.; Militão, L.; Tundisi, L.L.; Ataide, J.A.; Souto, E.B.; Mazzola, P.G. Rosemary (Rosmarinus officinalis L., syn Salvia rosmarinus Spenn.) and its topical applications: A review. Plants 2020, 9, 651. [Google Scholar] [CrossRef] [PubMed]
- Tanoh, E.A.; Boué, G.B.; Nea, F.; Genva, M.; Wognin, E.L.; Ledoux, A.; Martin, H.; Tonzibo, Z.F.; Frederich, M.; Fauconnier, M.-L. Seasonal effect on the chemical composition, insecticidal properties and other biological activities of Zanthoxylum leprieurii Guill. & Perr. essential oils. Foods 2020, 9, 550. [Google Scholar]
- Singh, S.; Das, S.S.; Singh, G.; Perotti, M.; Schuff, C.; Catalán, C.A.N. In vitro antioxidant potentials and chemistry of essential oils and oleoresins from fresh and sun-dried Mentha longifolia L. J. Essent. Oil. Res. 2015, 27, 61–69. [Google Scholar] [CrossRef]
- Sayed, S.M. Molecular diversity of the lady beetles, Coccinella undecimpunctata L. and Hippodamia variegata (Goeze) (Coleoptera: Coccinellidae) in Saudi Arabia. Egypt. J. Biol. Pest Control. 2016, 26, 351–355. [Google Scholar]
- Hasanshahi, G.; Abbasipour, H.; Jahan, F.; Askarianzadeh, A.; Karimi, J.; Rastegar, F. Fumigant toxicity and nymph production deterrence effect of three essential oils against two aphid species in the laboratory condition. J. Essent. Oil Bear. Plants 2016, 19, 706–711. [Google Scholar] [CrossRef]
- Tunc, I.; Erler, F. Repellency and repellent stability of essential oil constituents against Tribolium confusum. J. Plant. Dis. Prot. 2003, 110, 394–400. [Google Scholar]
- Kumar, N.; Kumar, S.; Vats, S.K.; Ahuja, P.S. Effect of elevation on the primary products of photosynthesis and the associated enzymes in barley and wheat. Photosynth. Res. 2006, 88, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Robertson, J.L.; Savin, N.; Russell, R.M.; Preisler, H.K. Bioassays with Arthropods; CRC Press: Boca Raton, FL, USA, 2007; p. 224. [Google Scholar]
- Chrysargyris, A.; Mikallou, M.; Petropoulos, S.; Tzortzakis, N. Profiling of essential oils components and polyphenols for their antioxidant activity of medicinal and aromatic plants grown in different environmental conditions. Agronomy 2020, 10, 727. [Google Scholar] [CrossRef]
- Daferera, D.; Ziogas, B.; Polissiou, M. GC-MS analysis of essential oils from some Greek aromatic plants and their fungitoxicity on Penicillium digitatum. J. Agric. Food Chem. 2000, 48, 2576–2581. [Google Scholar] [CrossRef]
- Goze, I.; Vural, N.; Ercan, N. Characterization of essential oil and antioxidant activities of some species of Salvia in Turkey. Nat. Volatiles Essent. Oils 2016, 3, 1–7. [Google Scholar]
- Kimbaris, A.C.; Papachristos, D.P.; Michaelakis, A.; Martinou, A.F.; Polissiou, M.G. Toxicity of plant essential oil vapours to aphid pests and their coccinellid predators. Biocontrol Sci. Technol. 2010, 20, 411–422. [Google Scholar] [CrossRef]
- Ainane, A.; Khammour, F.; Elkouali, M.; Talbi, M.; Abba, E.H.; Cherroud, S.; Elyaacoubi, A.; Oumaskour, K.; Boujaber, N.; Benhar, S.; et al. Chemical study by GC-MS of the essential oils of certain mints grown in the region of Settat (Morocco): Mentha Piperita, Mentha Pulegium and Mentha Spicata. Drug Des. Int. Prop. Int. J. 2018, 1, 124–127. [Google Scholar] [CrossRef]
- Turek, C.; Stintzing, F.C. Impact of different storage conditions on the quality of selected essential oils. Food Res. Int. 2012, 46, 341–353. [Google Scholar] [CrossRef]
- Glare, T. Types of biopesticides. In Biopesticides Handbook; Leo Nollet, L.M.L., Ed.; CRC Press: New York, NY, USA, 2015; pp. 7–25. [Google Scholar]
- Haddi, K.; Turchen, L.M.; Viteri Jumbo, L.O.; Guedes, R.N.; Pereira, E.J.; Aguiar, R.W.; Oliveira, E.E. Rethinking biorational insecticides for the pest management: Unintended effects and consequences. Pest Manag. Sci. 2020, 76, 2286–2293. [Google Scholar] [CrossRef] [PubMed]
- Durán-Lara, E.F.; Valderrama, A.; Marican, A. Natural organic compounds for application in organic farming. Agriculture 2020, 10, 41. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Zhang, W.J.; Wang, P.J.; Yang, K.; Huang, D.Y.; Wei, J.Y.; Tian, Z.F.; Bai, J.F.; Du, S.S. Contact toxicity and repellency of the essential oil of Liriope muscari (DECN.) Bailey against three insect tobacco storage pests. Molecules 2015, 20, 1676–1685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.C.; Liang, Y.; Shi, W.P.; Liu, Q.Z.; Zhou, L.; Liu, Z.L. Repellent and insecticidal effects of the essential oil of Kaempferia galanga rhizomes to Liposcelis bostrychophila (Psocoptera: Liposcelidae). J. Econ. Entomol. 2014, 107, 1706–1712. [Google Scholar] [CrossRef] [PubMed]
- Ingkaninan, K.; Temkitthawon, P.; Chuenchom, K.; Yuyaem, T.; Thongnoi, W. Screening for acetylcholinesterase inhibitory activity in plants used in Thai traditional rejuvenating and neurotonic remedies. J. Ethnopharmacol. 2003, 89, 261–264. [Google Scholar] [CrossRef]
- Tong, F.; Coats, J.R. Effects of monoterpenoid insecticides on [3H]-TBOB binding in house fly GABA receptor and 36Cl− uptake in American cockroach ventral nerve cord. Pestic. Biochem. Physiol. 2010, 98, 317–324. [Google Scholar] [CrossRef]
- Jankowska, M.; Rogalska, J.; Wyszkowska, J.; Stankiewicz, M. Molecular targets for components of essential oils in the insect nervous system—A review. Molecules 2018, 23, 34. [Google Scholar] [CrossRef] [Green Version]
- Pavela, R. Acute toxicity and synergistic and antagonistic effects of the aromatic compounds of some essential oils against Culex quinquefasciatus Say larvae. Parasitol. Res. 2015, 114, 3835–3853. [Google Scholar] [CrossRef]
- Digilio, M.C.; Mancini, E.; Voto, E.; De Feo, V. Insecticide activity of Mediterranean essential oils. J. Plant Inter. 2008, 3, 17–23. [Google Scholar] [CrossRef]
- Tak, J.H.; Jovel, E.; Isman, M.B. Effects of rosemary, thyme and lemongrass oils and their major constituents on detoxifying enzyme activity and insecticidal activity in Trichoplusia ni. Pest. Biochem. Phys. 2017, 140, 9–16. [Google Scholar] [CrossRef]
- Koorki, Z.; Shahidi-Noghabi, S.; Mahdian, K.; Pirmaoradi, M. Chemical composition and insecticidal properties of several plant essential oils on the melon aphid, Aphis gossypii glover (Hemiptera: Aphididae). J. Essent. Oil Bear. Plants 2018, 21, 420–429. [Google Scholar] [CrossRef]
- Liang, J.Y.; Xu, J.; Yang, Y.Y.; Shao, Y.Z.; Zhou, F.; Wang, J.L. Toxicity and synergistic effect of Elsholtzia Ciliata essential oil and its main components against the adult and larval stages of Tribolium Castaneum. Foods 2020, 16, 345. [Google Scholar] [CrossRef] [Green Version]
- Ebadollahi, A.; Setzer, W.N. Evaluation of the toxicity of Satureja intermedia C. A. Mey essential oil to storage and greenhouse insect pests and a predator ladybird. Foods 2020, 9, 712. [Google Scholar] [CrossRef] [PubMed]
- Abramson, C.I.; Wanderley, P.A.; Wanderley, M.J.A.; Miná, A.J.S.; de Souza, O.B. Effect of essential oil from citronella and alfazema on fennel aphids Hyadaphis foeniculi Passerini (Hemiptera: Aphididae) and its predator Cycloneda sanguinea L. (Coleoptera: Coccinelidae). Am. J. Environ. Sci. 2007, 3, 9–10. [Google Scholar] [CrossRef]
- Seixas, P.T.L.; Demuner, A.J.; Alvarenga, E.S.; Barbosa, L.C.A.; Marques, A.; Farias, E.D.S.; Picanço, M.C. Bioactivity of essential oils from Artemisia against Diaphania hyalinata and its selectivity to beneficial insects. Sci. Agric. 2018, 75, 519–525. [Google Scholar] [CrossRef]
- Ngassoum, M.B.; Tinkeu, N.S.L.; Ngatanko, I.; Tapondjou, A.L.; Lognay, G.; Malaisse, F.; Hance, T. Chemical composition, insecticidal effect and repellent activity of essential oils of three aromatic plants, alone and in combination towards Sitophilus oryzae L. (Coleoptera: Curculionidae). Nat. Prod. Commun. 2007, 2, 1229–1232. [Google Scholar] [CrossRef] [Green Version]
- Toledo, P.F.S.; Ferreira, T.P.; Bastos, I.M.A.S.; Rezende, S.M.; Jumbo, L.O.V.; Didonet, J.; Andrade, B.S.; Melo, T.S.; Smagghe, G.; Oliveira, E.E.; et al. Essential oil from Negramina (Siparuna guianensis) plants controls aphids without impairing survival and predatory abilities of non-target ladybeetles. Environ. Pollut. 2019, 255, 113153. [Google Scholar] [CrossRef] [PubMed]
- Hori, M.; Komatsu, H. Repellency of rosemary oil and its components against the onion aphid Neotoxoptera formosana (Takahashi) (Homoptera, Aphididae). App. Entomol. Zool. 1997, 32, 303–310. [Google Scholar] [CrossRef] [Green Version]
- Erdemir, T.; Erler, F. Repellent, oviposition-deterrent and egg-hatching inhibitory effects of some plant essential oils against citrus mealybug, Planococcus citri Risso (Hemiptera: Pseudococcidae). J. Plant. Dis. Prot. 2017, 124, 473–479. [Google Scholar] [CrossRef]
- Rojht, H.; Košir, I.J.; Trdan, S. Chemical analysis of three herbal extracts and observation of their activity against adults of Acanthoscelides obtectus and Leptinotarsa decemlineata using a video tracking system. J. Plant Dis. Prot. 2012, 119, 59–67. [Google Scholar] [CrossRef]
- Pascual-Villalobos, M.; Cantó-Tejero, M.; Vallejo, R.; Guirao, P.; RodríguezRojo, S.; Cocero, M.J. Use of nanoemulsions of plant essential oils as aphid repellents. Ind. Crops Prod. 2017, 110, 45–57. [Google Scholar] [CrossRef]
- Jahan, F.; Abbasipour, H.; Hasanshahi, G. Fumigant toxicity and nymph production deterrence effect of five essential oils on adults of the cabbage aphid, Brevicoryne brassicae L. (Hemiptera: Aphididae). J. Essent. Oil Bear. Plants 2016, 19, 1111–1118. [Google Scholar] [CrossRef]
- Işık, M.; Görür, G. Aphidicidial activity of seven essential oils against the cabbage aphid, Brevicoryne brassicae L. (Hemiptera: Aphididae). Mun. Ent. Zool. 2009, 4, 424–431. [Google Scholar]



{kind=link}
{kind=link}
| R.T. | Compound | Area (%) | Formula | |||
|---|---|---|---|---|---|---|
| Mentha piperita | Mentha longifolia | Salvia officinalis | Salvia rosmarinus | |||
| 3.32 | α-Phellandrene | 3.46 | C10H16 | |||
| 3.44 | 3-Thujene | 1.97 | 1.00 | C10H16 | ||
| 3.63 | α-pinene | 2.05 | 1.43 | 12.26 | 30.09 | C10H16 |
| 4.00 | Camphene | 14.44 | 6.80 | C10H16 | ||
| 4.05 | 2,4(10)-Thujadiene | - | 2.06 | C10H14 | ||
| 4.08 | 3-Octanol | 0.27 | C8H18O | |||
| 4.47 | α-Phellandrene | 0.60 | C10H16 | |||
| 4.49 | α-Phellandrene | 1.74 | - | C10H16 | ||
| 4.63 | α -Pinene | 10.42 | 6.54 | C10H16 | ||
| 4.71 | Eucalyptol | 2.09 | C10H18O | |||
| 4.79 | 1-Octen-3-ol | 0.65 | - | C8H16O | ||
| 4.82 | β-Pinene | 5.99 | 5.21 | C10H16 | ||
| 4.97 | α-Ocimene | 2.34 | C10H16 | |||
| 5.59 | α-Terpinene | 0.43 | 2.05 | C10H16 | ||
| 5.98 | D-Limonene | 4.08 | 0.75 | 3.71 | 5.23 | C10H16 |
| 6.08 | p-Cymene | - | 7.08 | C10H14 | ||
| 6.10 | o-Cymene | 0.75 | - | C10H14 | ||
| 6.45 | Eucalyptol | 4.50 | 33.52 | 6.91 | C10H18O | |
| 6.84 | γ-Terpinene | 1.14 | 6.01 | C10H16 | ||
| 7.07 | α-Ocimene | 0.23 | - | C10H16 | ||
| 7.70 | α-Terpinolene | 2.65 | C10H16 | |||
| 8.35 | Linalool | - | 1.06 | C10H18O | ||
| 9.46 | endo-Borneol | 0.50 | C10H18O | |||
| 9.90 | Sabinyl acetate | 0.15 | C10H16O | |||
| 10.56 | (+)-2-Bornanone | 11.58 | 0.62 | C10H16O | ||
| 10.60 | l-Menthone | 34.49 | C10H18O | |||
| 11.07 | Isomenthol | 0.35 | C10H20O | |||
| 11.16 | endo-Borneol | - | 2.55 | C10H18O | ||
| 11.26 | Thujone | 0.59 | - | C10H16O | ||
| 11.44 | dl-Menthol | 2.75 | C10H20O | |||
| 11.45 | Terpinen-4-ol | - | 0.48 | C10H18O | ||
| 12.02 | Carveol | 1.77 | C10H16O | |||
| 12.14 | α-Terpineol | - | 0.68 | C10H18O | ||
| 12.20 | α-Terpineol | 0.77 | C10H18O | |||
| 12.51 | Carvone | 61.16 | 3.77 | C10H14O | ||
| 12.67 | l-Verbenone | - | 0.87 | C10H14O | ||
| 14.05 | Pulegone | 48.60 | C10H16O | |||
| 14.38 | Geraniol | - | 2.20 | C10H18O | ||
| 15.55 | (-)-Bornyl acetate | - | 4.54 | C12H20O2 | ||
| 14.64 | Piperitone | 0.46 | C10H16O | |||
| 15.32 | Neocarveol | 2.28 | C10H18O | |||
| 16.77 | trans-Carveyl acetate | 3.64 | C12H18O2 | |||
| 17.01 | (-)-β-Bourbonene | 1.83 | C15H24 | |||
| 18.49 | Caryophyllene | 0.65 | C15H24 | |||
| 18.82 | trans-Verbenone | 0.51 | C10H14O | |||
| 19.17 | 1-Pentanol, 4-amino- | 0.23 | - | C5H13NO | ||
| 19.65 | Geranyl acetate | - | 1.04 | C12H20O2 | ||
| 20.32 | (+)-epi-Bicyclosesquiphellandrene | 0.58 | C15H24 | |||
| 20.83 | Caryophyllene | - | 1.64 | C15H24 | ||
| 20.85 | Caryophyllene | 0.39 | C15H24 | |||
| 21.16 | α -Cubebene | 10.99 | C15H24 | |||
| 22.20 | β-Elemen | 0.97 | C15H24 | |||
| 22.34 | Humulene | - | 0.90 | C15H24 | ||
| 22.88 | trans-calamenene | 0.46 | C15H22 | |||
| 26.81 | Cubenol | 0.22 | C15H26O | |||
| 27.48 | Caryophyllene oxide | - | 0.50 | C15H24O | ||
| 28.00 | ç-Muurolene | 0.38 | C15H24 | |||
| Total | 99.04 | 99.52 | 99.65 | 99.39 | ||
| Number of compounds | 17 | 14 | 16 | 26 | ||
| Tested Insect | Essential Oil | LC50 (Confidence Interval Limits) | S.R. * | Intercept ± SE | Slope ± SE | ꭓ2 | P |
|---|---|---|---|---|---|---|---|
| Aphis punicae | Mentha piperita | 2.971 (2.376–3.504) | −0.782 ± 0.126 | 0.263 ± 0.029 | 0.675 | 0.879 | |
| Mentha longifolia | 2.400 (1.750–2.945) | −0.642 ± 0.127 | 0.268 ± 0.030 | 4.443 | 0.217 | ||
| Salvia officinalis | 1.574 (0.940–2.043) | −0.563 ± 0.151 | 0.358 ± 0.047 | 1.001 | 0.801 | ||
| Salvia rosmarinus | 1.653 (0.945–2.190) | −0.497 ± 0.136 | 0.301 ± 0.037 | 6.046 | 0.109 | ||
| Coccinella undecimpunctata | Mentha piperita | 10.334 (6.513–16.716) | 3.478 | −0.970 ± 0.095 | 0.094 ± 0.009 | 9.898 | 0.019 |
| Mentha longifolia | 8.737 (3.862–15.949) | 3.640 | −0.840 ± 0.094 | 0.096 ± 0.010 | 13.583 | 0.004 | |
| Salvia officinalis | 6.237 (5.452–7.076) | 3.963 | −1.022 ± 0.106 | 0.164 ± 0.015 | 2.036 | 0.565 | |
| Salvia rosmarinus | 5.960 (1.764–14.809) | 3.606 | −0.877 ± 0.100 | 0.147 ± 0.013 | 26.640 | 0.001 |
| Plant Extract | Mentha piperita | Mentha longifolia | Salvia officinalis | Salvia rosmarinus |
|---|---|---|---|---|
| Mentha piperita | −0.933 | −2.728 | −3.484 | |
| Mentha longifolia | 0.538 | −1.795 | −2.551 | |
| Salvia officinalis | 1.705 | 1.166 | 0.756 | |
| Salvia rosmarinus | 1.509 | 0.971 | −0.195 |
| Source | Type III Sum of Squares | Df | Mean Square | F | Sig. |
|---|---|---|---|---|---|
| Corrected Model | 35,456.85 | 11 | 3223.35 | 20.35 | <0.001 |
| Intercept | 75,437.60 | 1 | 75,437.60 | 476.27 | <0.001 |
| Plant species | 5597.15 | 3 | 1865.72 | 11.78 | <0.001 |
| Concentration | 28,177.31 | 2 | 14,088.65 | 88.95 | <0.001 |
| Plant species x Concentration | 1682.39 | 6 | 280.40 | 1.77 | 0.125 |
| Error | 7602.80 | 48 | 158.39 | ||
| Total | 11,8497.25 | 60 | |||
| Corrected Total | 43,059.65 | 59 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sayed, S.; Soliman, M.M.; Al-Otaibi, S.; Hassan, M.M.; Elarrnaouty, S.-A.; Abozeid, S.M.; El-Shehawi, A.M. Toxicity, Deterrent and Repellent Activities of Four Essential Oils on Aphis punicae (Hemiptera: Aphididae). Plants 2022, 11, 463. https://doi.org/10.3390/plants11030463
Sayed S, Soliman MM, Al-Otaibi S, Hassan MM, Elarrnaouty S-A, Abozeid SM, El-Shehawi AM. Toxicity, Deterrent and Repellent Activities of Four Essential Oils on Aphis punicae (Hemiptera: Aphididae). Plants. 2022; 11(3):463. https://doi.org/10.3390/plants11030463
Chicago/Turabian StyleSayed, Samy, Mohamed Mohamed Soliman, Saad Al-Otaibi, Mohamed M. Hassan, Sayed-Ashraf Elarrnaouty, Samia M. Abozeid, and Ahmed M. El-Shehawi. 2022. "Toxicity, Deterrent and Repellent Activities of Four Essential Oils on Aphis punicae (Hemiptera: Aphididae)" Plants 11, no. 3: 463. https://doi.org/10.3390/plants11030463