
Assessing Seed Longevity of the Invasive Weed Navua Sedge (Cyperus aromaticus), by Artificial Ageing
 ,
,
Abstract
:1. Introduction
2. Results
3. Discussion
4. Materials and Methods
4.1. Seed Collection
4.2. Preparation
4.3. Seed Rehydration
4.4. Seed Ageing
4.5. Germination Test
4.6. Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Kraehmer, H.; Jabran, K.; Mennan, H.; Chauhan, B.S. Global distribution of rice weeds—A review. Crop Prot. 2016, 80, 73–86. [Google Scholar] [CrossRef]
- Black, I. Navua sedge in pastures in Fiji. Aust. Weeds 1984, 3, 16–19. [Google Scholar]
- Shi, B.; Osunkoya, O.O.; Chadha, A.; Florentine, S.K.; Dhileepan, K. Biology, Ecology and Management of the Invasive Navua Sedge (Cyperus aromaticus)—A Global Review. Plants 2021, 10, 1851. [Google Scholar] [CrossRef] [PubMed]
- Karan, B. Studies of Navua Sedge (Cyperus aromaticus). 1. Review of the problem and study of morphology, seed output and germination. Fiji Agric. J. 1975, 37, 59–67. [Google Scholar]
- Kerr, D.V.; Fell, R.F.; Murray, A.J.; Chaseling, J. An assessment of factors associated with increased productivity of dairy farms in Fiji. Asian-Australas. J. Anim. Sci. 1995, 8, 481–487. [Google Scholar] [CrossRef]
- Vitelli, J.S.; Madigan, B.A.; van Haaren, P.E. Control techniques and management strategies for the problematic Navua sedge (Cyperus aromaticus). Invasive Plant Sci. Manag. 2010, 3, 315–326. [Google Scholar] [CrossRef]
- Chadha, A.; Osunkoya, O.; Shi, B.; Florentine, S.K.; Dhileepan, K. Soil seed bank dynamics of pastures invaded by Navua sedge (Cyperus aromaticus) in tropical north Queensland. Front. Agron. 2022, 47, 897417. [Google Scholar] [CrossRef]
- Thompson, K.; Bakker, J.P.; Bekker, R.M.; Hodgson, J.G. Ecological correlates of seed persistence in soil in the north-west European flora. J. Ecol. 1998, 86, 163–169. [Google Scholar] [CrossRef]
- Zhou, W.; Chen, F.; Luo, X.; Dai, Y.; Yang, Y.; Zheng, C.; Yang, W.; Shu, K. A matter of life and death: Molecular, physiological, and environmental regulation of seed longevity. Plant Cell Environ. 2020, 43, 293–302. [Google Scholar] [CrossRef]
- Waterworth, W.M.; Bray, C.M.; West, C.E. The importance of safeguarding genome integrity in germination and seed longevity. J. Exp. Bot. 2015, 66, 3549–3558. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.-S.; Velasco-Punzalan, M.; Pacleb, M.; Valdez, R.; Kretzschmar, T.; McNally, K.L.; Ismail, A.M.; Cruz, P.C.S.; Sackville Hamilton, N.R.; Hay, F.R. Variation in seed longevity among diverse Indica rice varieties. Ann. Bot. 2019, 124, 447–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Presotto, A.; Hernández, F.; Casquero, M.; Vercellino, R.; Pandolfo, C.; Poverene, M.; Cantamutto, M. Seed bank dynamics of an invasive alien species, Helianthus annuus L. J. Plant Ecol. 2020, 13, 313–322. [Google Scholar] [CrossRef]
- Mašková, T.; Poschlod, P. Soil seed bank persistence across time and burial depth in calcareous grassland habitats. Front. Plant Sci. 2022, 12, 790867. [Google Scholar] [CrossRef] [PubMed]
- Long, R.L.; Panetta, F.D.; Steadman, K.J.; Probert, R.; Bekker, R.M.; Brooks, S.; Adkins, S.W. Seed persistence in the field may be predicted by laboratory-controlled aging. Weed Sci. 2008, 56, 523–528. [Google Scholar] [CrossRef]
- Sethi, R.; Kaur, N.; Singh, M. Morphological and physiological characterization of seed heteromorphism in Medicago denticulata Willd. Plant Physiol. Rep. 2020, 25, 107–119. [Google Scholar] [CrossRef]
- Fenollosa, E.; Jené, L.; Munné-Bosch, S. A rapid and sensitive method to assess seed longevity through accelerated aging in an invasive plant species. Plant Methods 2020, 16, 1–11. [Google Scholar] [CrossRef]
- Brooks, S.J.; Brazier, D.A.; Warren, C. Estimating tropical weed seed longevity with a laboratory test. In Proceedings of the 22nd Australian Weeds Conference, Adelaide, Australia, 22–29 September 2022. [Google Scholar]
- Chadha, A.; Florentine, S.K.; Dhileepan, K.; Dowling, K.; Turville, C. Germination biology of three populations of Navua sedge (Cyperus aromaticus). Weed Sci. 2021, 69, 69–81. [Google Scholar] [CrossRef]
- Bebawi, F.F.; Campbell, S.D.; Mayer, R.J. Gamba grass (Andropogon gayanus Kunth.) seed persistence and germination temperature tolerance. Rangel. J. 2018, 40, 463–472. [Google Scholar] [CrossRef]
- Vargas, A.; Agostinetto, D.; Zandoná, R.; Fraga, D.; Avila Neto, R. Longevity of horseweed seed bank depending on the burial depth. Planta Daninha 2018, 36. [Google Scholar] [CrossRef]
- Sarabi, V. Factors that influence the level of weed seed predation: A review. Weed Biol. Manag. 2019, 19, 61–74. [Google Scholar] [CrossRef]
- Chung, N.-J.; Paek, N.-C. Photoblastism and ecophysiology of seed germination in weedy rice. Agron. J. 2003, 95, 184–190. [Google Scholar] [CrossRef]
- Mavi, K.; Demir, I.; Matthews, S. Mean germination time estimates the relative emergence of seed lots of three cucurbit crops under stress conditions. Seed Sci. Technol. 2010, 38, 14–25. [Google Scholar] [CrossRef]
- Bailly, C.; Bogatek-Leszczynska, R.; Côme, D.; Corbineau, F. Changes in activities of antioxidant enzymes and lipoxygenase during growth of sunflower seedlings from seeds of different vigour. Seed Sci. Res. 2002, 12, 47–55. [Google Scholar] [CrossRef]
- Long, R.L.; Kranner, I.; Panetta, F.D.; Birtic, S.; Adkins, S.W.; Steadman, K.J. Wet-dry cycling extends seed persistence by re-instating antioxidant capacity. Plant Soil 2011, 338, 511–519. [Google Scholar] [CrossRef]
- Zhang, K.; Zhang, Y.; Sun, J.; Meng, J.; Tao, J. Deterioration of orthodox seeds during ageing: Influencing factors, physiological alterations and the role of reactive oxygen species. Plant Physiol. Biochem. 2021, 158, 475–485. [Google Scholar] [CrossRef]
- Newton, R.; Hay, F.R.; Probert, R.J. Protocol for Comparative Seed Longevity Testing; The Royal Botanic Gardens: London, UK, 2009. [Google Scholar]
- Saatkamp, A.; Affre, L.; Dutoit, T.; Poschlod, P. Germination traits explain soil seed persistence across species: The case of Mediterranean annual plants in cereal fields. Ann. Bot. 2011, 107, 415–426. [Google Scholar] [CrossRef]
- Van Waes, J.M.; Debergh, P.C. Adaptation of the tetrazolium method for testing the seed viability, and scanning electron microscopy study of some Western European orchids. Physiol. Plant. 1986, 66, 435–442. [Google Scholar] [CrossRef]
- Kader, M. A comparison of seed germination calculation formulae and the associated interpretation of resulting data. J. Proceeding R. Soc. N. S. W. 2005, 138, 65–75. [Google Scholar]


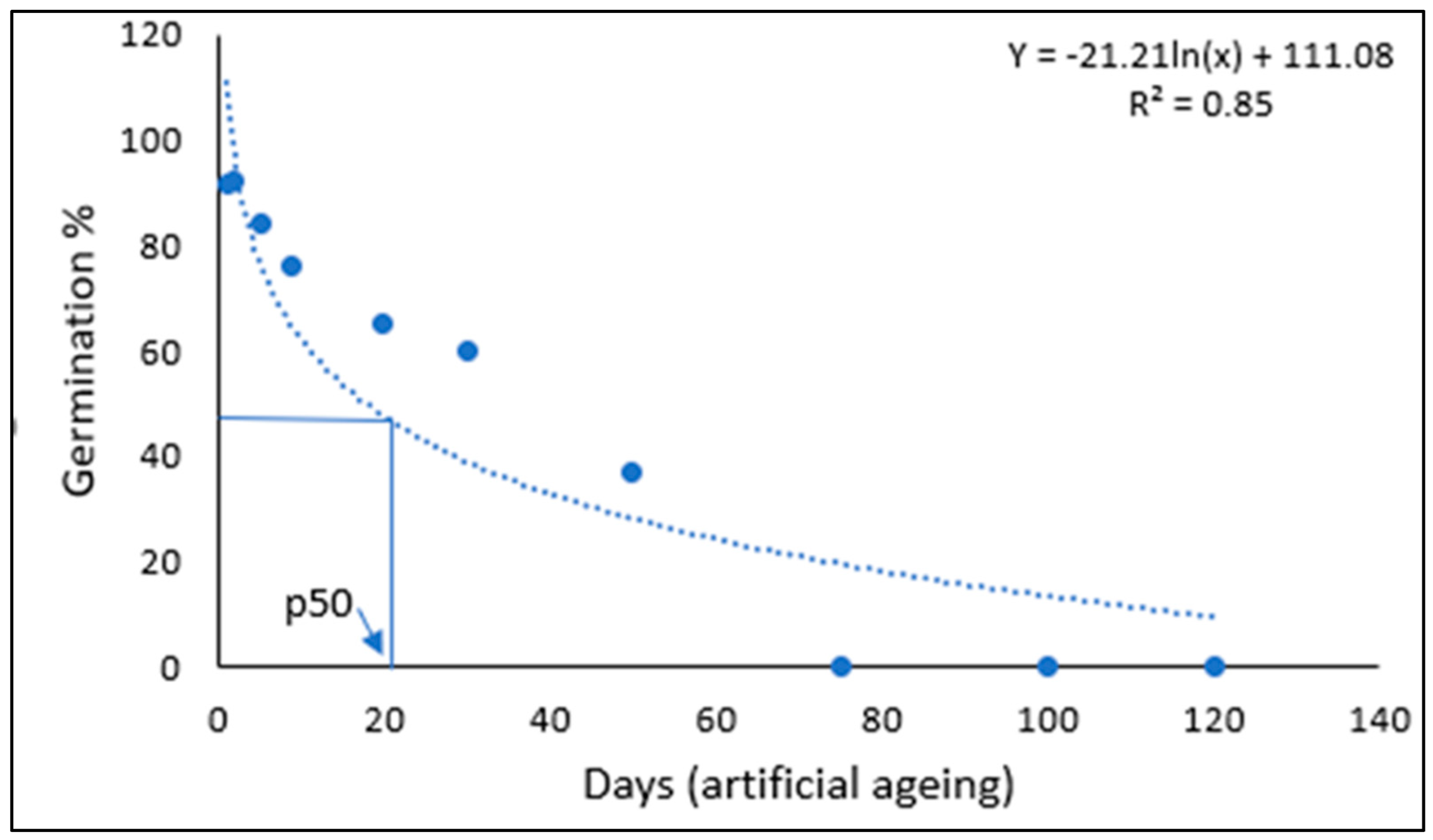{kind=link}
| Ageing Times | Mean Germination % | Mean Germination Time (days) | Germination Index |
|---|---|---|---|
| Day 0 | 93.3 ± 2. 11 | 12.9 ± 0.66 | 1.6 ± 0.11 |
| Day 1 | 91.7 ± 1.67 | 10.4 ± 0.31 | 1.9 ± 0.07 |
| Day 2 | 92.5 ± 2.14 | 10.5 ± 0.48 | 1.8 ± 0.07 |
| Day 5 | 84.2 ± 2.01 | 12.9 ± 0.84 | 1.4 ± 0.1 |
| Day 9 | 75.8 ± 3.01 | 12.9 ± 0.33 | 1.2 ± 0.06 |
| Day 20 | 65.0 ± 2.24 | 14.2 ± 0.69 | 1.0 ± 0.06 |
| Day 30 | 60.0 ± 4.47 | 14.6 ± 0.45 | 0.9 ± 0.07 |
| Day 50 | 36.7 ± 6.67 | 16.8 ± 0.56 | 0.5 ± 0.09 |
| Day 75 | 0.0 ± 0.0 | NA | NA |
| Day 100 | 0.0 ± 0.0 | NA | NA |
| Day 125 | 0.0 ± 0.0 | NA | NA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chadha, A.; Florentine, S.K.; Dhileepan, K.; Turville, C. Assessing Seed Longevity of the Invasive Weed Navua Sedge (Cyperus aromaticus), by Artificial Ageing. Plants 2022, 11, 3469. https://doi.org/10.3390/plants11243469
Chadha A, Florentine SK, Dhileepan K, Turville C. Assessing Seed Longevity of the Invasive Weed Navua Sedge (Cyperus aromaticus), by Artificial Ageing. Plants. 2022; 11(24):3469. https://doi.org/10.3390/plants11243469
Chicago/Turabian StyleChadha, Aakansha, Singarayer K. Florentine, Kunjithapatham Dhileepan, and Christopher Turville. 2022. "Assessing Seed Longevity of the Invasive Weed Navua Sedge (Cyperus aromaticus), by Artificial Ageing" Plants 11, no. 24: 3469. https://doi.org/10.3390/plants11243469