
Environmental Factors Driving the Spatial Distribution Pattern of Venerable Trees in Sichuan Province, China

Abstract
:1. Introduction
2. Results
2.1. Species Composition and Importance Value
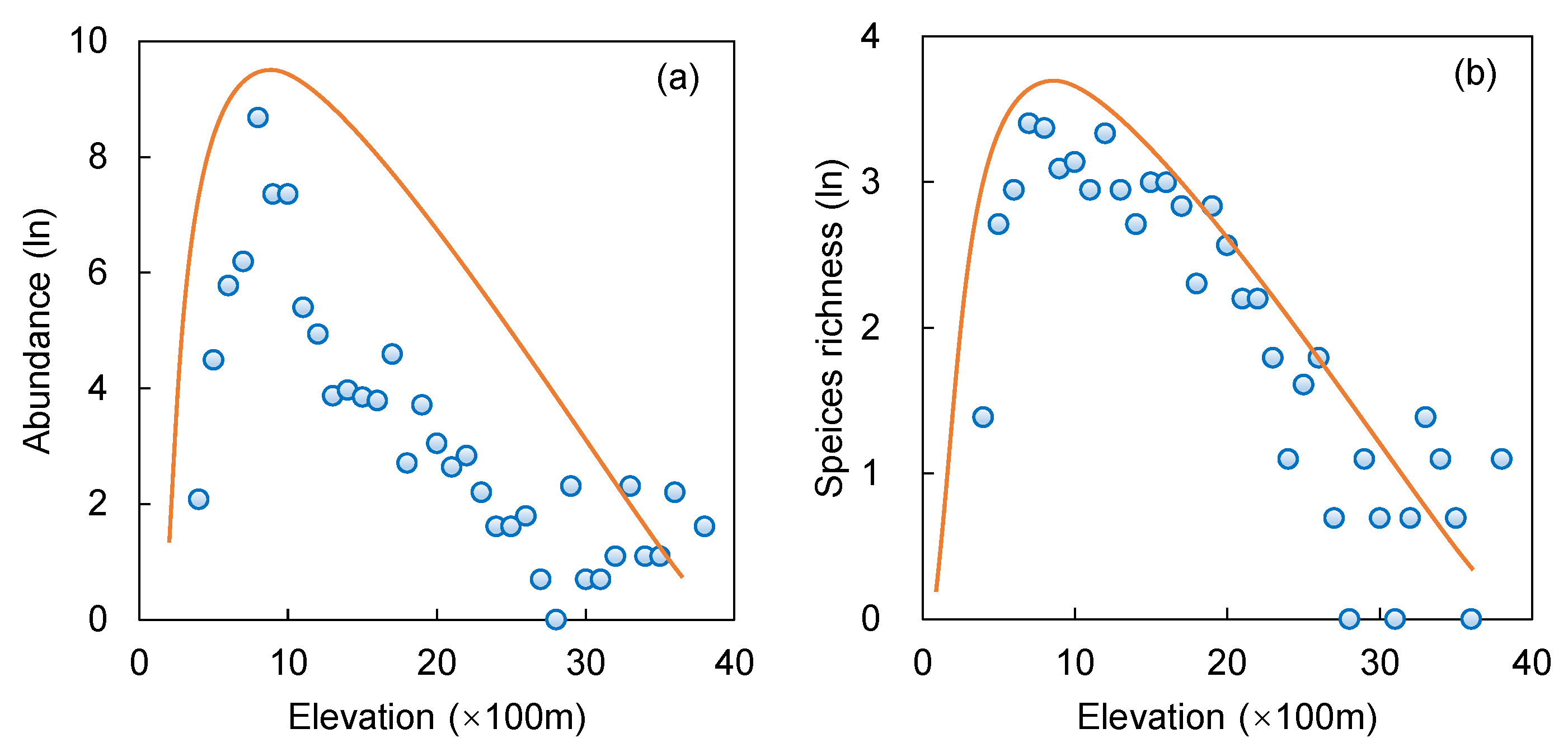
2.2. Elevation Distribution Patterns
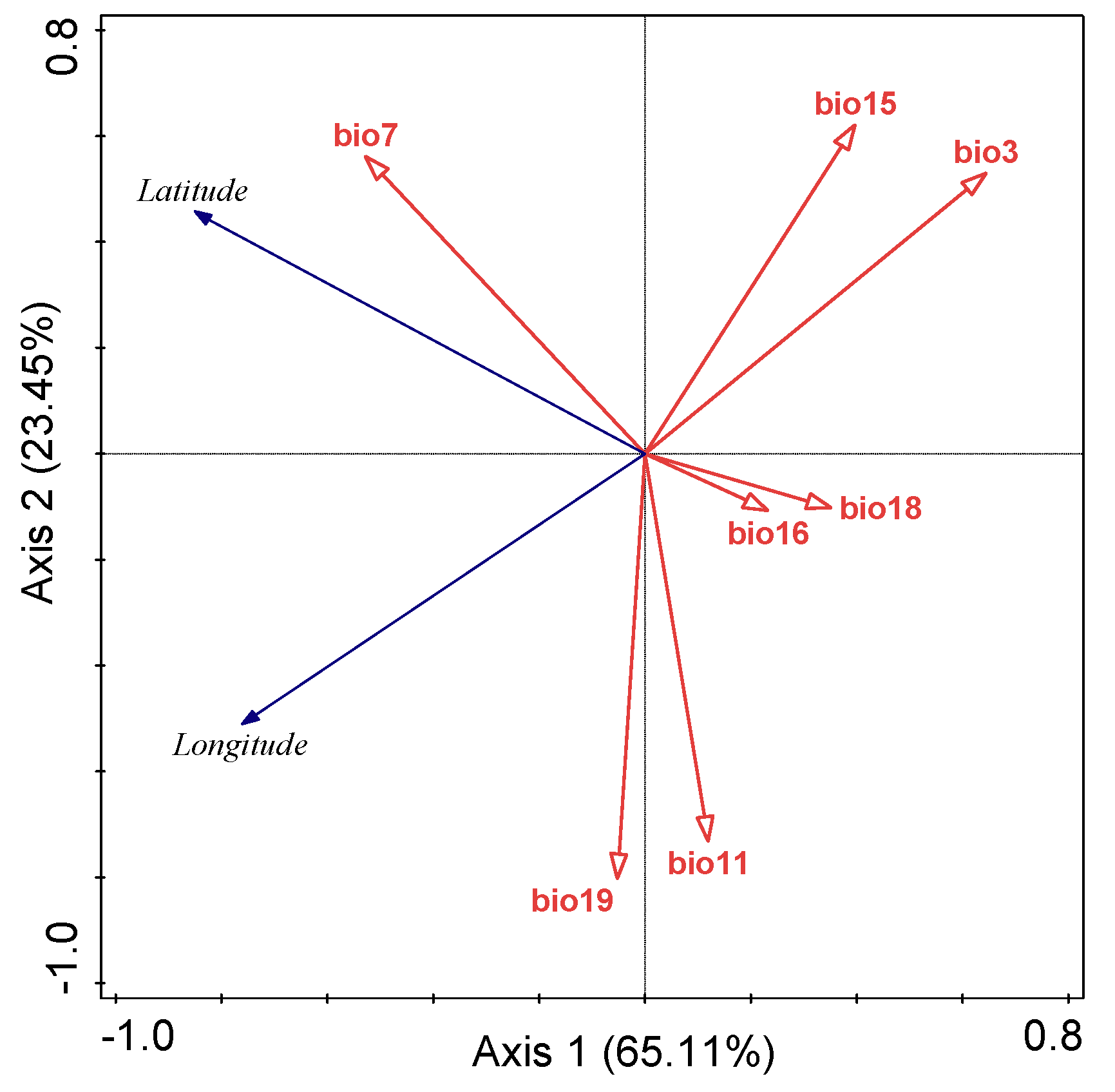
2.3. Key Bioclimatic Factors
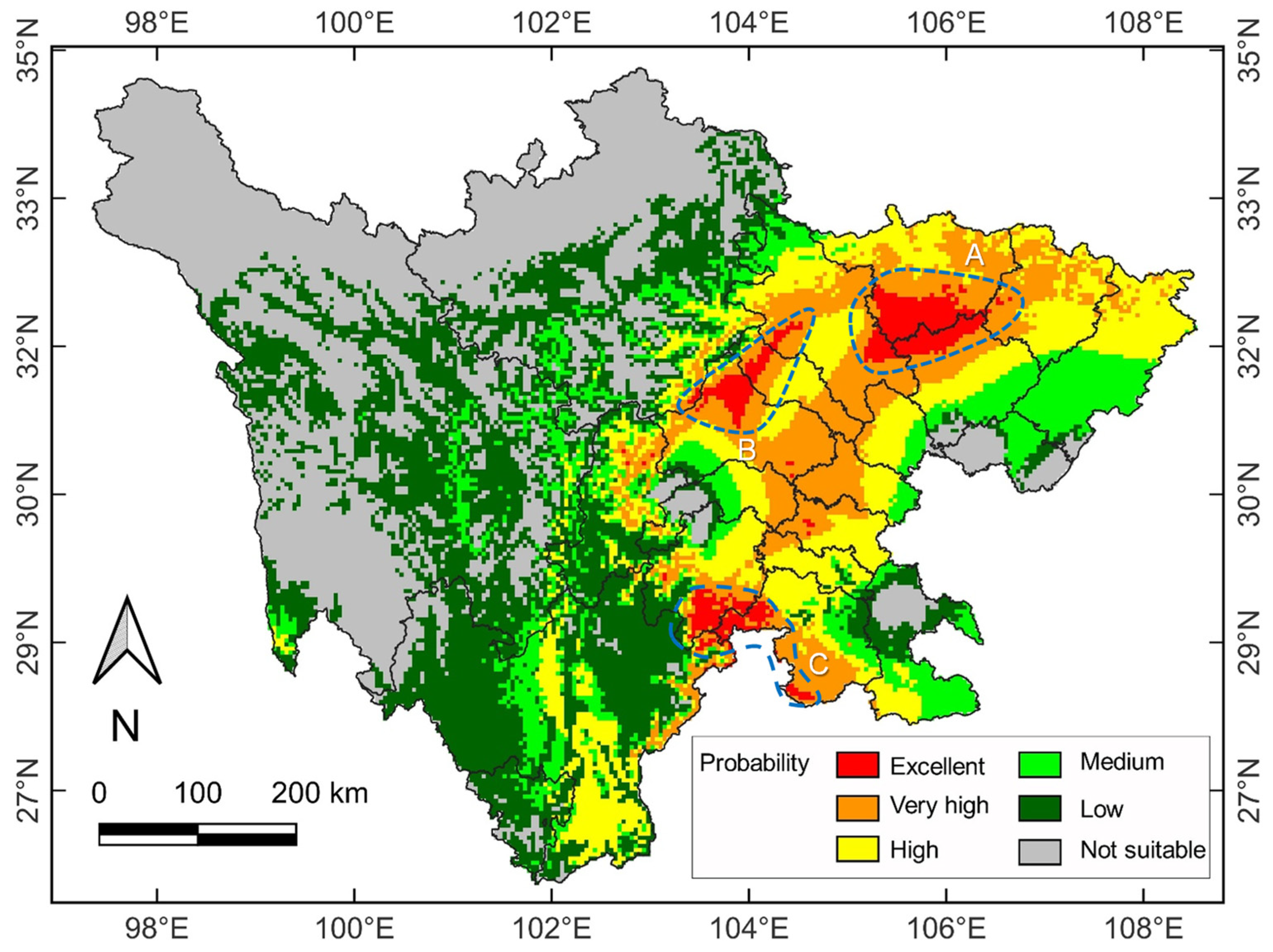
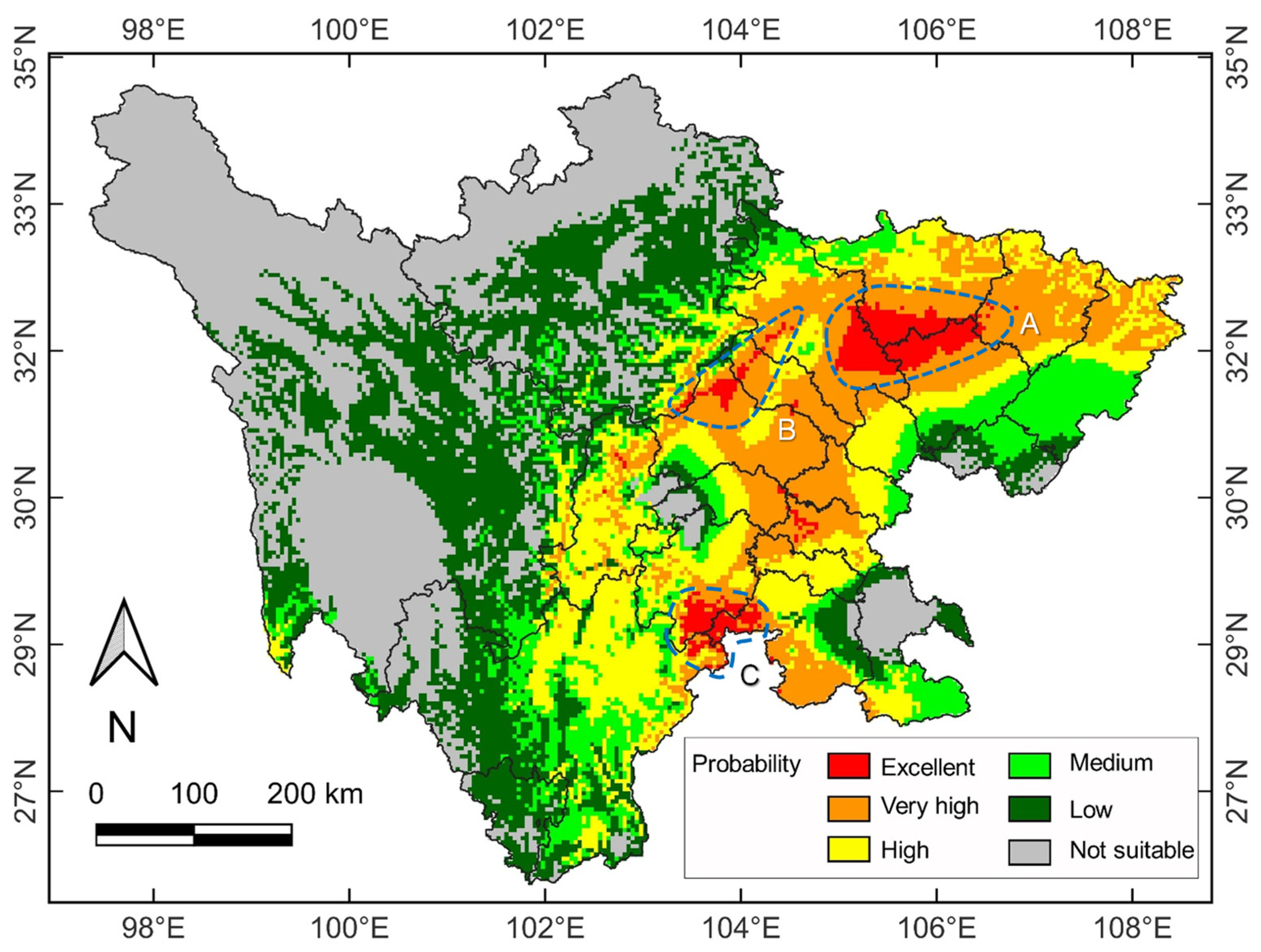
2.4. Current and Future Distribution Patterns
3. Discussion
3.1. Characteristics of Venerable Trees Resources in Sichuan
3.2. Elevation Effect on Distribution Patterns
3.3. Current Potential Suitability Habitats and Key Environmental Variables
3.4. Spatial Distribution under Future Climate Change Scenario
4. Materials and Methods
4.1. Study Area
4.2. Collecting Venerable Tree Data
4.3. Selecting Environmental Variables
4.4. Establishing the Species Distribution Model
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jim, C.Y. Urban heritage trees: Natural-cultural significance informing management and conservation. In Greening Cities: Forms and Functions; Tan, P.Y., Jim, C.Y., Eds.; Springer: Singapore, 2017; pp. 279–305. [Google Scholar]
- Lindenmayer, D.B.; Laurance, W.F. The ecology, distribution, conservation and management of large old trees. Biol. Rev. 2017, 92, 1434–1458. [Google Scholar] [CrossRef] [PubMed]
- Taxel, I. Towards an integration of historical trees into the Mediterranean archaeological record: Case studies from Central Israel. Environ. Archaeol. 2021, 1–24. [Google Scholar] [CrossRef]
- Stagoll, K.; Lindenmayer, D.B.; Knight, E.; Fischer, J.; Manning, A.D. Large trees are keystone structures in urban parks. Conserv. Lett. 2012, 5, 115–122. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Laurance, W.F. The unique challenges of conserving large old trees. Trends Ecol. Evol. 2016, 31, 416–418. [Google Scholar] [CrossRef]
- Stahle, D.W.; Edmondson, J.R.; Howard, I.M.; Robbins, C.R.; Griffin, R.D.; Carl, A.; Hall, C.B.; Stahle, D.K.; Torbenson, M.C.A. Longevity, climate sensitivity, and conservation status of wetland trees at Black River, North Carolina. Environ. Res. Commun. 2019, 1, 041002. [Google Scholar] [CrossRef]
- Blicharska, M.; Mikusiński, G. Incorporating social and cultural significance of large old trees in conservation policy. Conserv. Biol. 2014, 28, 1558–1567. [Google Scholar] [CrossRef]
- Blicharska, M.; Mikusiński, G. Old trees: Cultural value. Science 2013, 339, 904. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Laurance, W.F.; Franklin, J.F. Global decline in large old trees. Science 2012, 338, 1305–1306. [Google Scholar] [CrossRef]
- Jones, G.M.; Keane, J.J.; Gutiérrez, R.J.; Peery, M.Z. Declining old-forest species as a legacy of large trees lost. Divers. Distrib. 2018, 24, 341–351. [Google Scholar] [CrossRef] [Green Version]
- Le Roux, D.S.; Ikin, K.; Lindenmayer, D.B.; Manning, A.D.; Gibbons, P. The future of large old trees in urban landscapes. PLoS ONE 2014, 9, e99403. [Google Scholar] [CrossRef]
- Chen, S.; Li, Z.; Zhang, M. Progress in national and international research on old trees. J. Chin. Urban For. 2022, 20, 147–152. [Google Scholar]
- Zhang, H.; Lai, P.Y.; Jim, C.Y. Species diversity and spatial pattern of old and precious trees in Macau. Landsc. Urban Plann. 2017, 162, 56–67. [Google Scholar] [CrossRef]
- Jim, C.Y.; Zhang, H. Species diversity and spatial differentiation of old-valuable trees in urban Hong Kong. Urban Urban Gree 2013, 12, 171–182. [Google Scholar] [CrossRef]
- Hartel, T.; Hanspach, J.; Moga, C.I.; Holban, L.; Szapanyos, Á.; Tamás, R.; Hováth, C.; Réti, K.-O. Abundance of large old trees in wood-pastures of Transylvania (Romania). Sci. Total Environ. 2018, 613, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Lindenmayer, D.B.; Yang, W.; Ren, Y.; Campbell, M.J.; Wu, C.; Luo, Y.; Zhong, L.; Yu, M. Diversity and density patterns of large old trees in China. Sci. Total Environ. 2019, 655, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Jin, C.; Zhen, M.; Zhou, L.; Qian, S.; Jim, C.Y.; Lin, D.; Zhao, L.; Minor, J.; Coggins, C.; et al. Biogeographic and anthropogenic factors shaping the distribution and species assemblage of heritage trees in China. Urban Urban Gree 2020, 50, 126652. [Google Scholar] [CrossRef]
- Hansen, A.J.; Neilson, R.P.; Dale, V.H.; Flather, C.H.; Iverson, L.R.; Currie, D.J.; Shafer, S.; Cook, R.; Bartlein, P.J. Global change in forests: Responses of species, communities, and biomes: Interactions between climate change and land use are projected to cause large shifts in biodiversity. Bioscience 2001, 51, 765–779. [Google Scholar] [CrossRef]
- Albrich, K.; Rammer, W.; Seidl, R. Climate change causes critical transitions and irreversible alterations of mountain forests. Glob. Chang. Biol. 2020, 26, 4013–4027. [Google Scholar] [CrossRef]
- Zhao, L.; Yang, K.; Luo, Y. The temporal and spatial distribution of Chinese old trees and its analysis. IOP Conf. Ser. Earth Environ. Sci. 2021, 658, 012010. [Google Scholar] [CrossRef]
- Xie, C.; Yu, X.; Liu, D.; Fang, Y. Modelling suitable habitat and ecological characteristics of old trees using DIVA-GIS in Anhui Province, China. Pol. J. Environ. Stud. 2020, 29, 1931–1943. [Google Scholar] [CrossRef]
- Huang, L.; Tian, L.; Zhou, L.; Jin, C.; Qian, S.; Jim, C.Y.; Lin, D.; Zhao, L.; Minor, J.; Coggins, C.; et al. Local cultural beliefs and practices promote conservation of large old trees in an ethnic minority region in southwestern China. Urban Urban Gree 2020, 49, 126584. [Google Scholar] [CrossRef]
- Chi, X.; Yang, G.; Sun, K.; Li, X.; Wang, T.; Zhang, A.; Li, Y.; Cheng, M.; Wang, Q. Old ginkgo trees in China: Distribution, determinants and implications for conservation. Glob. Ecol. Conserv. 2020, 24, e01304. [Google Scholar] [CrossRef]
- Dyderski, M.K.; Paź, S.; Frelich, L.E.; Jagodziński, A.M. How much does climate change threaten European forest tree species distributions? Glob. Chang. Biol. 2018, 24, 1150–1163. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, V.; Lafond, V.; Griess, V.C. Species distribution models (SDM): Applications, benefits and challenges in invasive species management. CABI Rev. 2019, 14, 1–13. [Google Scholar] [CrossRef]
- Urbina-Cardona, N.; Blair, M.E.; Londoño, M.C.; Loyola, R.; Velásquez-Tibatá, J.; Morales-Devia, H. Species distribution modeling in Latin America: A 25-year retrospective review. Trop. Conserv. Sci. 2019, 12, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Volis, S. Species Distribution Modeling—A conservation tool with a wide spectrum of applications in Central Asia. Plant Divers. Cent. Asia 2022, 1, 26–66. [Google Scholar] [CrossRef]
- Booth, T.H. Why understanding the pioneering and continuing contributions of BIOCLIM to species distribution modelling is important. Austral. Ecol. 2018, 43, 852–860. [Google Scholar] [CrossRef] [Green Version]
- Vaz, U.; Cunha, H.; Nabout, J. Trends and biases in global scientific literature about ecological niche models. Braz. J. Biol. 2015, 75, 17–24. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Shen, J. Existing problems in and suggestions for the conservation of ancient and famous trees in Sichuan Province. Anhui For. Sci. Technol. 2020, 46, 50–52. [Google Scholar] [CrossRef]
- Munné-Bosch, S. Limits to tree growth and longevity. Trends Plant Sci. 2018, 23, 985–993. [Google Scholar] [CrossRef]
- Liu, J.; Yang, B.; Lindenmayer, D.B. The oldest trees in China and where to find them. Front. Ecol. Environ. 2019, 17, 319–322. [Google Scholar] [CrossRef]
- Chen, C.; Xia, X.; Peng, J.; Wang, D. Comparative analyses of six complete chloroplast genomes from the genus Cupressus and Juniperus (Cupressaceae). Gene 2022, 837, 146696. [Google Scholar] [CrossRef] [PubMed]
- Feng, G. Review and progress of research on plant culture. Chin. Sci. Bull. 2013, 58, 1–8. [Google Scholar] [CrossRef]
- Wang, R.; Guo, Y.; Lin, H.; Liu, Z.; Zhang, R.; Zhou, Z. Research progress and prospect of Cupressus funebris. Hubei For. Sci. Technol. 2021, 50, 60–63. [Google Scholar] [CrossRef]
- Wu, B.; Qi, S. Effects of underlay on hill-slope surface runoff process of Cupressus funebris Endl. plantations in southwestern China. Forests 2021, 12, 644. [Google Scholar] [CrossRef]
- Colman, P. Corpses, Coffins, and Crypts: A History of Burial; Henry Holt and Company, LLC: New York, NY, USA, 1997. [Google Scholar]
- Ding, J. On the custom about planting pines and cypresses in cemeteries in ancient China and its cultural connotation of funeral. J. Beijing For. Univ. (Soc. Sci.) 2022, 21, 39–46. [Google Scholar] [CrossRef]
- Zeng, H.; Qu, X. Analysis of ancient ceremonial plants in China. Bull. Biol. 2022, 57, 1–5. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, F. Artificial old trees in Sichuan: A preliminary study. Agric. Archaeol. 1985, 1, 187–192. [Google Scholar] [CrossRef]
- Yu, K.; Li, D.; Li, N. The evolution of greenways in China. Landsc. Urban Plann. 2006, 76, 223–239. [Google Scholar] [CrossRef]
- Fu, J.; Tang, T.; He, X.; Cheng, J.; Li, X.; Fu, J.; Yu, D. A preliminary report on investigation of ancient cypress trees in Jiange County. J. Sichuan For. Sci. Technol. 2018, 39, 97–101. [Google Scholar] [CrossRef]
- Li, R. The street trees of ancient China. Green. Life 2003, 5, 29. [Google Scholar]
- Guan, C. Culture of ginkgo worship in China. Agric. Archaeol. 2007, 1, 169–173. [Google Scholar] [CrossRef]
- Chen, F.; Fan, B. Influence of Buddhism on ginkgo culture. World For. Res. 2013, 26, 10–14. [Google Scholar] [CrossRef]
- Ding, X.; Xiao, J.; Huang, J.; Li, J. Investigation on wild resource of valuable timber tree Phoebe zhennan(Lauraceae). Plant Divers. 2015, 37, 629–639. [Google Scholar] [CrossRef]
- Wu, Y.; Lei, F. Species richness patterns and mechanisms along the elevational gradients. Chin. J. Zool. 2013, 48, 797–807. [Google Scholar] [CrossRef]
- Pauchard, A.; Kueffer, C.; Dietz, H.; Daehler, C.C.; Alexander, J.; Edwards, P.J.; Arévalo, J.R.; Cavieres, L.A.; Guisan, A.; Haider, S.; et al. Ain’t no mountain high enough: Plant invasions reaching new elevations. Front. Ecol. Environ. 2009, 7, 479–486. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Huang, Q. A study of the relationship between population migration and forest vicissitude: A case study of Tongjiang County. J. Yangtze Norm. Univ. 2015, 31, 30–34. [Google Scholar] [CrossRef]
- Abbas, M.; Sharma, G.; Dambire, C.; Marquez, J.; Alonso-Blanco, C.; Proaño, K.; Holdsworth, M.J. An oxygen-sensing mechanism for angiosperm adaptation to altitude. Nature 2022, 606, 565–569. [Google Scholar] [CrossRef]
- Shi, D. Plant diversity in Sichuan and Chengdu. J. Sichuan For. Sci. Technol. 1999, 20, 53–57. [Google Scholar]
- Chen, B. Spatial-temporal characteristics of precipitable water over Sichuan. Plateau Mt. Meteorol. Res. 2010, 30, 52–57. [Google Scholar] [CrossRef]
- Zhong, Z.; Qin, W.; Xu, M. A discussion of the geographic and historical evolution of vegetation in Sichuan. J. Southwest China Norm. Univ. (Nat. Sci. Ed. ) 1979, 4, 1–13. [Google Scholar] [CrossRef]
- Blach-Overgaard, A.; Svenning, J.-C.; Dransfield, J.; Greve, M.; Balslev, H. Determinants of palm species distributions across Africa: The relative roles of climate, non-climatic environmental factors, and spatial constraints. Ecography 2010, 33, 380–391. [Google Scholar] [CrossRef]
- Mahmoodi, S.; Heydari, M.; Ahmadi, K.; Khwarahm, N.R.; Karami, O.; Almasieh, K.; Naderi, B.; Bernard, P.; Mosavi, A. The current and future potential geographical distribution of Nepeta crispa Willd., an endemic, rare and threatened aromatic plant of Iran: Implications for ecological conservation and restoration. Ecol. Indic. 2022, 137, 108752. [Google Scholar] [CrossRef]
- González-Orozco, C.E.; Thornhill, A.H.; Knerr, N.; Laffan, S.; Miller, J.T. Biogeographical regions and phytogeography of the eucalypts. Divers. Distrib. 2014, 20, 46–58. [Google Scholar] [CrossRef]
- Asefa, M.; Cao, M.; He, Y.; Mekonnen, E.; Song, X.; Yang, J. Ethiopian vegetation types, climate and topography. Plant Diversity 2020, 42, 302–311. [Google Scholar] [CrossRef]
- Yilmaz, H.Ç.; Yilmaz, O.Y.; Yilmaz, H. Bioclimatic factors affecting the distribution of Quercus robur L.(pedunculate oak) subspecies in Turkey. Eurasian J. For. Sci. 2016, 4, 31–39. [Google Scholar] [CrossRef] [Green Version]
- Litton, C.M.; Giardina, C.P.; Freeman, K.R.; Selmants, P.C.; Sparks, J.P. Impact of mean annual temperature on nutrient availability in a tropical montane wet forest. Front. Plant Sci. 2020, 11, 784. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; An, M.; Wu, J.; Liu, F.; Wang, W. Geographical distribution pattern and dominant climatic factors of the Paphiopedilum Subgen. Brachypetalum in China. Chin. J. Plant Ecol. 2022, 46, 40–50. [Google Scholar] [CrossRef]
- Liu, W.; You, J.; Zeng, W.; Qi, D. Prediction of the geographical distribution of Carex moorcroftii under global climate change based on MaxEnt model. Chin. J. Grassl. 2018, 40, 43–49. [Google Scholar] [CrossRef]
- Peer, L.A.; Dar, Z.A.; Lone, A.A.; Bhat, M.Y.; Ahamad, N. High temperature triggered plant responses from whole plant to cellular level. Plant Physiol. Rep. 2020, 25, 611–626. [Google Scholar] [CrossRef]
- Lippmann, R.; Babben, S.; Menger, A.; Delker, C.; Quint, M. Development of wild and cultivated plants under global warming conditions. Curr. Biol. 2019, 29, R1326–R1338. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P. Assessment of impact of climate change on Rhododendrons in Sikkim Himalayas using MaxEnt modelling: Limitations and challenges. Biodivers. Conserv. 2012, 21, 1251–1266. [Google Scholar] [CrossRef]
- Morueta-Holme, N.; Engemann, K.; Sandoval-Acuña, P.; Jonas, J.D.; Segnitz, R.M.; Svenning, J.-C. Strong upslope shifts in Chimborazo’s vegetation over two centuries since Humboldt. Proc. Natl. Acad. Sci. USA 2015, 112, 12741–12745. [Google Scholar] [CrossRef] [PubMed]
- Colwell, R.K.; Brehm, G.; Cardelús, C.L.; Gilman, A.C.; Longino, J.T. Global warming, elevational range shifts, and lowland biotic attrition in the wet tropics. Science 2008, 322, 258–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prevéy, J.S.; Parker, L.E.; Harrington, C.A.; Lamb, C.T.; Proctor, M.F. Climate change shifts in habitat suitability and phenology of huckleberry (Vaccinium membranaceum). Agric. For. Meteorol. 2020, 280, 107803. [Google Scholar] [CrossRef]
- Abdelaal, M.; Fois, M.; Fenu, G.; Bacchetta, G. Using MaxEnt modeling to predict the potential distribution of the endemic plant Rosa arabica Crép. in Egypt. Ecol. Inform. 2019, 50, 68–75. [Google Scholar] [CrossRef]
- Lu, S.; Zhou, S.; Yin, X.; Zhang, C.; Li, R.; Chen, J.; Ma, D.; Wang, Y.; Yu, Z.; Chen, Y. Patterns of tree species richness in Southwest China. Environ. Monit. Assess. 2021, 193, 1–13. [Google Scholar] [CrossRef]
- Liu, F.; Luo, J.; Yang, M. Potential geographic distribution areas of Toona sinensis and their climatic characteristics in southwest China regions. J. Sichuan For. Sci. Technol. 2017, 38, 16–21. [Google Scholar] [CrossRef]
- Committee, S.P.G. Announcement on the List of Old and Valuable Trees in Sichuan Province; Sichuan Greening Committee: Chengdu, China, 2020.
- Lai, P.Y.; Jim, C.Y.; Tang, G.D.; Hong, W.J.; Zhang, H. Spatial differentiation of heritage trees in the rapidly-urbanizing city of Shenzhen, China. Landsc. Urban Plann. 2019, 181, 148–156. [Google Scholar] [CrossRef]
- Villalba, R.; Veblen, T.T. Improving estimates of total tree ages based on increment core samples. Ecoscience 1997, 4, 534–542. [Google Scholar] [CrossRef]
- Fortin, M.-J. Effects of sampling unit resolution on the estimation of spatial autocorrelation. Ecoscience 1999, 6, 636–641. [Google Scholar] [CrossRef]
- Hannah, L.; Aguilar, G.; Blanchon, D. Spatial distribution of the Mexican daisy, Erigeron karvinskianus, in New Zealand under climate change. Climate 2019, 7, 24. [Google Scholar] [CrossRef] [Green Version]
- Varol, T.; Canturk, U.; Cetin, M.; Ozel, H.B.; Sevik, H. Impacts of climate change scenarios on European ash tree (Fraxinus excelsior L.) in Turkey. For. Ecol. Manag. 2021, 491, 119199. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Guarino, L.; Cruz, M.; Rojas, E. Computer tools for spatial analysis of plant genetic resources data: 1. DIVA-GIS. Plant Genet. Resour. Newsl. 2001, 127, 15–19. [Google Scholar]
- Sharma, S.; Arunachalam, K.; Bhavsar, D.; Kala, R. Modeling habitat suitability of Perilla frutescens with MaxEnt in Uttarakhand—A conservation approach. J. Appl. Res. Med. Aromat. Plants 2018, 10, 99–105. [Google Scholar] [CrossRef]
- Govindasamy, B.; Duffy, P.B.; Coquard, J. High-resolution simulations of global climate, part 2: Effects of increased greenhouse cases. Clim. Dyn. 2003, 21, 391–404. [Google Scholar] [CrossRef]
- Liu, G.; Mai, J. Habitat shifts of Jatropha curcas L. in the Asia-Pacific region under climate change scenarios. Energy 2022, 251, 123885. [Google Scholar] [CrossRef]
- QGIS. QGIS Geographic Information System Version 3.20; Open Source Geospatial Foundation Project; QGIS Association: Uster, Switzerland, 2021. [Google Scholar]
- Welch, J.M. Street and park trees of Boston: A comparison of urban forest structure. Landsc. Urban Plann. 1994, 29, 131–143. [Google Scholar] [CrossRef]
- Ter Braak, C.J.; Smilauer, P. CANOCO Reference Manual and CanoDraw for Windows User’s Guide: Software for Canonical Community Ordination (Version 4.5). 2002. Available online: www.canoco.com (accessed on 15 August 2017).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bioclimatic Factor | Explained Variation % | Contribution % | F | p |
|---|---|---|---|---|
| bio7 | 34.0 | 38.3 | 113 | 0.002 |
| bio3 | 33.5 | 37.8 | 54.9 | 0.002 |
| bio19 | 9.2 | 10.4 | 42.0 | 0.002 |
| bio15 | 6.7 | 7.6 | 43.0 | 0.002 |
| bio16 | 2.6 | 3.0 | 19.8 | 0.002 |
| bio11 | 2.2 | 2.5 | 19.5 | 0.002 |
| bio18 | 0.3 | 0.4 | 3.1 | 0.066 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, C.; Li, M.; Jim, C.Y.; Liu, D. Environmental Factors Driving the Spatial Distribution Pattern of Venerable Trees in Sichuan Province, China. Plants 2022, 11, 3581. https://doi.org/10.3390/plants11243581
Xie C, Li M, Jim CY, Liu D. Environmental Factors Driving the Spatial Distribution Pattern of Venerable Trees in Sichuan Province, China. Plants. 2022; 11(24):3581. https://doi.org/10.3390/plants11243581
Chicago/Turabian StyleXie, Chunping, Meng Li, C. Y. Jim, and Dawei Liu. 2022. "Environmental Factors Driving the Spatial Distribution Pattern of Venerable Trees in Sichuan Province, China" Plants 11, no. 24: 3581. https://doi.org/10.3390/plants11243581