
Dynamics Changes in Basal Area Increment, Carbon Isotopes Composition and Water Use Efficiency in Pine as Response to Water and Heat Stress in Silesia, Poland

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
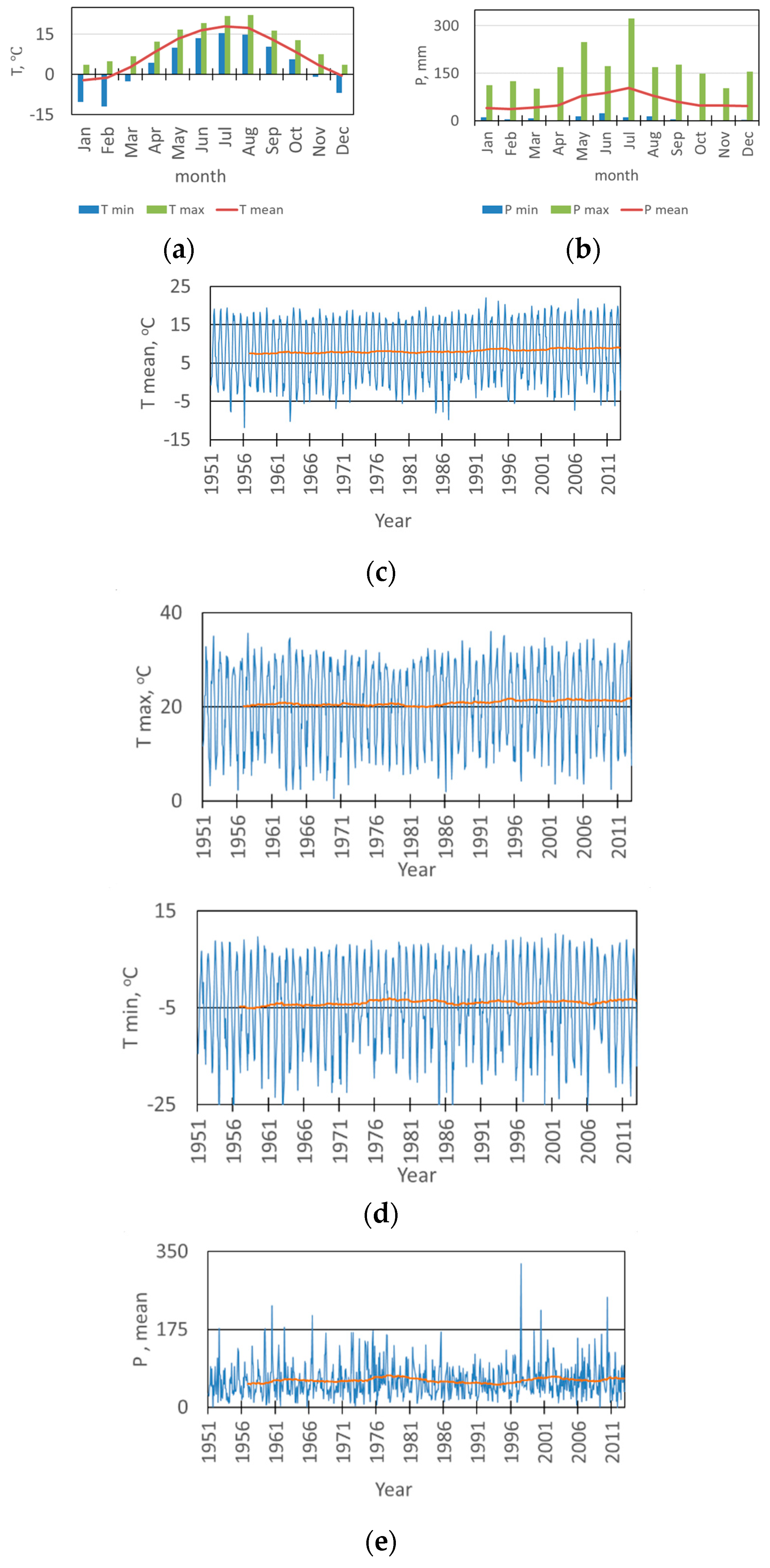
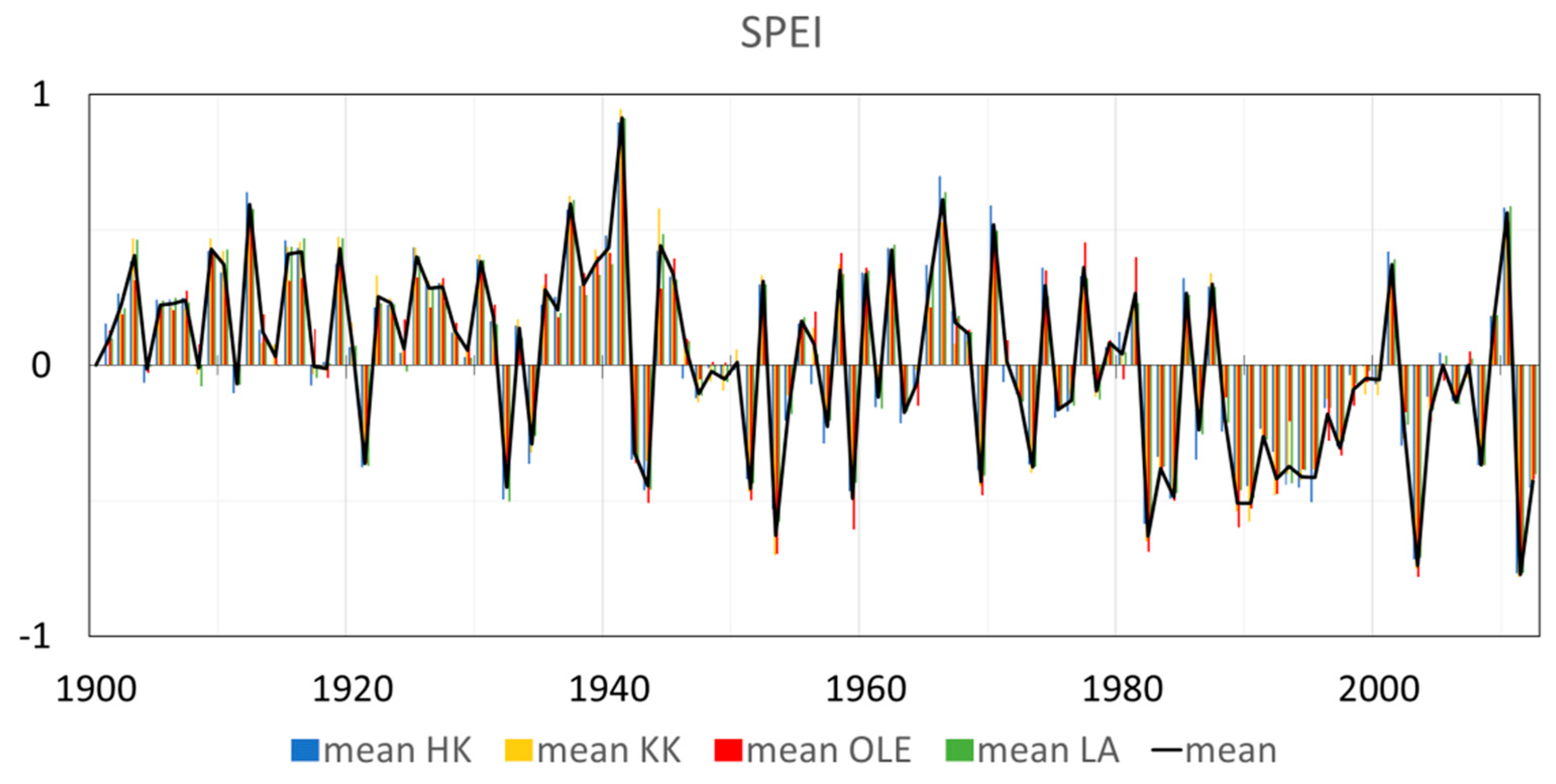
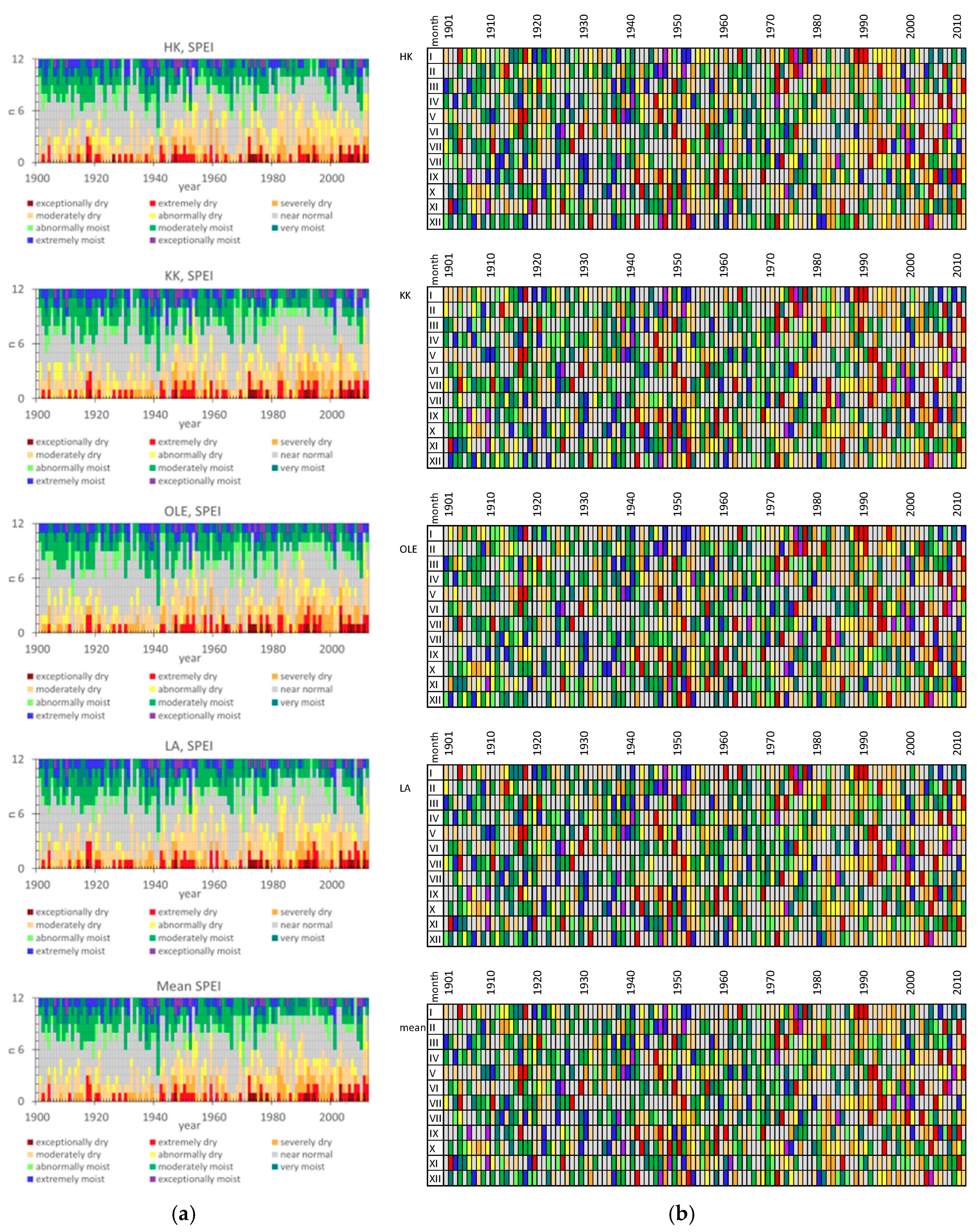
2. Results
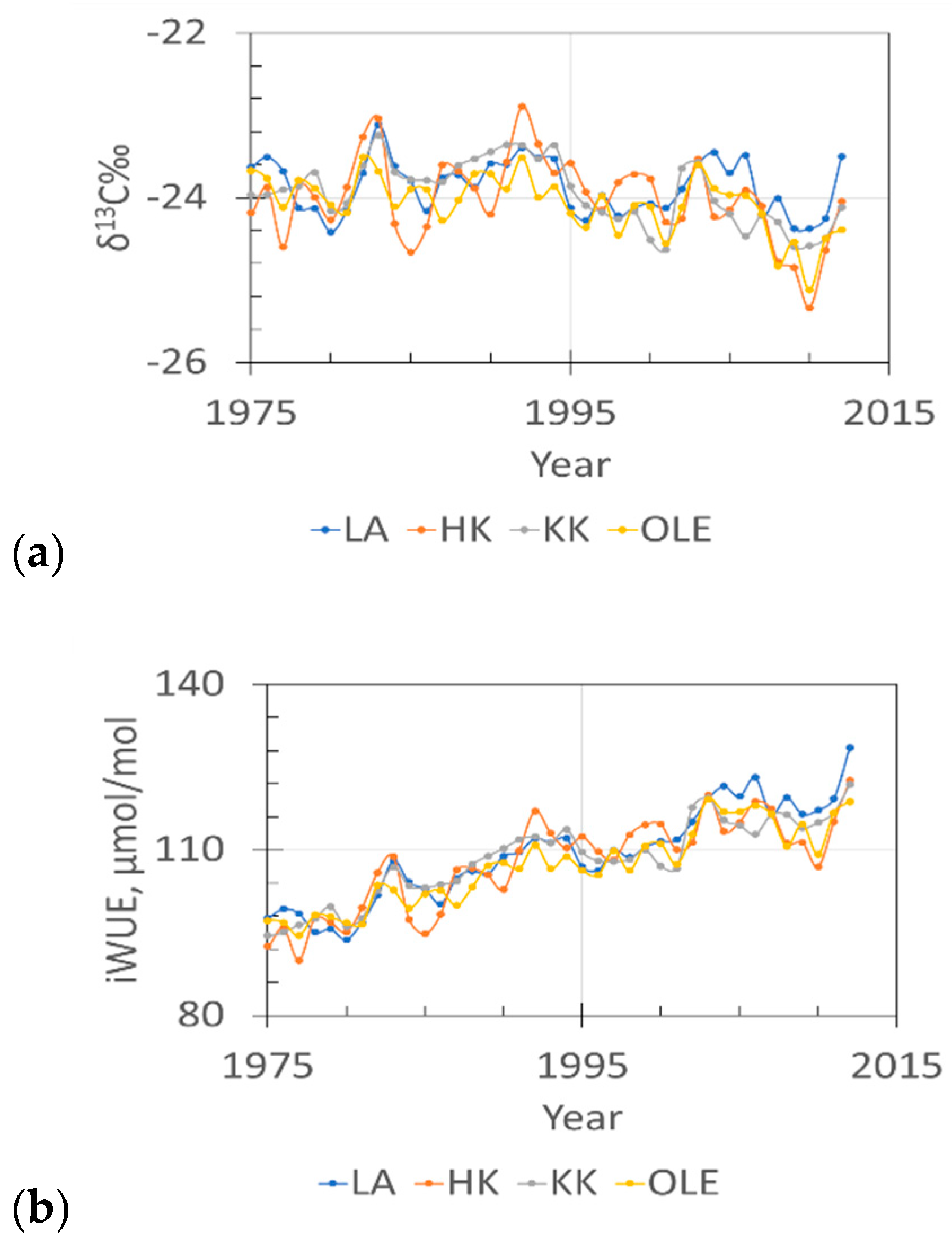
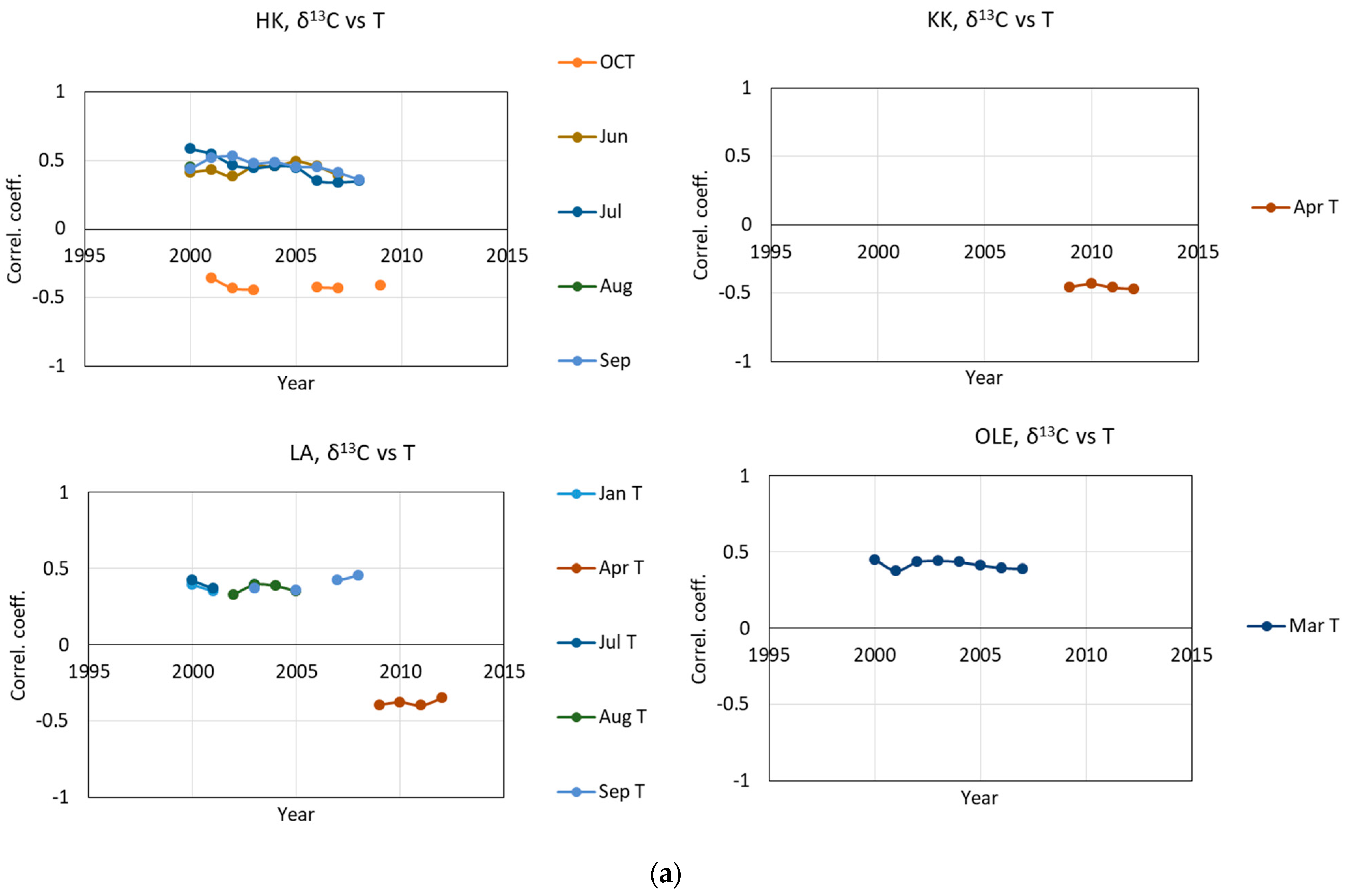
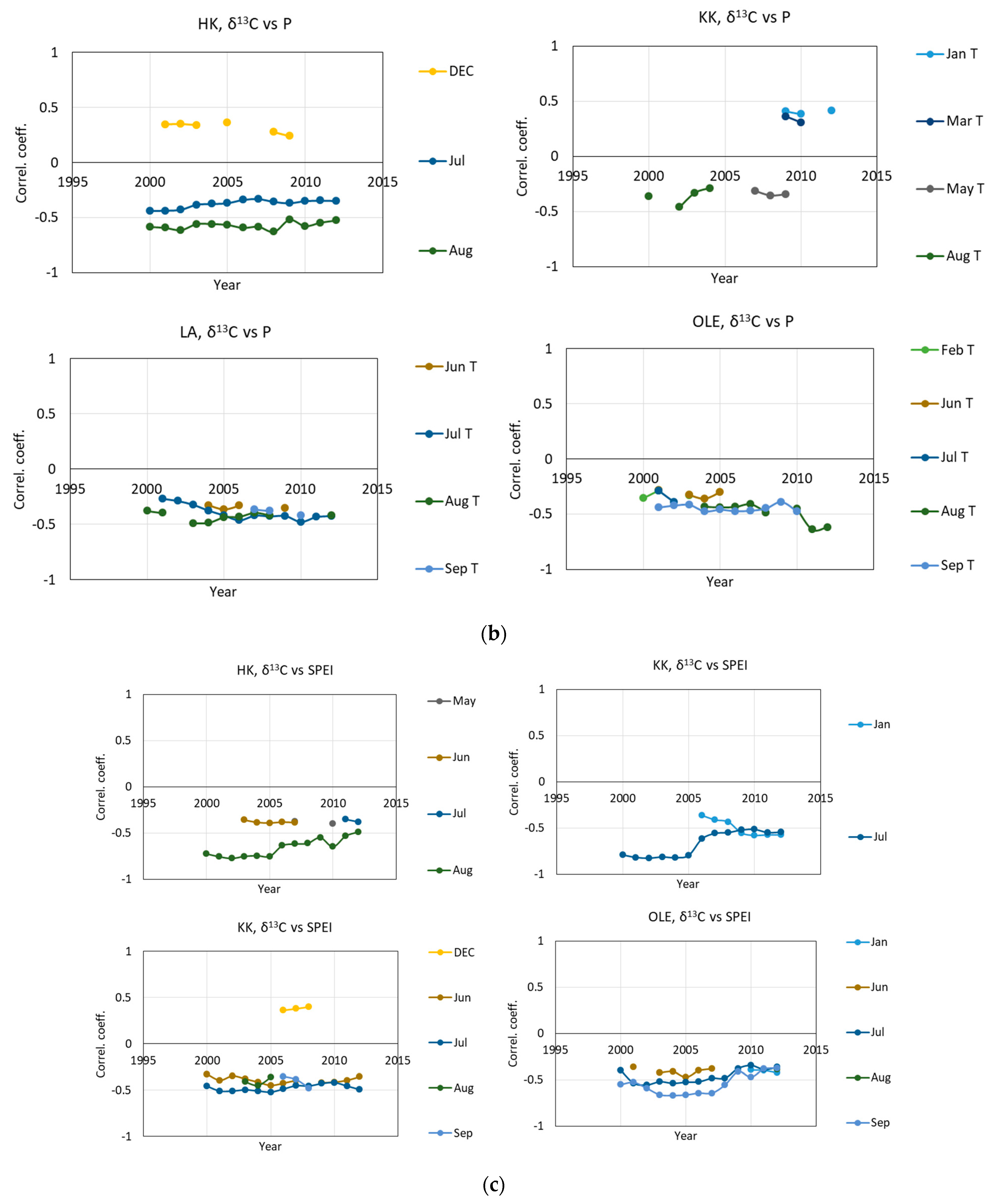
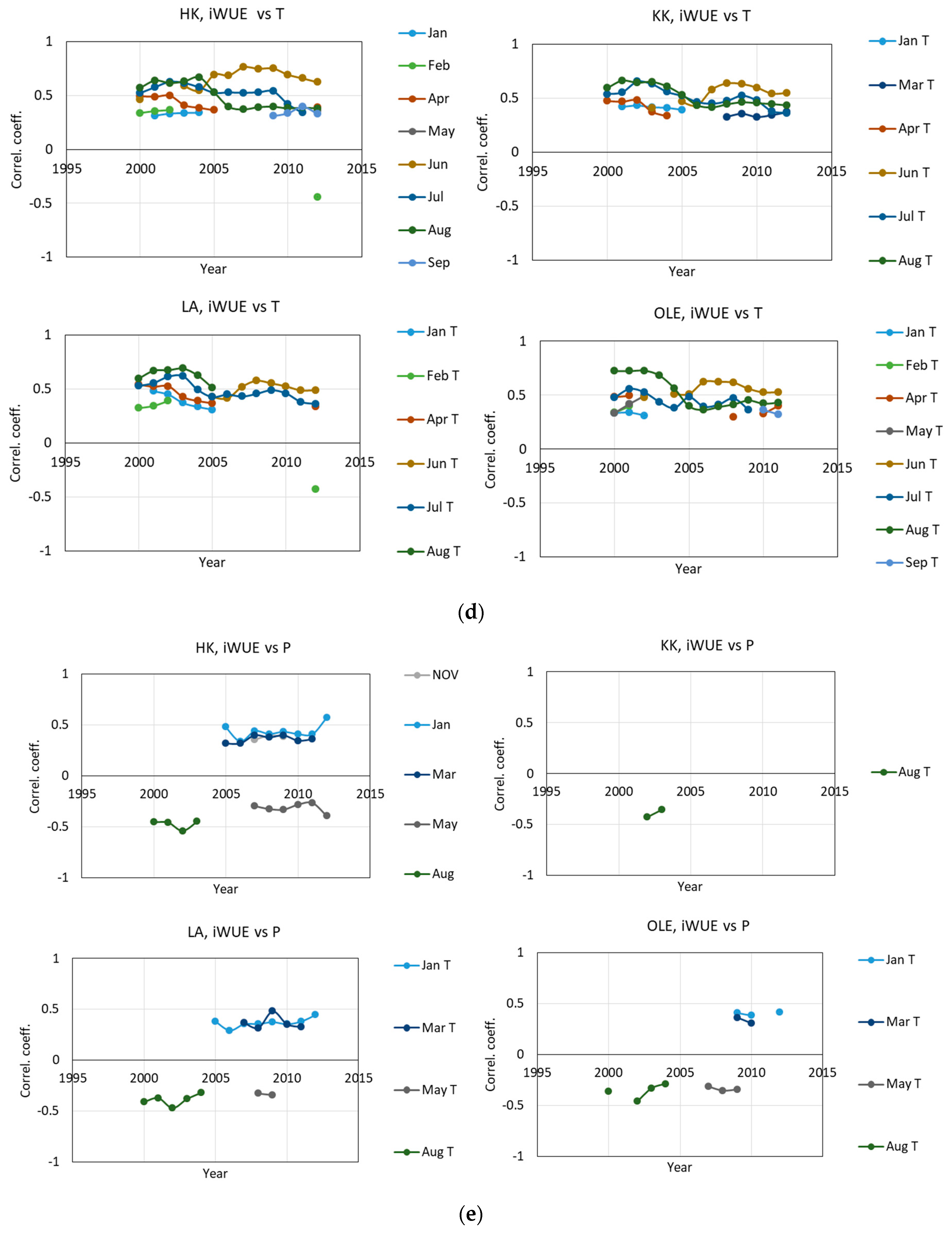
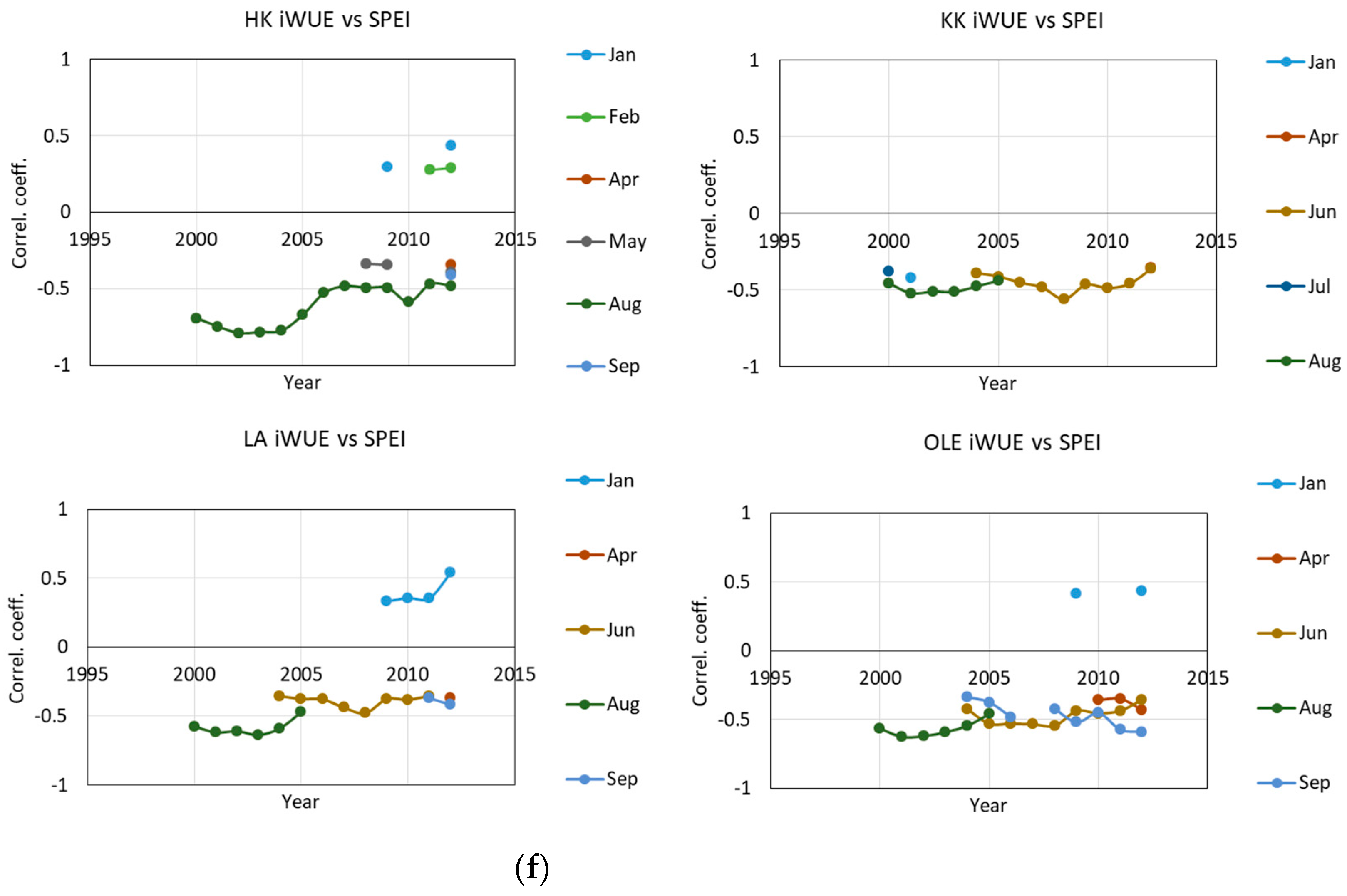
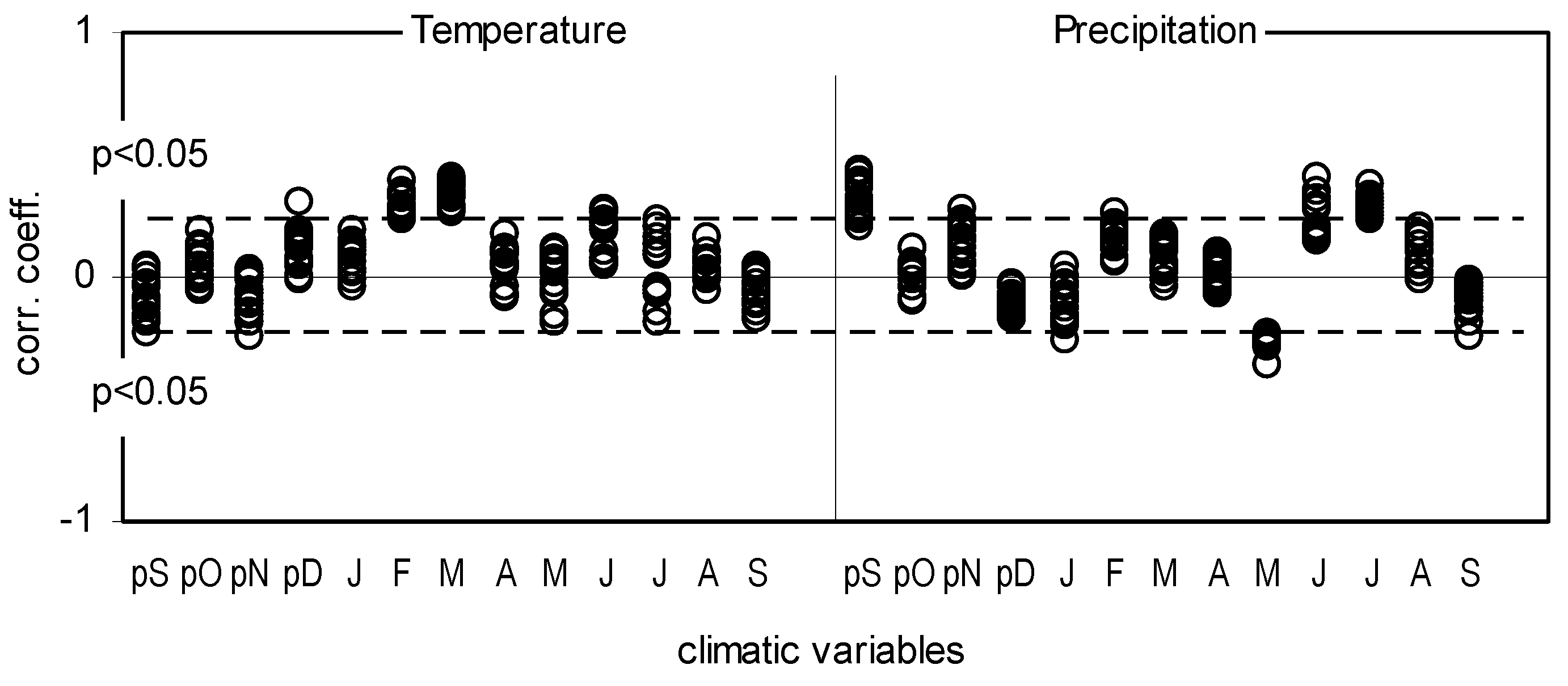
2.1. δ13 C and iWUE and Climate Changes
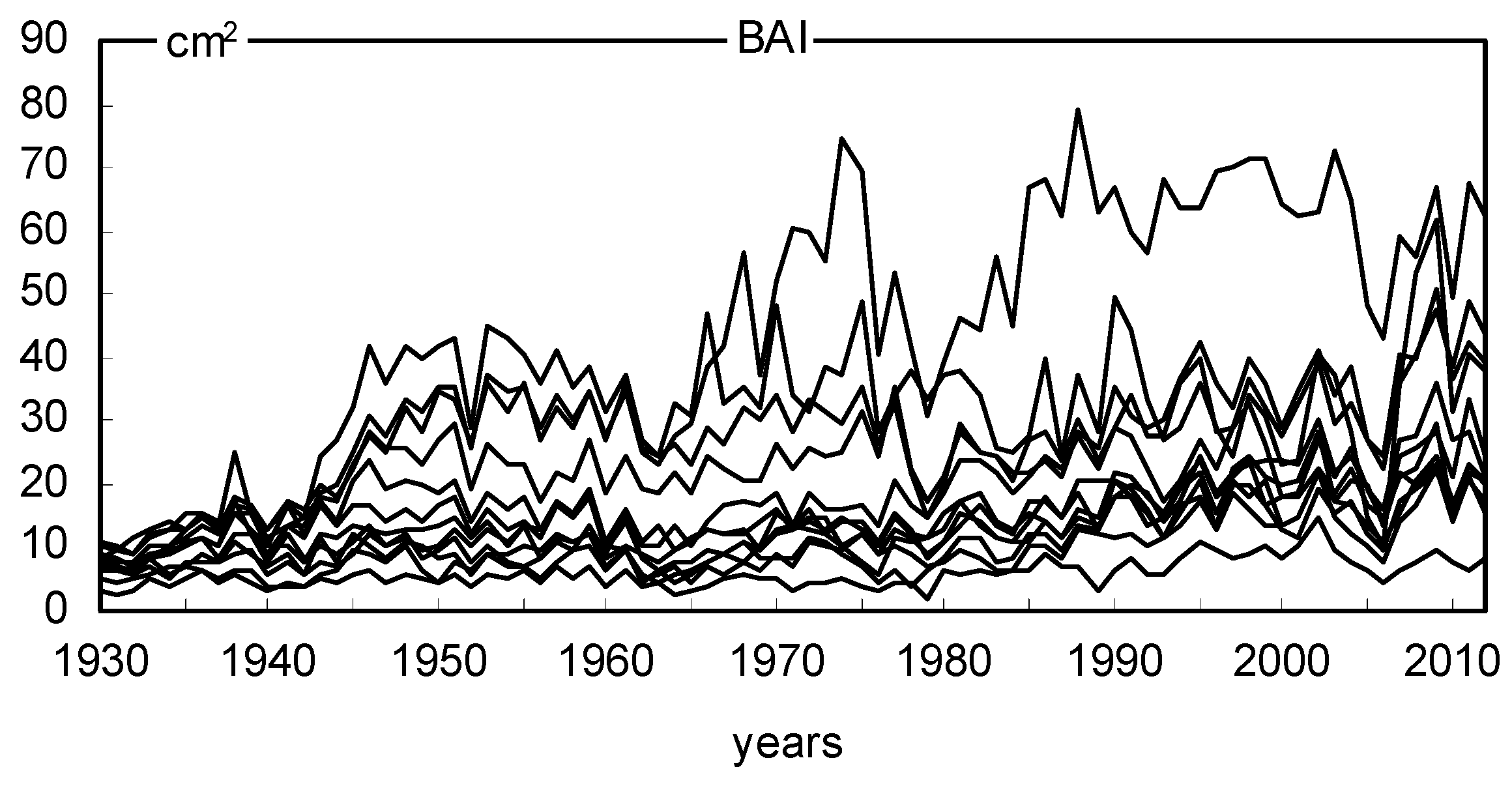
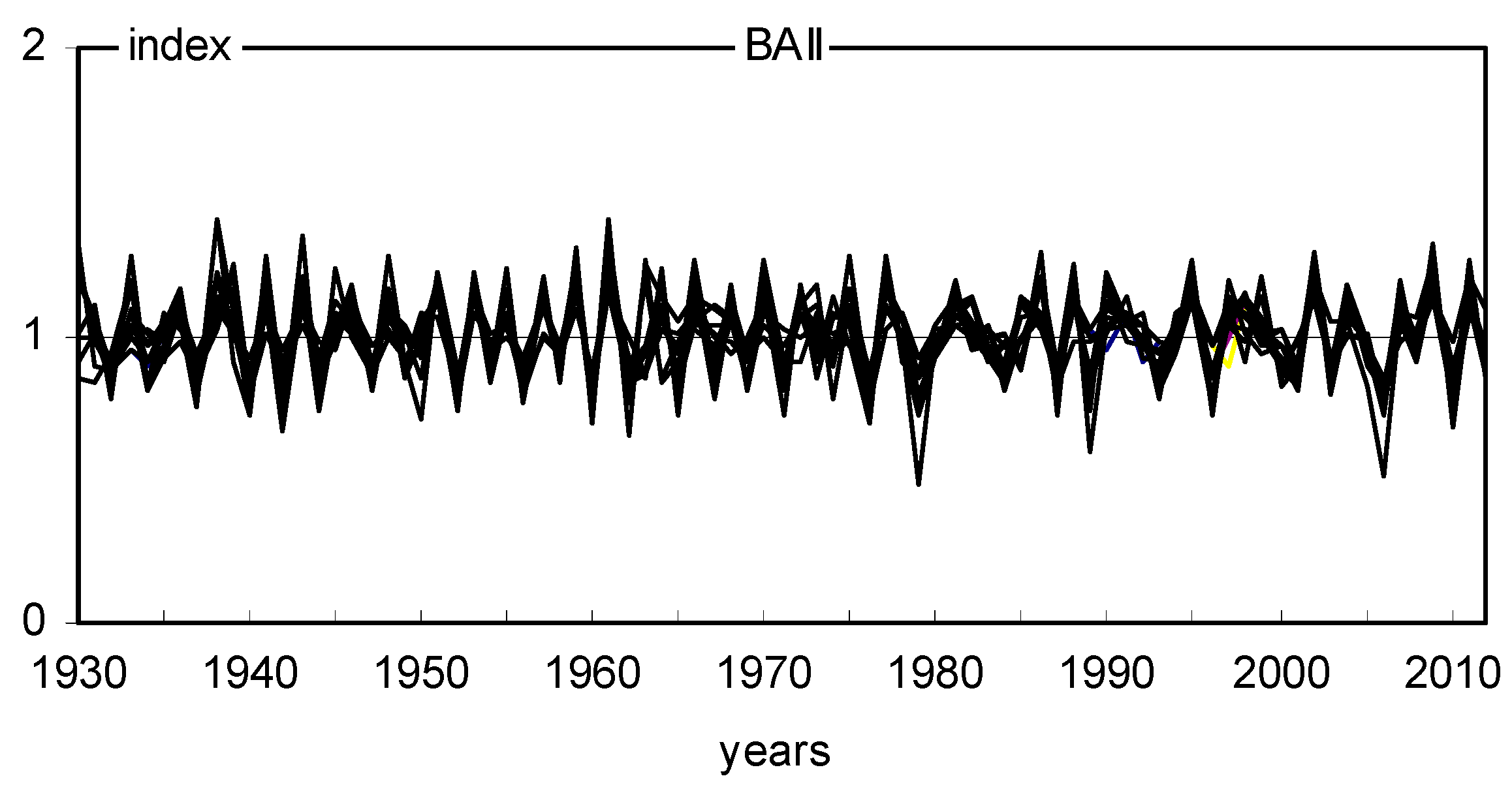
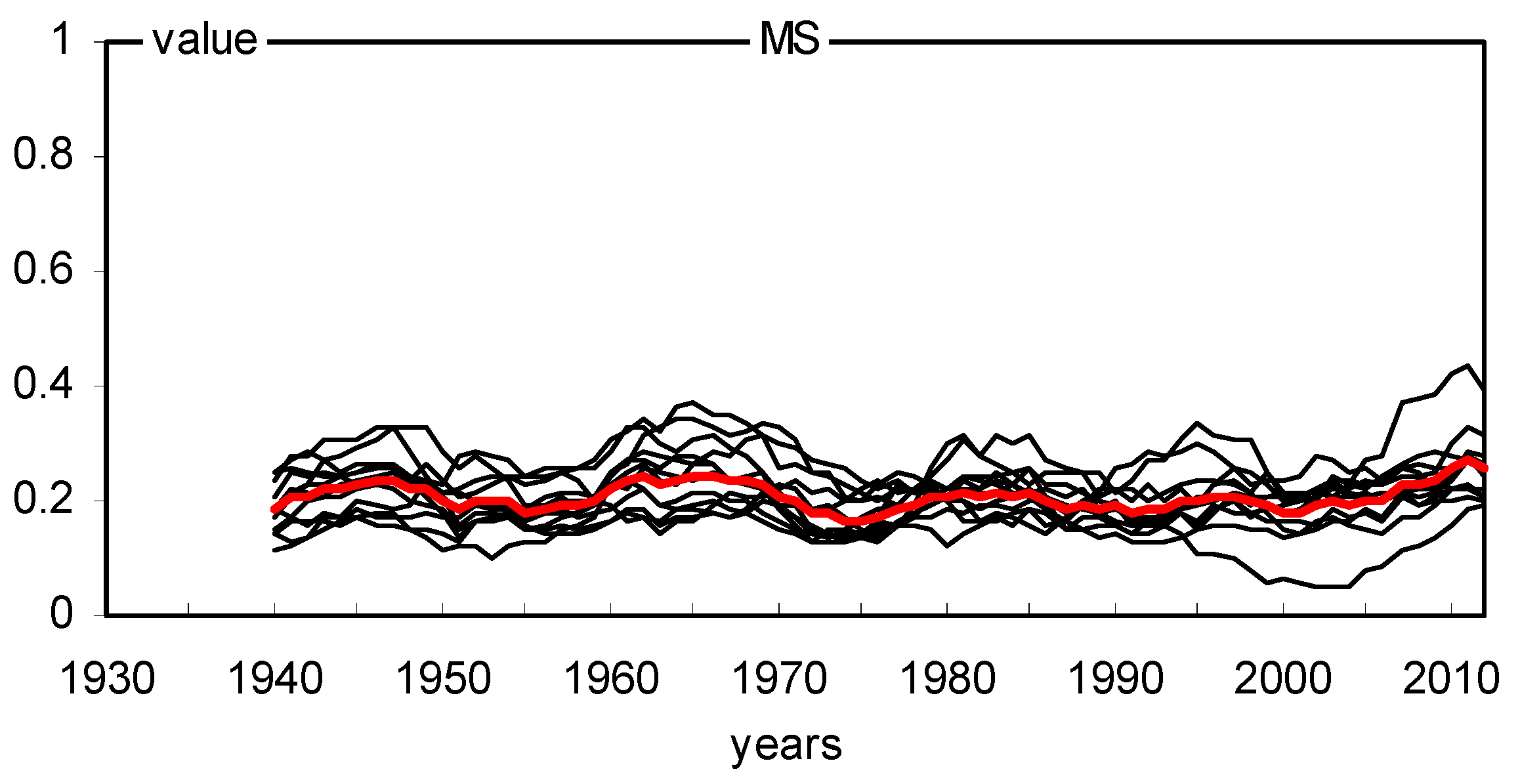
2.2. Long- and Short-Term Basal Area Increment
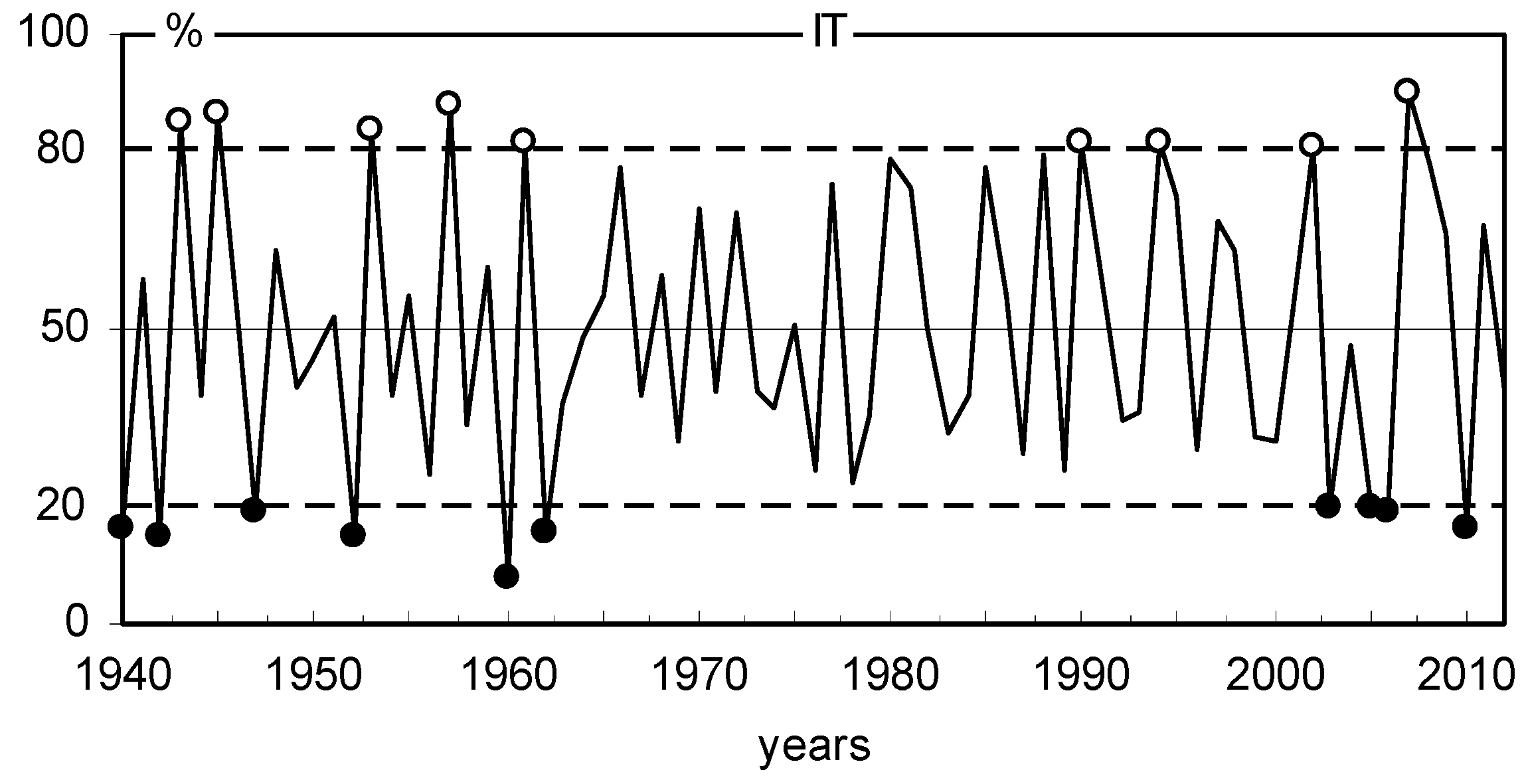
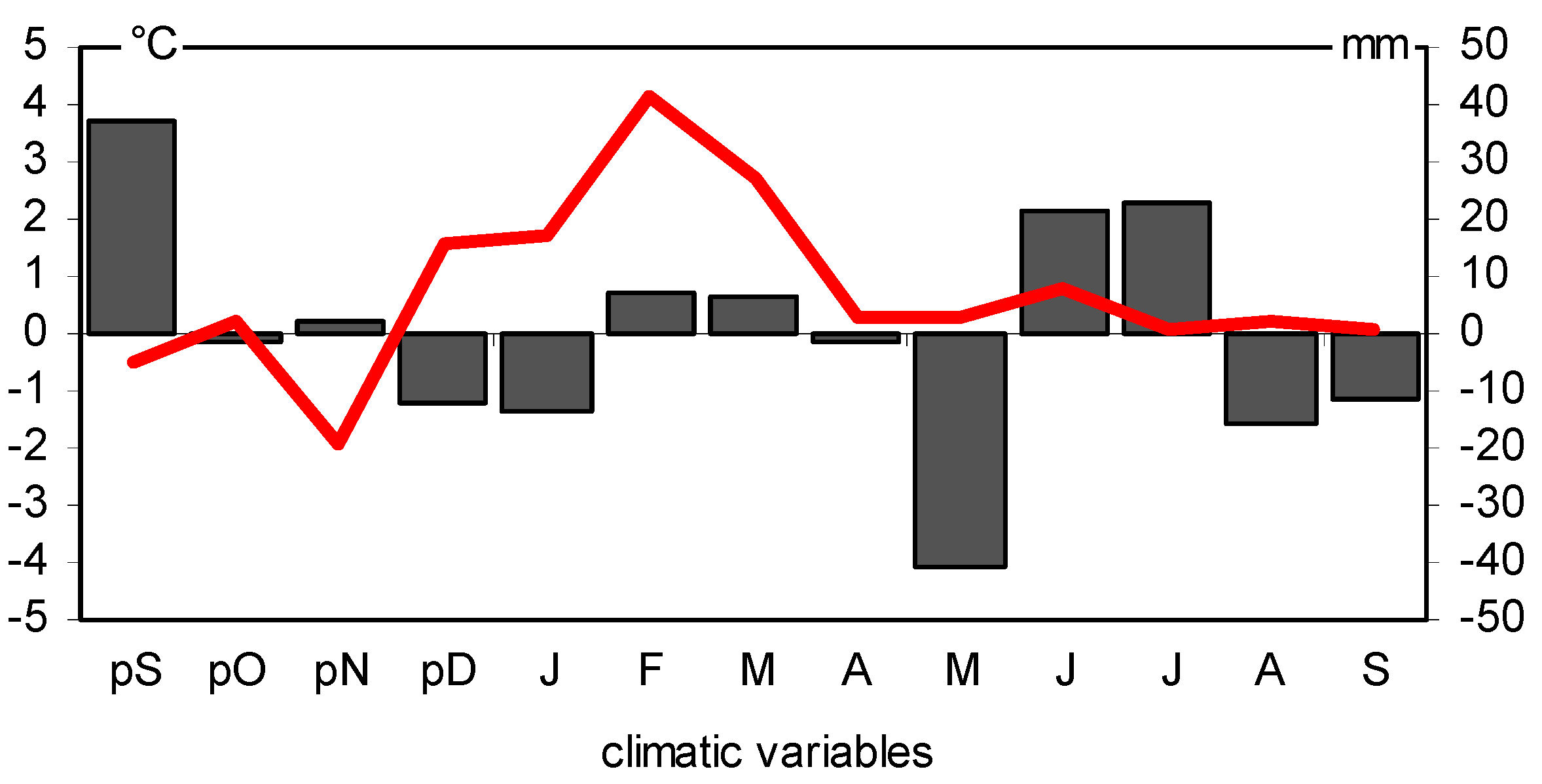
2.3. BAI, Climate Conditions, and Pointer Years
3. Discussion
3.1. Climatic Factors That Influence the Basal Area Increment
3.2. The Impact of Climatic Changes on δ13C, iWUE
- The photosynthesis rate increases significantly and the stomata conductance is at the same level or does not significantly
- The photosynthesis rate is constant, and the stomata conductance is reduced.
- The photosynthesis rate decreases and the stomata conductance is reduced, but stomata conductance reduction is a much more significant process than a reduction in photosynthesis speed.
4. Materials and Methods
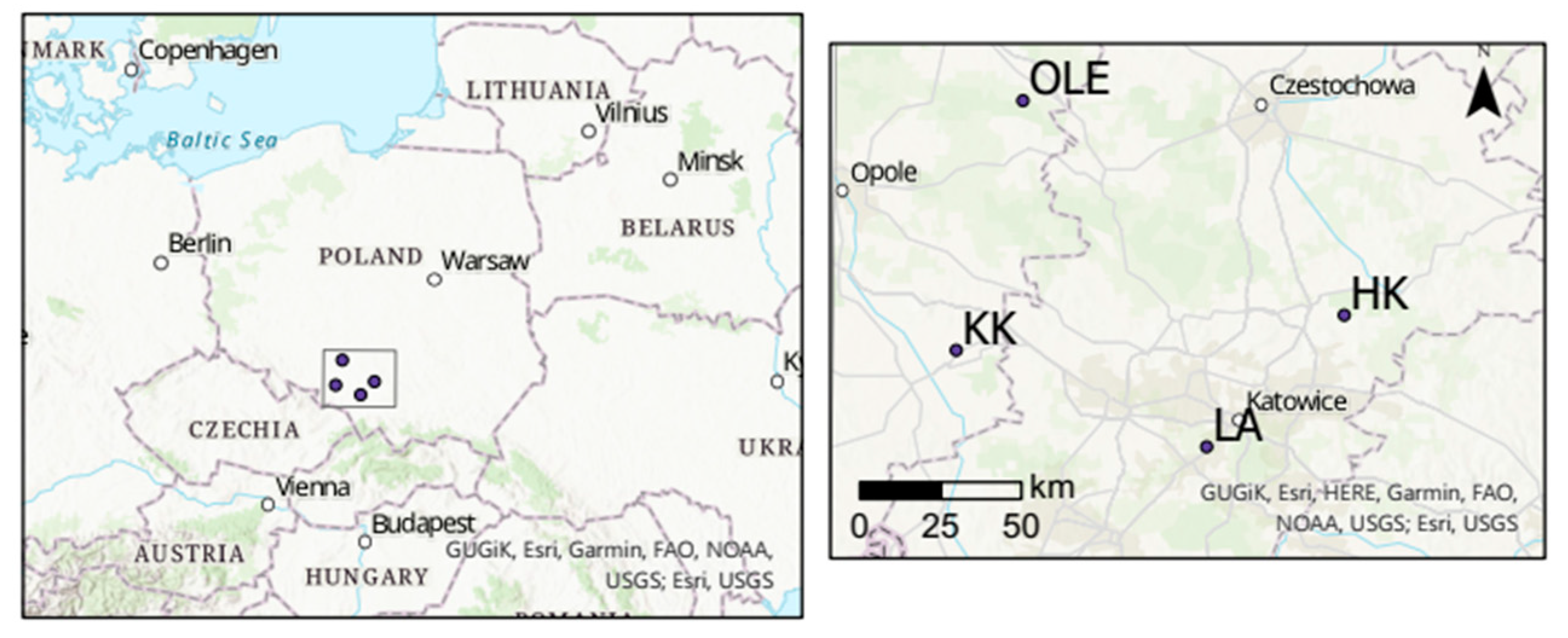
4.1. Study Area
4.2. Dendrochronological Analysis
4.3. Isotopic Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Savard, M.M. Tree-ring stable isotopes and historical perspectives on pollution: An overview. Environ. Pollut. 2010, 158, 2007–2013. [Google Scholar] [CrossRef] [PubMed]
- Ferrio, J.; Voltas, J.; Araus, J. Use of carbon isotope composition in monitoring environmental changes. Manag. Environ. Qual. 2003, 14, 82–98. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Lloyd, L. Carbon and oxygen isotope effects in the exchange of carbon dioxide between plants and the atmosphere. In Stable Isotopes and Plant Carbon-Water Relations; Ehrelinger, J.R., Hall, A.E., Farquhar, G.D., Eds.; Academic Press: New York, NY, USA, 1993; pp. 47–70. [Google Scholar]
- Malik, I.; Danek, M.; Marchwińska-Wyrwał, E.; Danek, T.; Wistuba, M.; Krąpiec, M. Scots pine (Pinus sylvestris L.) growth suppression and adverse effects on human health due to air pollution in the Upper Silesian Industrial District (USID), Southern Poland. Water Air Soil Pollut. 2012, 223, 3345–3364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dmuchowski, W.; Bytnerowicz, A. Monitoring environmental pollution in Poland by chemical analysis of Scots pine (Pinus sylvestris L.) needles. Environ. Pollut. 1995, 87, 87–104. [Google Scholar] [CrossRef]
- Białobok, S. Outline of Physiology of Scots Pine (Zarys Fizjologii Sosny Zwyczajnej), 1st ed.; SORUS: Poznań, Poland; Kórnik, Poland, 1976; pp. 1–613. [Google Scholar]
- Sensuła, B.; Wilczyński, S.; Monin, L.; Allan, M.; Pazdur, A.; Fagel, N. Variations of tree ring width and chemical composition of wood of pine growing in the area nearby chemical factories. Geochronometria 2017, 44, 226–239. [Google Scholar] [CrossRef] [Green Version]
- Sensuła, B.; Fagel, N.; Michczyński, A. Radiocarbon, trace elements and Pb isotope composition of pine needles from a highly industrialized region in southern Poland. Radiocarbon 2021, 63, 713–726. [Google Scholar] [CrossRef]
- Ehrelinger, J.R.; Hall, A.E.; Farquhar, G.D. Stable Isotopes and Plant Carbon-Water Relation, 1st ed.; Academic Press: New York, NY, USA, 1993; pp. 1–555. [Google Scholar]
- Leavitt, S.; Long, A. Stable carbon isotopes as a potential supplemental tool in dendrochronology. Tree Ring Bull. 1982, 42, 49–56. [Google Scholar]
- McCarroll, D.; Gagen, M.H.; Loader, N.J.; Robertson, I.; Anchukaitis, K.J.; Los, S.; Young, G.H.F.; Jalkanen, R.; Kirchhefer, A.; Waterhouse, J.S. Correction of tree ring stable carbon isotope chronologies for changes in the carbon dioxide content of the atmosphere. Geochim. Cosmochim. Acta 2009, 73, 1539–1547. [Google Scholar] [CrossRef]
- Craig, H. Carbon-13 in plants and the relationship between carbon-13 and carbon-14 variations in nature. J. Geol. 1954, 62, 115–149. [Google Scholar] [CrossRef]
- Keeling, C.D.; Chin, J.F.S.; Whorf, T.P. Increased activity of northern vegetation inferred from atmospheric CO2 measurements. Nature 1996, 382, 146–149. [Google Scholar] [CrossRef]
- Sensuła, B.; Michczyński, A.; Piotrowska, N.; Wilczyński, S. Anthropogenic CO2 Emission Records in Scots Pine Growing in the Most Industrialized Region of Poland from 1975 to 2014. Radiocarbon 2018, 60, 1041–1053. [Google Scholar] [CrossRef]
- Gagen, M.; Finsinger, W.; Wagner-Cremer, F.; McCarroll, D.; Loader, N.; Robertson, I.; Jalkanen, R.; Young, G.; Kirchhefer, A. Evidence of changing intrinsic water-use efficiency under rising atmospheric CO2 concentrations in Boreal Fennoscandia from sub-fossil leaves and tree ring δ13C ratios. Glob. Chang. Biol. 2011, 17, 1064–1072. [Google Scholar] [CrossRef] [Green Version]
- Scheidegger, Y.; Saurer, M.; Bahn, M.; Siegwolf, R. Linking stable oxygen and carbon isotopes with stomatal conductance and photosynthetic capacity: A conceptual model. Oecologia 2000, 125, 350–357. [Google Scholar] [CrossRef] [PubMed]
- Sensuła, B.; Wilczyński, S.; Opała, M. Tree growth and climate relationship: Dynamics of scots pine (Pinus sylvestris L.) growing in the near-source region of the combined heat and power plant during the development of the pro-ecological strategy in Poland. Water Air Soil Pollut. 2015, 226, 220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sensuła, B.; Wilczyński, S. Climatic signals in tree-ring width and stable isotopes composition of Pinus sylvestris L. Growing in the industrialized area nearby Kędzierzyn-Koźle. Geochronometria 2017, 44, 240–255. [Google Scholar] [CrossRef] [Green Version]
- Sensuła, B.; Wilczyński, S. Tree-ring widths and the stable isotope composition of pine tree-rings as climate indicators in the most industrialised part of Poland during CO2 elevation. Geochronometria 2018, 45, 130–145. [Google Scholar] [CrossRef] [Green Version]
- Sensula, B.; Opala, M.; Wilczynski, S.; Pawelczyk, S. Long- and short-term incremental response of Pinus sylvestris L. from industrial area nearby steelworks in Silesian Upland, Poland. Dendrochronologia 2015, 36, 1–12. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Beguería, S.; López-Moreno, J.I. A Multi-scalar drought index sensitive to global warming: The Standardized Precipitation Evapotranspiration Index—SPEI. J. Clim. 2010, 23, 1696–1718. [Google Scholar] [CrossRef] [Green Version]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop Evapotranspiration: Guidelines for Computing Crop Requirements; FAO Irrigation and Drainage Paper; FAO: Rome, Italy, 1998; p. 56. [Google Scholar]
- SPEI Database. Available online: https://spei.csic.es/spei_database_2_7 (accessed on 24 October 2022).
- World Bank Group GOST Climate. Available online: https://bennyistanto.github.io/gost-climate/indices/spei.html (accessed on 24 October 2022).
- Eckstein, D.; Schweingruber, F. Dendrochronologia—A mirror for 25 years of tree-ring research and a sensor for promising topics. Dendrochronologia 2009, 27, 7–13. [Google Scholar] [CrossRef]
- Hughes, M.K.; Swetnam, T.W.; Diaz, H.F. Dendroclimatology progress and prospects. In Development in Paleoenvironmental Research; Springer: Dordrecht, The Netherlands, 2011; p. 11. [Google Scholar]
- Fritts, H.C. Tree Rings and Climate; Academic Press: London, UK, 1976; pp. 1–567. [Google Scholar]
- Juknys, R.; Stravinskiene, V.; Vencloviene, J. Tree-ring analysis for the assessment of anthropogenic changes and trends. Environ. Monit. Assess. 2002, 77, 81–97. [Google Scholar] [CrossRef]
- Wilczyński, S. The variation of tree−ring widths of Scots pine (Pinus sylvestris L.) affected by air pollution. Eur. J. For. Res. 2006, 125, 213–219. [Google Scholar] [CrossRef]
- Elling, W.; Dittmar, C.; Pfaffelmoser, K.; Rotzer, T. Dendroecological assessment of the complex causes of decline and recovery of the growth of silver fir (Abies alba Mill.) in Southern Germany. For. Ecol. Manag. 2009, 257, 1175–1187. [Google Scholar] [CrossRef]
- Stravinskiene, V.; Bartkevicius, E.; Plausinyte, E. Dendrochronological research of Scots pine (Pinus sylvestris L.) radial growth in vicinity of industrial pollution. Dendrochronologia 2013, 31, 179–186. [Google Scholar] [CrossRef]
- Sensuła, B.; Wilczyński, S. Records of Anthropogenic Pollution in Silesia Captured in Scots Pine Tree Rings: Analysis by Radiocarbon, Stable Isotopes, and Basal Area Increment Analysis. Water Air Soil Pollut. 2022, 233, 143. [Google Scholar] [CrossRef]
- Wilczyński, S. Record of the changes in environment by Scots pine and Jack pine trees. Sylwan 2020, 164, 583–593. [Google Scholar] [CrossRef]
- Lindholm, M.; Timonen, M.; Meriläinen, J. Extracting mid-summer temperatures from ring-width chronologies of living pines at the northern forest limit in Fennoscandia. Dendrochronologia 1996, 14, 99–113. [Google Scholar]
- Wilczyński, S.; Skrzyszewski, J. The climatic signal in tree-rings of Scots pine (Pinus sylvestris L.) from foot-hills of the Sudetic Mountains, southern Poland. For. Cent. 2002, 121, 15–24. [Google Scholar] [CrossRef]
- Tuovinen, M. Response of tree−ring width and density of Pinus sylvestris to climate beyond the continuous northern forest line in Finland. Dendrochronologia 2005, 22, 83–91. [Google Scholar] [CrossRef]
- Helama, S.; Mielikäinen, K.; Timonen, M.; Herva, H.; Tuomenvirta, H.; Veneäläinen, A. Regional climatic signals in Scots pine growth with insights into snow and soil associations. Dendrobiology 2013, 70, 27–34. [Google Scholar] [CrossRef]
- Juknys, R.; Augustaitis, A.; Vencloviene, J.; Kliučius, A.; Vitas, A.; Bartkevičius, E.; Jurkonis, N. Dynamic response of tree growth to changing environmental pollution. Eur. J. For. Res. 2014, 133, 713–724. [Google Scholar] [CrossRef]
- Ermich, K. Studies on the seasonal course of the radial increment of Pinus silvestris L. and Quercus robur L. Acta Soc. Bot. Pol. 1959, 28, 15–63. [Google Scholar] [CrossRef] [Green Version]
- Andersson, E. Cone and Seed Studies in Norway Spruce (Picea abies (L.) Karst). In Studia Forestalia Suecica; Skogshögskolan: Stockholm, Sweden, 1965; p. 23. [Google Scholar]
- Fober, H. Relation between climatic factors and Scots pine (Pinus sylvestris L.) cone crops in Poland. Arbor. Kórnickie 1976, 21, 367–374. [Google Scholar]
- Chałupka, W.; Giertych, M.; Królikowski, Z. The effect of cone crops on growth in Scot pine on tree diameter increment. Arbor. Kórnickie 1976, 21, 361–366. [Google Scholar]
- Hanninen, H.; Tanino, K. Tree seasonality in a warming climate. Trends Plant Sci. 2011, 16, 412–416. [Google Scholar] [CrossRef]
- Rossi, S.; Morin, H.; Deslauriers, A.; Plourde, P.Y. Predicting xylem phenology in back spruce under climate warming. Glob. Chang. Biol. 2011, 17, 614–625. [Google Scholar] [CrossRef]
- Rossi, S.; Deslauriers, A.; Anfodillo, T.; Carraro, V. Evidence of threshold temperatures for xylogenesis in conifers at high altitudes. Oecologia 2007, 152, 1–12. [Google Scholar] [CrossRef]
- Gruber, A.; Zimmermann, J.; Wieser, G.; Oberhuber, W. Effects of climate variables on intra-annual stem radial increment in Pinus cembra along the alpine treeline ecotone. Ann. For. Sci. 2009, 66, 503. [Google Scholar] [CrossRef] [Green Version]
- Swidrak, I.; Gruber, A.; Kofler, W.; Oberhuber, W. Effects of environmental conditions on onset of xylem growth in Pinus sylvestris under drought. Tree Physiol. 2011, 31, 483–493. [Google Scholar] [CrossRef] [Green Version]
- Oberhuber, W.; Gruber, A.; Kofler, W.; Swidrak, I. Radial stem growth in response to microclimate and soil moisture in a drought-prone mixed coniferous at an inner Alpine site. Eur. J. Res. 2014, 133, 467–479. [Google Scholar] [CrossRef] [Green Version]
- Richter, K.; Eckstein, D.; Holmes, R.L. The dendrochronological signal of the pine trees, Pinus sp. in Spain. Tree-Ring Bull. 1991, 51, 1–13. [Google Scholar]
- Pilcher, P.; Oberhuber, W. Radial growth response of coniferous forest trees in an inner Alpine environment to heat-wave in 2003. For. Ecol. Manag. 2007, 242, 1513–1523. [Google Scholar]
- Thabeet, A.; Vennetier, M.; Gadbin-Henry, C.; Denelle, N.; Roux, M.; Caraglio, Y.; Vila, B. Response of Pinus sylvestris L. to recent climatic events in the French Mediterranean region. Trees 2009, 23, 843–853. [Google Scholar] [CrossRef] [Green Version]
- Irvine, J.; Perks, M.P.; Magnani, F.; Grace, J. The response of Pinus sylvestris to drought: Stomatal control of transpiration and hydraulic conductance. Tree Physiol. 1998, 18, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Cinnirella, S.; Magnani, F.; Saracino, A.; Borghetti, M. Response of a mature Pinus laricio plantation to a three-year restriction of water supply: Structural and functional acclimation to drought. Tree Physiol. 2002, 22, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Pensa, M.; Salminen, H.; Jalkanen, R. A 250-year-long height-increment chronology for Pinus sylvestris at the northern coniferous timberline: A novel tool for reconstructing past summer temperatures? Dendrochronologia 2005, 22, 75–81. [Google Scholar] [CrossRef]
- Gruber, A.; Strobl, S.; Veit, B.; Oberhuber, W. Impact of drought on the temporal dynamics of wood formation in Pinus sylvestris. Tree Physiol. 2010, 30, 490–501. [Google Scholar] [CrossRef]
- Cook, E.R.; Holmes, R.L. User’s manual for program ARSTAN. In Tree−Ring Chronologies of Western North America: California, Eastern Oregon a Northern Great Basin; Chronology; Holmes, R.L., Adams, R.K., Fritts, H.C., Eds.; Laboratory of Tree-Ring Research, University of Arizona: Tucson, AZ, USA, 1986; Volume 6, pp. 50–56. [Google Scholar]
- Pålnäsvägen, Sweden. Available online: https://www.cybis.se (accessed on 24 October 2022).
- Holmes, R.L. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bull. 1983, 43, 69–78. [Google Scholar]
- Green, J. Wood cellulose. In Methods in Carbohydrate Chemistry 3; Whistler, R.L., Ed.; Academic Press: New York, NY, USA, 1963; pp. 9–21. [Google Scholar]
- Schweingruber, F.H.; Eckstein, D.; Serre-Bachet, F.; Bräker, O.U. Identification, presentation and interpretation of event years and pointer years in dendrochronology. Dendrochronologia 2009, 8, 9–38. [Google Scholar]
- Saurer, M.; Spahni, R.; Frank, D.C.; Joos, F.; Leuenberger, M.; Loader, N.J.; McCarroll, D.; Gagen, M.; Poulter, B.; Siegwolf, R.T.W.; et al. Spatial variability and temporal trends in water-use efficiency of European forests. Glob. Chang. Biol. 2014, 20, 3700–3712. [Google Scholar] [CrossRef]
- Biondi, F. Evolutionary and moving response functions in dendroclimatology. Dendrochronologia 1997, 15, 139–150. [Google Scholar]















Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sensuła, B.; Wilczyński, S. Dynamics Changes in Basal Area Increment, Carbon Isotopes Composition and Water Use Efficiency in Pine as Response to Water and Heat Stress in Silesia, Poland. Plants 2022, 11, 3569. https://doi.org/10.3390/plants11243569
Sensuła B, Wilczyński S. Dynamics Changes in Basal Area Increment, Carbon Isotopes Composition and Water Use Efficiency in Pine as Response to Water and Heat Stress in Silesia, Poland. Plants. 2022; 11(24):3569. https://doi.org/10.3390/plants11243569
Chicago/Turabian StyleSensuła, Barbara, and Sławomir Wilczyński. 2022. "Dynamics Changes in Basal Area Increment, Carbon Isotopes Composition and Water Use Efficiency in Pine as Response to Water and Heat Stress in Silesia, Poland" Plants 11, no. 24: 3569. https://doi.org/10.3390/plants11243569