
Assessment of the Hormetic Effect of Arsenic on Growth and Physiology of Two Cultivars of Maize (Zea mays L.)

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Effect of Arsenic on Growth and Relative Water Content
2.2. Effect of Arsenic on the Content of Photosynthetic Pigments
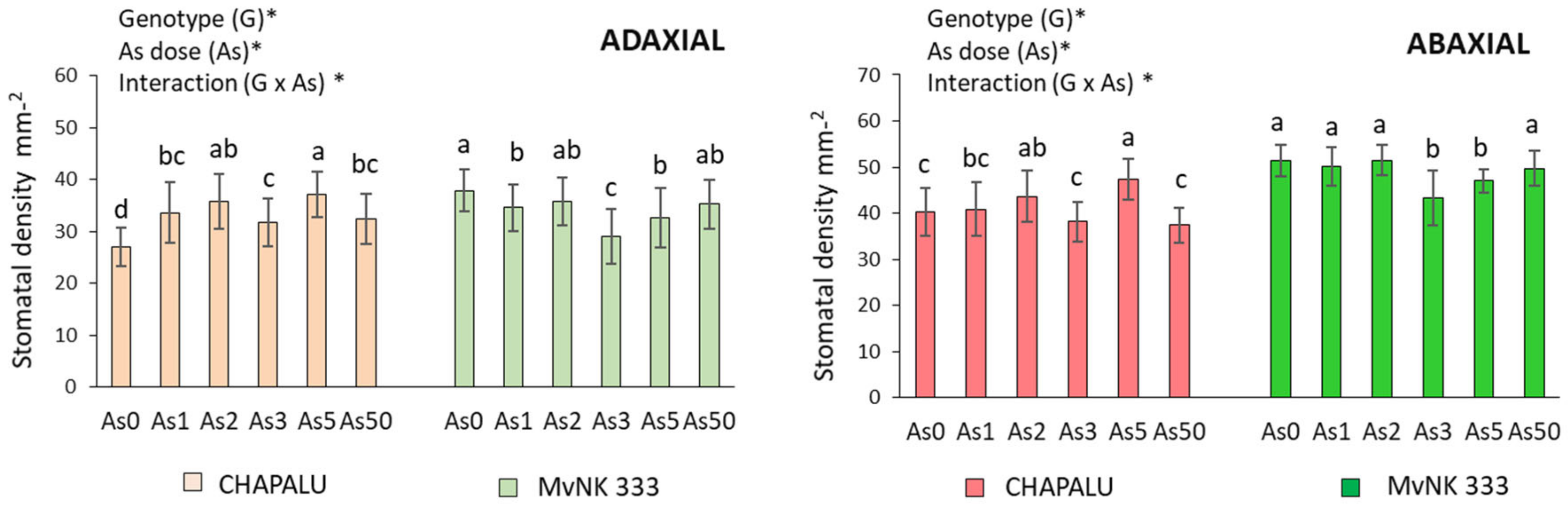
2.3. Effect of Arsenic on Stomatal Density and Stomata Size
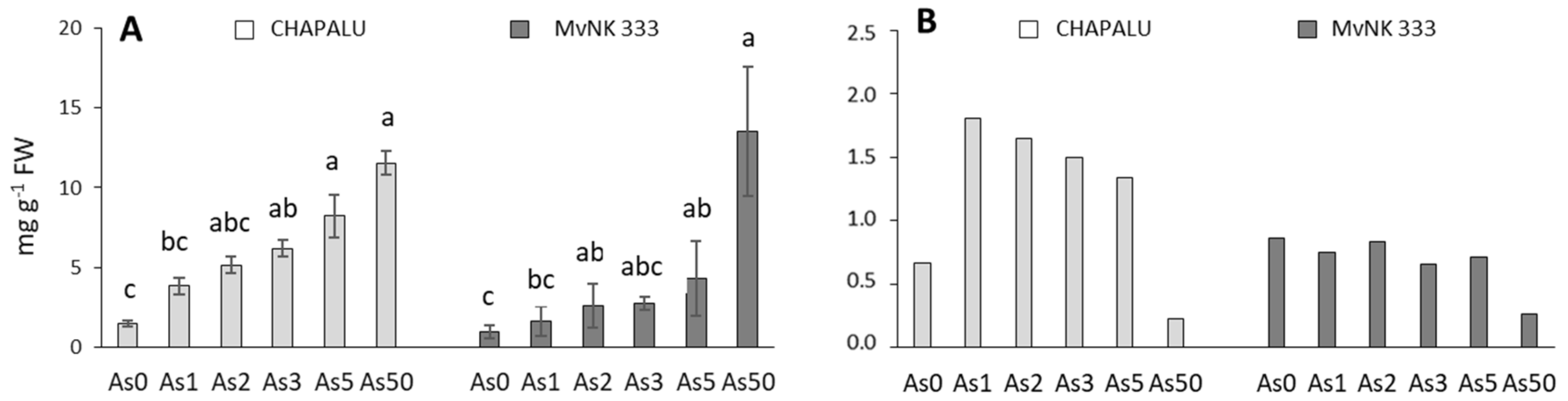
2.4. Arsenic Accumulation in Shoots
2.5. Evaluation of the Effect of Genotype and Arsenic Dose on Tested Parameters
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growth Condition
4.2. Determination of Growth Parameters and Tolerance Index
4.3. Determination of Photosynthetic Pigments
4.4. Determination of Relative Water Content (RWC)
4.5. Determination of Number and Size of Stomata
4.6. Determination of As in Shoots and Soil
4.7. Determination of Bioacumulation Factor for As
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- EPA. Environmental Protection Agency of U.S. 2012. Available online: http://www.epa.gov/iris/subst/0278.htm (accessed on 14 June 2012).
- Meharg, A.A.; Hartley-Whitaker, J. Arsenic uptake and metabolism in arsenic resistant and nonresistant plant species. New Phytol. 2002, 154, 29–43. [Google Scholar] [CrossRef] [Green Version]
- Finnegan, P.M.; Chen, W. Arsenic toxicity. The effects on plant metabolism. Front. Physiol. 2012, 3, 182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, M.; Yang, J. Characteristic of binding forms of arsenic in polluted rice seed and their stability. Chin. J. Appl. Ecol. 2002, 13, 1141–1144. (In Chinese) [Google Scholar]
- Gulz, P.A.; Gupta, S.K.; Schulin, R. Arsenic accumulation of common plants from contaminated soils. Plant Soil 2005, 272, 337–347. [Google Scholar] [CrossRef]
- Pickering, I.J.; Gumaelius, L.; Harris, H.H.; Prince, R.C.; Hirsch, G.; Banks, J.A.; Salt, D.E.; George, G.N. Localizing the biochemical transformations of arsenate in a hyperaccumulating fern. Environ. Sci. Technol. 2006, 40, 5010–5014. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.Y.; McGrath, S.P.; Zhao, F.J. Rapid reduction of arsenate in the medium mediated by plant roots. New Phytol. 2007, 176, 590–599. [Google Scholar] [CrossRef] [PubMed]
- Marwa, E.M.M.; Meharg, A.A.; Rice, C.M. Risk assessment of potentially toxic elements in agricultural soils and maize tissues from selected districts in Tanzania. Sci. Total Environ. 2012, 416, 180–186. [Google Scholar] [CrossRef]
- Rosas-Castor, J.M.; Guzmán-Mar, J.L.; Hernández-Ramírez, A.; Garza-González, M.T.; Hinojosa-Reyes, L. Arsenic accumulation in maize crop (Zea mays): A review. Sci. Total Environ. 2014, 488–489, 176–187. [Google Scholar] [CrossRef]
- Sadee, B.A.; Foulkes, M.E.; Hill, S.J. An evaluation of extraction techniques for arsenic in staple diets (fish and rice) utilising both classical and enzymatic extraction methods. Food Addit. Contam. A 2016, 33, 433–441. [Google Scholar] [CrossRef] [Green Version]
- Garg, N.; Singla, P. Arsenic toxicity in crop plants: Physiological effects and tolerance mechanisms. Environ. Chem. Lett. 2011, 9, 303–321. [Google Scholar] [CrossRef]
- Päivöke, A.; Simola, L.K. Arsenate toxicity to Pisum sativum: Mineral nutrients, chlorophyll content, and phytase activity. Ecotoxicol. Environ. Saf. 2001, 49, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Zemanová, V.; Popov, M.; Pavlíková, D.; Kotrba, P.; Hnilička, F.; Česká, J.; Pavlík, M. Effect of arsenic stress on 5-methylcytosine, photosynthetic parameters and nutrient content in arsenic hyperaccumulator Pteris cretica (L.) var. Albo-lineata. MC Plant Biol. 2020, 20, 130. [Google Scholar] [CrossRef] [PubMed]
- Vezza, M.E.; Llanes, A.; Travaglia, C.; Agostini, E.; Talano, M.A. Arsenic stress effects on root water absorption in soybean plants: Physiological and morphological aspects. Plant Physiol. Biochem. 2018, 123, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Abbas, G.; Murtaza, B.; Bibi, I.; Shahid, M.; Niazi, N.; Khan, M.; Amjad, M.; Hussain, M.; Tahir, N. Arsenic uptake, toxicity, detoxification, and speciation in plants: Physiological, biochemical, and molecular aspects. Int. J. Environ. Res. 2018, 15, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carbonell-Barrachina, A.A.; Burló, F.; López, E.; Martínez-Sánchez, F. Arsenic toxicity and, in radish as affected by arsenic chemical speciation. J. Environ. Sci. Health B 1999, 34, 661–679. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, E.J.; Blain, R.B. Hormesis and plant biology. Environ. Pollut. 2009, 157, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, E.J.; Mattson, M.P. Hormesis provides a generalized quantitative estimate of biological plasticity. J. Cell Commun. Signal. 2011, 5, 25–38. [Google Scholar] [CrossRef] [Green Version]
- Burló, F.; Guijarro, I.; Carbonell-Barrachina, A.A.; Valero, D.; Martínez-Sánchez, F. Arsenic species: Effects on and accumulation by tomato plants. J. Agric. Food Chem. 1999, 47, 1247–1253. [Google Scholar] [CrossRef]
- Sushant, K.S.; Ghosh, A.K. Effect of arsenic on photosynthesis, growth and its accumulation in the tissues of Allium cepa (Onion). Int. J. Environ. Eng. Manag. 2010, 1, 39–50. [Google Scholar] [CrossRef]
- Ren, J.H.; Sun, H.J.; Wang, S.F.; Luo, J.; Ma, L.Q. Interactive effects of mercury and arsenic on their uptake, speciation and toxicity in rice seedling. Chemosphere 2014, 117, 737–744. [Google Scholar] [CrossRef]
- Evans, G.; Evans, J.; Redman, A.; Johnson, N. Unexpected beneficial effects of arsenic on corn roots grown in culture. Environ. Chem. 2005, 2, 167–170. [Google Scholar] [CrossRef]
- Stoeva, N.; Berova, M.; Zlatev, Z. Physiological response of maize to arsenic contamination. Biol. Plant. 2003, 47, 449–452. [Google Scholar] [CrossRef]
- Allender, W.J.; Cresswell, G.C.; Kaldor, J.; Kennedy, I.R. Effect of lithium and lanthium on herbicide induced hormesis in hydrophonically-grown cotton and corn. J. Plant Nutr. 1997, 20, 81–95. [Google Scholar] [CrossRef]
- Pinto, A.P.; Mota, A.M.; Varennes, A.; de Pinto, F.C. Influence of organic matter on the uptake of cadmium, zinc, copper and iron by sorghum plants. Sci. Total Environ. 2004, 326, 239–247. [Google Scholar] [CrossRef]
- Poschenrieder, C.; Cabot, C.; Martos, S.; Gallego, B.; Barceló, J. Do toxic ions induce hormesis in plants? Plant Sci. 2013, 212, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Shahid, M.; Niazi, N.K.; Rinklebe, J.; Bundschuh, J.; Dumt, C.; Pinelli, E. Trace elements-induced phytohormesis: A critical review and mechanistic interpretation. Crit. Rev. Environ. Sci. Technol. 2020, 50, 1984–2015. [Google Scholar] [CrossRef]
- Małkowski, E.; Sitko, K.; Szopiński, M.; Gieroń, Ż.; Pogrzeba, M.; Kalaji, H.M.; Zieleźnik-Rusinowska, P. Hormesis in Plants: The role of oxidative stress, auxins and photosynthesis in corn treated with Cd or Pb. Int. J. Mol. Sci. 2020, 21, 2099. [Google Scholar] [CrossRef] [Green Version]
- Salinitro, M.; Mattarello, G.; Guardigli, G.; Odajiu, M.; Tassoni, A. Induction of hormesis in plants by urban trace metal pollution. Sci. Rep. 2021, 11, 20329. [Google Scholar] [CrossRef]
- Carvalho, M.E.A.; Castro, P.R.C.; Azevedo, R.A. Hormesis in plants under Cd exposure: From toxic to beneficial element? J. Hazard. Mater. 2020, 84, 121434. [Google Scholar] [CrossRef]
- Hayes, D.P. Nutritional hormesis. Eur. J. Clin. Nutr. 2007, 61, 147–159. [Google Scholar] [CrossRef]
- Piršelová, B.; Galuščáková, Ľ.; Lengyelová, L. Hormetická odpoveď rastlín na ióny kovov a polokovov. Chem. Listy 2018, 112, 317–323. [Google Scholar]
- Agathokleous, E. The rise and fall of photosynthesis: Hormetic dose response in plants. J. For. Res. 2021, 32, 889–898. [Google Scholar] [CrossRef]
- Erofeeva, E.A. Method for assessing the frequency of hormetic trade-offs in plants. MethodsX 2022, 9, 101610. [Google Scholar] [CrossRef]
- Agathokleous, E.; Zhong, Z.; Peñuelas, F.J. Chlorophyll hormesis: Are chlorophylls major components of stress biology in higher plants? Sci. Total Environ. 2020, 726, 138637. [Google Scholar] [CrossRef] [PubMed]
- Agathokleous, E.; Calabrese, E.J. Hormesis: The dose response for the 21st Century: The future has arrived. Toxicology 2019, 425, 152249. [Google Scholar] [CrossRef]
- Rizwan, M.; Ali, S.; Abbas, F.; Adrees, M.; Zia-Ur-Rehman, M.; Gill, R.A.; Ali, B. Role of organic and inorganic amendments in alleviating heavy metal stress in oil seed crops. In Oil Seed Crops: Yield and Adaptations under Environmental Stress, 1st ed.; Ahmad, P., Ed.; John Wiley & Sons Ltd.: Chichester, UK, 2017; pp. 224–235. [Google Scholar] [CrossRef]
- Kaya, C.; Ashraf, M.; Alyemeni, M.N.; Corpas, F.J.; Ahmad, P. Salicylic acid-induced nitric oxide enhances arsenic toxicity tolerance in maize plants by upregulating the ascorbate-glutathione cycle and glyoxalase system. J. Hazard. Mater. 2020, 399, 123020. [Google Scholar] [CrossRef]
- Mishra, S.; Stark, H.J.; Kupper, H. A different sequence of events than previously reported leads to arsenic-induced damage in Ceratophyllum demersum L. Metallomics 2014, 3, 444–454. [Google Scholar] [CrossRef] [Green Version]
- Patnaik, A.R.; Achary, V.M.M.; Panda, B.B. Chromium (Cr)-induced hormesis and genotoxicity are mediated through oxidative stress in root cells of Allium cepa L. Plant Growth Regul. 2013, 71, 57–170. [Google Scholar] [CrossRef]
- Mittler, R. ROS are good. Trends Plant Sci. 2017, 22, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Seabra, A.B.; Oliveira, H.C. How nitric oxide donors can protect plants in a changing environment: What we know so far and perspectives. AIMS Mol. Sci. 2016, 3, 692–718. [Google Scholar] [CrossRef]
- Jacobs, L.W.; Keeney, D.R. Arsenic—Phosphorus interaction in corn. Commun. Soil Sci. Plant Anal. 1970, 1, 85–93. [Google Scholar] [CrossRef]
- Lambkin, D.C.; Alloway, B.J. Arsenate-induced phosphate release from soils and its effect on plant phosphorus. Water Air Soil Pollut. 2003, 144, 41–56. [Google Scholar] [CrossRef]
- Du, L.; Xia, X.; Lan, M.; Liu, M.; Zhao, L.; Zhang, P.; Wu, Y. Influence of arsenic stress on physiological, biochemical, and morphological characteristics in seedlings of two cultivars of maize (Zea mays L.). Water Air Soil Pollut. 2017, 228, 55. [Google Scholar] [CrossRef]
- Ci, X.K.; Liu, H.L.; Hao, Y.B.; Zhang, J.W.; Liu, P.; Dong, S.T. Arsenic distribution, species, and its effect on maize growth treated with arsenate. J. Integr. Agric. 2012, 11, 416–423. [Google Scholar] [CrossRef]
- Li, C.X.; Feng, S.L.; Shao, Y.; Jiang, L.N.; Lu, X.Y.; Hou, X. Effects of arsenic on seed germination and physiological activities of wheat seedlings. J. Environ. Sci. 2007, 19, 725–732. [Google Scholar] [CrossRef]
- Jia, L.; Liu, Z.; Chen, W.; Ye, Y.; Yu, S.; He, X. Hormesis effects induced by cadmium on growth and photosynthetic performance in a hyperaccumulator, Lonicera japonica. Thunb. J. Plant Growth Regul. 2015, 34, 13–21. [Google Scholar] [CrossRef]
- Gusman, G.S.; Oliveira, J.A.; Farnese, F.S.; Cambraia, J. Arsenate and arsenite: The toxic effects on photosynthesis and growth of lettuce plants. Acta Physiol. Plant. 2013, 35, 1201–1209. [Google Scholar] [CrossRef]
- Sade, N.; Gebremedhin, A.; Moshelion, M. Risk-taking plants: Anisohydric behavior as a stress-resistance trait. Plant Signal Behav. 2012, 7, 767–770. [Google Scholar] [CrossRef] [Green Version]
- Gostin, I.N. Air pollution effects on the leaf structure of some fabaceae species. Not. Bot. Horti. Agrobot. Cluj-Napoca 2009, 37, 57–63. [Google Scholar] [CrossRef]
- Weryszko-Chmielewska, E.; Chwil, M. Lead induced histological and ultrastructural changes in the leaves of soybean (Glycine max (L.) Merr.). Soil Sci. Plant Nutr. 2005, 51, 203. [Google Scholar] [CrossRef]
- Farber, M.; Attia, Z.; Weiss, D. Cytokinin activity increases stomatal density and transpiration rate in tomato. J. Exp. Bot. 2016, 67, 6351. [Google Scholar] [CrossRef] [PubMed]
- Gálusová, T.; Piršelová, B.; Rybanský, Ľ.; Krasylenko, Y.; Mészáros, P.; Blehová, A.; Bardáčová, M.; Moravčíková, J.; Matušíková, I. Plasticity of soybean stomatal responses to arsenic and cadmium at the whole plant level. Pol. J. Environ. Stud. 2020, 29, 3569–3580. [Google Scholar] [CrossRef] [PubMed]
- Melo, H.C.; Castro, E.M.; Soares, A.M.; Melo, L.A.; Alves, J.D. Anatomical and physiological alterations in Setaria anceps Stapf ex Massey and Paspalum paniculatum L. under water deficit conditions. Hoehnea 2007, 34, 145. [Google Scholar] [CrossRef] [Green Version]
- Bertolino, L.T.; Caine, R.S.; Gray, J.E. Impact of stomatal density and morphology on water-use efficiency in a changing world. Front. Plant Sci. 2019, 10, 225. [Google Scholar] [CrossRef] [Green Version]
- Shi, G.; Caia, Q. Leaf plasticity in peanut (Arachis hypogaea L.) in response to heavy metal stress. Environ. Exp. Bot. 2009, 67, 112–117. [Google Scholar] [CrossRef]
- Imran, M.A.; Sajid, Z.A.; Chaudhry, M.N. Arsenic (As) Toxicity to germination and vegetative growth of sunflower (Helianthus annuus L.). Pol. J. Environ. Stud. 2015, 24, 1993–2002. [Google Scholar] [CrossRef]
- Vromman, D.; Flores-Bavestrello, A.; Šlejkovec, Z.; Lapaillea, S.; Teixeira-Cardoso, C.; Briceño, M.; Kumar, M.; Martínez, J.P.; Lutts, S. Arsenic accumulation and distribution in relation to young seedling growth in Atriplex atacamensis Phil. Sci. Total Environ. 2011, 412–413, 286–295. [Google Scholar] [CrossRef]
- Labancová, E.; Vivodová, Z.; Kučerová, D.; Lišková, D.; Kollárová, K. The cadmium tolerance development of poplar callus is influenced by silicon. Ecotoxicology 2020, 29, 987–1002. [Google Scholar] [CrossRef]
- Cedergreen, N.; Olesen, C.F. Can glyphosate stimulate photosynthesis? Pestic. Biochem. Physiol. 2010, 96, 140–148. [Google Scholar] [CrossRef]
- Wan, X.; Lei, M.; Chen, T.; Yang, J.; Liu, H.; Chen, Y. Role of transpiration in arsenic accumulation of hyperaccumulator Pteris vittata L. Environ. Sci. Pollut. Res. 2015, 22, 16631–16639. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Torii, K.U. Hormonal and environmental signals guiding stomatal development. BMC Biol. 2018, 16, 21. [Google Scholar] [CrossRef] [PubMed]
- Kasim, W.A. Changes induced by copper and cadmium in the anatomy and grain yield of Sorghum bicolor (L.) Moench. Int. J. Agric. Biol. 2006, 8, 123–128. [Google Scholar]
- Baryla, A.; Carrier, P.; Franck, F.; Coulomb, C.; Sahut, C.; Havaux, M. Leaf chlorosis in oilseed rape plants (Brassica napus) grown on cadmium-polluted soil: Causes and consequences for photosynthesis and growth. Planta 2015, 212, 696–709. [Google Scholar] [CrossRef] [PubMed]
- Aliu, S.; Rusinovci, I.; Doko, A.; Salihu, S.; Fetahu, S.; Elezi, F.; Gashi, B. Stomatal characteristics and their relationship to heavy metals in maize (Zea mays L.) seedlings. J. Food Agric. Environ. 2015, 13, 168–171. [Google Scholar] [CrossRef]
- Buscaroli, A. An overview of indexes to evaluate terrestrial plants for phytoremediation purposes. Ecol. Indic. 2017, 82, 367–380. [Google Scholar] [CrossRef]
- Zandstra, B.H.; de Kryger, T.A. Arsenic and lead residues in carrots from foliar applications of monosodium methanearsonate (MSMA): A comparison between mineral and organic soils, or from soil residues. Food Addit. Contam. 2007, 24, 34–42. [Google Scholar] [CrossRef]
- GB 2715-2005; Hygienic Standard for Grains. Ministry of Health of the People’s Republic of China (MHPRC): Beijing, China; Standardization Administration of China: Beijing, China, 2005.
- McLaughlin, M.J.; Hamon, R.E.; McLaren, R.G.; Speir, T.W.; Rogers, S.L. Review: A bioavailability–based rationale for controlling metal and metalloid contamination of agricultural land in Australia and New Zealand. Aust. J. Soil. Res. 2000, 38, 1037–1086. [Google Scholar] [CrossRef]
- Norra, S.; Berner, Z.A.; Agarwala, P.; Wagner, F.; Chandrasekharam, D.; Stüben, D. Impact of irrigation with arsenic rich groundwater on soil and crops: A geochemical case study in West Bengal delta plain, India. J. Appl. Geochem. 2005, 20, 1890–1906. [Google Scholar] [CrossRef]
- Cao, X.; Bai, L.; Zeng, X.; Zhang, J.; Wang, Y.; Wu, C.; Su, S. Is maize suitable for substitution planting in arsenic contaminated farmlands? Plant Soil Environ. 2019, 65, 425–434. [Google Scholar] [CrossRef] [Green Version]
- Erofeeva, E.A. Hormesis and paradoxical effects of wheat seedling (Triticum aestivum L.) parameters upon exposure to different pollutants in a wide range of doses. Dose Response 2014, 12, 121–135. [Google Scholar] [CrossRef]
- Sadiq, M. Solubility relationships of arsenic in calcareous soils and its uptake by corn. Plant Soil 1986, 91, 241–248. [Google Scholar] [CrossRef]
- Ritchie, S.W.; Hanway, J.J.; Benson, G.O. How a Corn Plant Develops; CES Special Report No. 48; Iowa State University: Ames, IA, USA, 1993; p. 21. [Google Scholar]
- Wilkins, D.A. The measurement of tolerance to edaphic factors by means of root length. New Phytol. 1978, 80, 623–633. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Meth. Enzymol. 1987, 148, 350–382. [Google Scholar] [CrossRef]
- Dhopte, A.M.; Manuel, L.M. Principles and Techniques for Plant Scientists, 1st ed.; Updesh Purohit for Agrobios (India): Odhpur, India, 2002; p. 373. [Google Scholar]
- Xu, Z.Z.; Zhou, G.S. Effects of water stress and nocturnal temperature on carbon allocation in the perennial grass, Leymus chinensis. Physiol. Plant. 2005, 123, 272–280. [Google Scholar] [CrossRef]
- ČSN EN 12457-4 (838005); Characterisation of waste. Leaching. Compliance test for leaching of granular waste materials and sludges. Part 4: One stage batch test at a liquid to solid ratio of 10 L/kg for materials with particle size below 10 mm (without or with size reduction). The Czech Office for Standards, Metrology and Testing (COSMT): Prague, Czech Republic, 2002.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chapalu | MvNK 333 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| TI (%) | As1 | As2 | As3 | As5 | As50 | As1 | As2 | As3 | As5 | As50 |
| Length of shoots | 103.59 | 109.70 | 104.82 | 102.66 | 64.75 | 112.28 | 121.64 | 128.19 | 139.19 | 112.20 |
| Fresh weight of shoots | 108.83 | 106.55 | 104.91 | 101.78 | 67.52 | 111.08 | 113.08 | 121.73 | 135.52 | 108.06 |
| Dry weight of shoots | 107.91 | 106.35 | 107.63 | 105.54 | 93.69 | 113.54 | 112.51 | 126.19 | 130.45 | 118.18 |
| Country | Regulation Item | Statutory Limit | Reference |
|---|---|---|---|
| Switzerland | fodder | 4 mg kg−1 DW | [5] |
| Canada | food crops | 1 mg kg−1 FW | [68] |
| China | rice | 0.15 mg kg−1 DW | [69] |
| Australia | cereals | 1 mg kg−1 FW | [70] |
| Germany | cerals | 1 mg kg−1 FW | [71] |
| India | cerals | 1 mg kg−1 FW | [71] |
| The Netherlands | cereals | 1 mg kg−1 FW | [71] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piršelová, B.; Galuščáková, Ľ.; Lengyelová, L.; Kubová, V.; Jandová, V.; Hegrová, J. Assessment of the Hormetic Effect of Arsenic on Growth and Physiology of Two Cultivars of Maize (Zea mays L.). Plants 2022, 11, 3433. https://doi.org/10.3390/plants11243433
Piršelová B, Galuščáková Ľ, Lengyelová L, Kubová V, Jandová V, Hegrová J. Assessment of the Hormetic Effect of Arsenic on Growth and Physiology of Two Cultivars of Maize (Zea mays L.). Plants. 2022; 11(24):3433. https://doi.org/10.3390/plants11243433
Chicago/Turabian StylePiršelová, Beáta, Ľudmila Galuščáková, Libuša Lengyelová, Veronika Kubová, Vilma Jandová, and Jitka Hegrová. 2022. "Assessment of the Hormetic Effect of Arsenic on Growth and Physiology of Two Cultivars of Maize (Zea mays L.)" Plants 11, no. 24: 3433. https://doi.org/10.3390/plants11243433