

Insight into Population Structure and Evolutionary Analysis of the Emerging Tomato Brown Rugose Fruit Virus

 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Results
2.1. Incidence and Sequencing
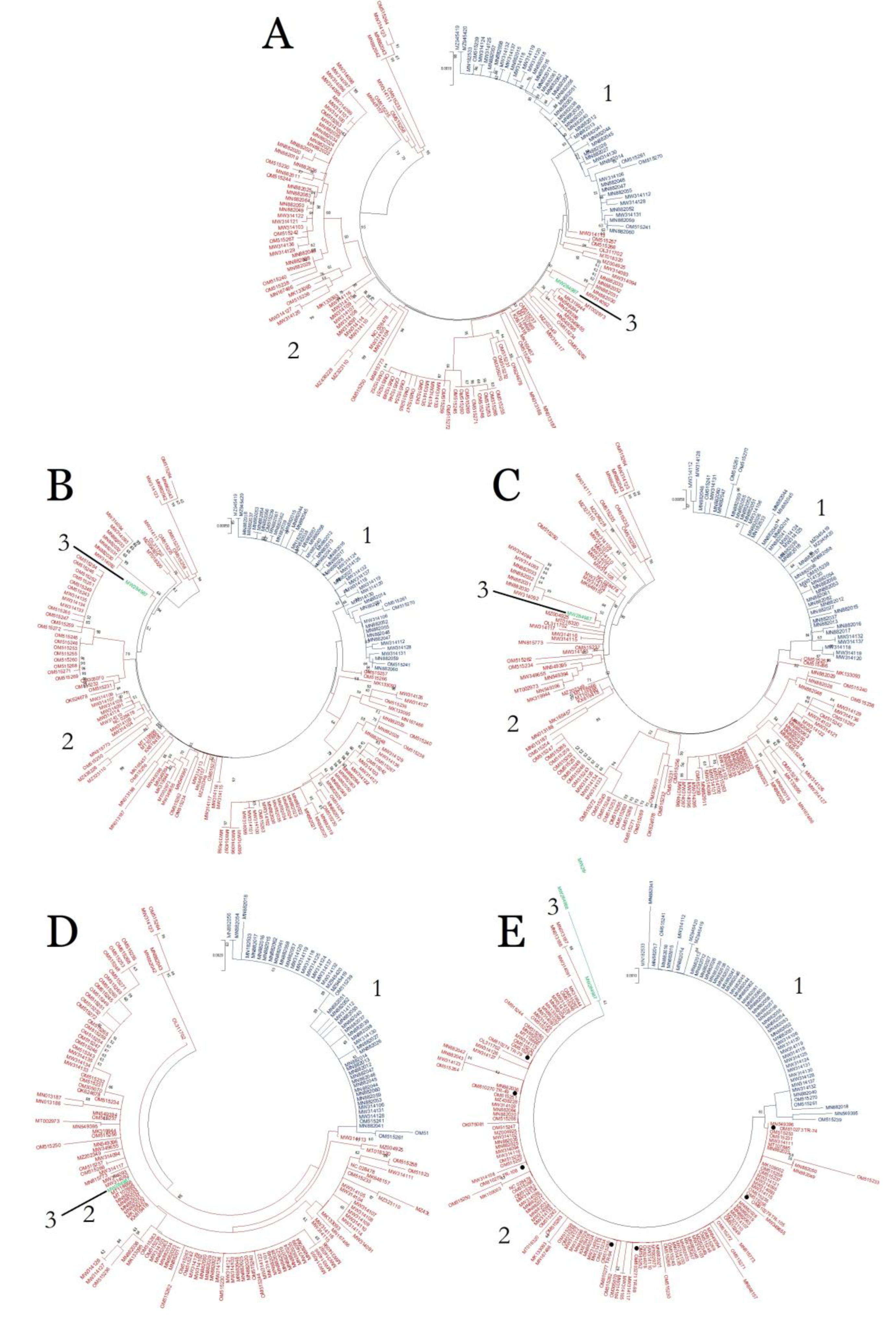
2.2. Recombination and Phylogenetic Analyses
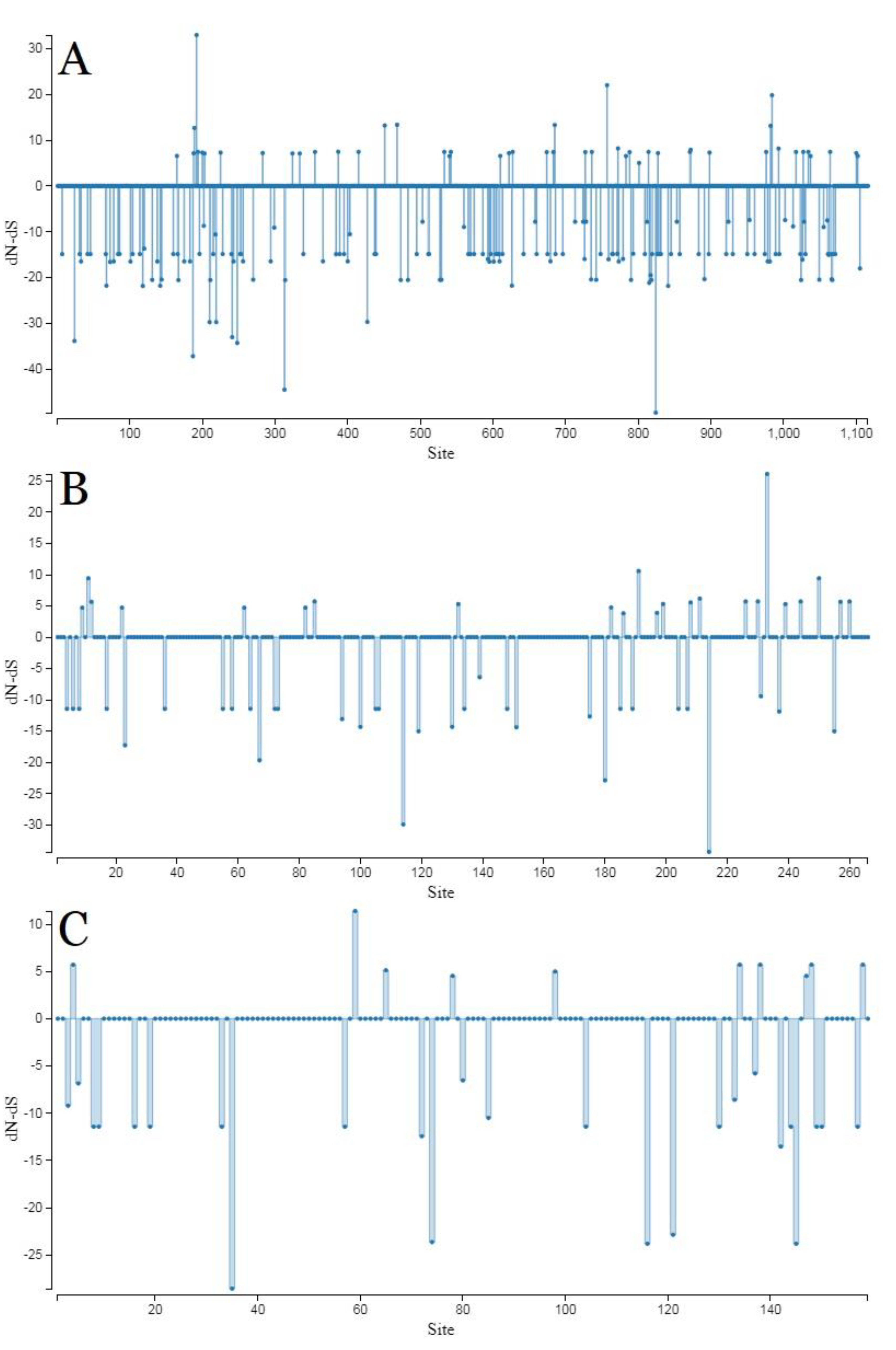
2.3. Genetic Variation and Selection Pressure Analyses
2.4. Neutrality Tests
2.5. Gene Flow and Genetic Separation among Populations
2.6. Molecular Dating
3. Discussion
4. Materials and Methods
4.1. Samples Collection
4.2. Serological Test
4.3. RNA Isolation, RT-PCR, and Sequencing
4.4. Recombination Analysis and Phylogeny
4.5. Population Structure of ToBRFV
4.6. Molecular Dating Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Ethical Approval
Data Availability Statement
Conflicts of Interest
References
- Panno, S.; Caruso, A.G.; Barone, S.; Lo Bosco, G.; Rangel, E.A.; Davino, S. Spread of tomato brown rugose fruit virus in Sicily and evaluation of the spatiotemporal dispersion in experimental conditions. Agronomy 2020, 10, 834. [Google Scholar] [CrossRef]
- Cambrón-Crisantos, J.M.; Rodríguez-Mendoza, J.; Valencia-Luna, J.B.; Alcasio Rangel, S.; de Jesús García-Ávila, C.; López-Buenfil, J.A.; Ochoa-Martínez, D.L. First report of Tomato brown rugose fruit virus (ToBRFV) in Michoacan. Rev. Mex. Fitopatol. 2019, 37, 185–192. [Google Scholar] [CrossRef] [Green Version]
- Abou Kubaa, R.; Choueiri, E.; Heinoun, K.; Cillo, F.; Saponari, M. First report of tomato brown rugose fruit virus infecting sweet pepper in Syria and Lebanon. J. Plant Pathol. 2022, 104, 425. [Google Scholar] [CrossRef]
- King, A.M.; Lefkowitz, E.; Adams, M.J.; Carstens, E.B. Virus Taxonomy: Ninth Report of the International Committee on Taxonomy of Viruses; Elsevier: Amsterdam, The Netherlands, 2011. [Google Scholar]
- Salem, N.; Mansour, A.; Ciuffo, M.; Falk, B.W.; Turina, M. A new tobamovirus infecting tomato crops in Jordan. Arch. Virol. 2016, 161, 503–506. [Google Scholar] [CrossRef] [PubMed]
- Davino, S.; Caruso, A.G.; Bertacca, S.; Barone, S.; Panno, S. Tomato brown rugose fruit virus: Seed transmission rate and efficacy of different seed disinfection treatments. Plants 2020, 9, 1615. [Google Scholar] [CrossRef] [PubMed]
- Oladokun, J.O.; Halabi, M.H.; Barua, P.; Nath, P.D. Tomato brown rugose fruit disease: Current distribution, knowledge and future prospects. Plant Pathol. 2019, 68, 1579–1586. [Google Scholar] [CrossRef] [Green Version]
- Van de Vossenberg, B.T.L.H.; Dawood, T.; Wozny, M.; Botermans, M. First expansion of the public Tomato brown rugose fruit Virus (ToBRFV) Nextstrain Build; Inclusion of New Genomic and Epidemiological Data. PhytoFrontiers 2021, 1, 359–363. [Google Scholar] [CrossRef]
- Luria, N.; Smith, E.; Reingold, V.; Bekelman, I.; Lapidot, M.; Levin, I.; Elad, N.; Tam, Y.; Sela, N.; Abu-Ras, A.; et al. A new Israeli Tobamovirus isolate infects tomato plants harboring Tm-22 resistance genes. PLoS ONE 2017, 12, e0170429. [Google Scholar] [CrossRef] [Green Version]
- Ling, K.S.; Tian, T.; Gurung, S.; Salati, R.; Gilliard, A. First report of tomato brown rugose fruit virus infecting greenhouse tomato in the United States. Plant Dis. 2019, 103, 1439. [Google Scholar] [CrossRef]
- Menzel, W.; Knierim, D.; Winter, S.; Hamacher, J.; Heupel, M. First report of tomato brown rugose fruit virus infecting tomato in Germany. New Dis. Rep. 2019, 39, 1. [Google Scholar] [CrossRef]
- Panno, S.; Caruso, A.G.; Davino, S. First report of tomato brown rugose fruit virus on tomato crops in Italy. Plant Dis. 2019, 103, 1443. [Google Scholar] [CrossRef]
- Alkowni, R.; Alabdallah, O.; Fadda, Z. Molecular identification of tomato brown rugose fruit virus in tomato in Palestine. J. Plant Pathol. 2019, 101, 719–723. [Google Scholar] [CrossRef]
- Fidan, H.; Sarikaya, P.; Calis, O. First report of Tomato brown rugose fruit virus on tomato in Turkey. New Dis. Rep. 2019, 39, 18. [Google Scholar] [CrossRef] [Green Version]
- Skelton, A.; Buxton-Kirk, A.; Ward, R.; Harju, V.; Frew, L.; Fowkes, A.; Long, M.; Negus, A.; Forde, S.; Adams, I.P.; et al. First report of Tomato brown rugose fruit virus in tomato in the United Kingdom. New Dis. Rep. 2019, 40, 12. [Google Scholar] [CrossRef] [Green Version]
- Yan, Z.Y.; Ma, H.Y.; Han, S.L.; Geng, C.; Tian, Y.P.; Li, X.D. First report of Tomato brown rugose fruit virus infecting tomato in China. Plant Dis. 2019, 103, 2973. [Google Scholar] [CrossRef]
- Beris, D.; Malandraki, I.; Kektsidou, O.; Theologidis, I.; Vassilakos, N.; Varveri, C. First report of tomato brown rugose fruit virus infecting tomato in Greece. Plant Dis. 2020, 104, 2035. [Google Scholar] [CrossRef] [Green Version]
- Van de Vossenberg, B.T.L.H.; Visser, M.; Bruinsma, M.; Koenraadt, H.M.; Westenberg, M.; Botermans, M. Real-time tracking of Tomato brown rugose fruit virus (ToBRFV) outbreaks in the Netherlands using Nextstrain. PLoS ONE 2020, 15, e0234671. [Google Scholar] [CrossRef]
- Alfaro-Fernández, A.; Castillo, P.; Sanahuja, E.; Rodríguez-Salido, M.C.; Font, M.I. First report of tomato brown rugose fruit virus in tomato in Spain. Plant Dis. 2021, 105, 515. [Google Scholar] [CrossRef]
- Esmaeilzadeh, F.; Koolivand, D. Occurrence of tomato brown rugose fruit virus in tomato in Iran. J. Plant Pathol. 2022, 104, 457. [Google Scholar] [CrossRef]
- Esmaeilzadeh, F.; Koolivand, D. First report of tomato brown rugose fruit virus infecting bell pepper in Iran. J. Plant Pathol. 2022, 104, 893. [Google Scholar] [CrossRef]
- Skelton, A.; Gentit, P.; Porcher, L.; Visage, M.; Fowkes, A.; Adams, I.P.; Harju, V.; Webster, G.; Pufal, H.; McGreig, S.; et al. First report of Tomato brown rugose fruit virus in tomato in France. New Dis. Rep. 2022, 45, e12061. [Google Scholar] [CrossRef]
- Hanssen, I.M.; Mumford, R.; Blystad, D.R.; Cortez, I.; Hasiów-Jaroszewska, B.; Hristova, D.; Pagán, I.; Pereira, A.M.; Peters, J.; Pospieszny, H.; et al. Seed transmission of Pepino mosaic virus in tomato. Eur. J. Plant Pathol. 2010, 126, 145–152. [Google Scholar] [CrossRef]
- Moya, A.; Holmes, E.C.; González-Candelas, F. The population genetics and evolutionary epidemiology of RNA viruses. Nat. Rev. Microbiol. 2004, 2, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Maayan, Y.; Pandaranayaka, E.P.; Srivastava, D.A.; Lapidot, M.; Levin, I.; Dombrovsky, A.; Harel, A. Using genomic analysis to identify tomato Tm-2 resistance-breaking mutations and their underlying evolutionary path in a new and emerging tobamovirus. Arch. Virol. 2018, 163, 1863–1875. [Google Scholar] [CrossRef]
- Chanda, B.; Rivera, Y.; Nunziata, S.O.; Galvez, M.E.; Gilliard, A.; Ling, K.S. Complete genome sequence of a tomato brown rugose fruit virus isolated in the United States. Microbiol. Resour. Announc. 2020, 9, e00630-20. [Google Scholar] [CrossRef]
- Ma, Z.; Zhang, H.; Ding, M.; Zhang, Z.; Yang, X.; Zhou, X. Molecular characterization and pathogenicity of an infectious cDNA clone of tomato brown rugose fruit virus. Phytopathol. Res. 2021, 3, 14. [Google Scholar] [CrossRef]
- Yan, Z.Y.; Zhao, M.S.; Ma, H.Y.; Liu, L.Z.; Yang, G.L.; Geng, C.; Tian, Y.; Li, X.D. Biological and molecular characterization of tomato brown rugose fruit virus and development of quadruplex RT-PCR detection. J. Integr. Agric. 2021, 20, 1871–1879. [Google Scholar] [CrossRef]
- Zhang, S.; Griffiths, J.S.; Marchand, G.; Bernards, M.A.; Wang, A. Tomato brown rugose fruit virus: An emerging and rapidly spreading plant RNA virus that threatens tomato production worldwide. Mol. Plant Pathol. 2022, 23, 1262–1277. [Google Scholar] [CrossRef]
- Soost, R.K. Hybrid tomato resistant to tobacco mosaic virus: Inheritance for Resistance in Derivatives of a Complex Species Hybrid. J. Hered. 1963, 54, 241–244. [Google Scholar] [CrossRef]
- Alexander, L.J. Transfer of a dominant type of resistance to the four known Ohio pathogenic strains of tobacco mosaic virus (TMV) from Lycopersicon peruvianum to L. esculentum. Phytopathology 1963, 53, 869. [Google Scholar]
- Pfitzner, A.J.P. Resistance to Tobacco mosaic virus and Tomato mosaic virus in tomato. In Natural Resistance Mechanisms of Plants to Viruses; Loebenstein, G., Carr, J.P., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 399–413. [Google Scholar]
- Dombrovsky, A.; Smith, E. Seed transmission of tobamoviruses: Aspects of global disease distribution. In Seed Biology; Jimenez-Lopez, J.C., Ed.; IntechOpen: London, UK, 2017; pp. 234–260. [Google Scholar]
- Fidan, H.; Sarikaya, P.; Yildiz, K.; Topkaya, B.; Eekis, G.; Calis, O. Robust molecular detection of the new Tomato brown rugose fruit virus in infected tomato and pepper plants from Turkey. J. Integr. Agric. 2021, 20, 2170–2179. [Google Scholar] [CrossRef]
- Zinger, A.; Lapidot, M.; Harel, A.; Doron-Faigenboim, A.; Gelbart, D.; Levin, I. Identification and mapping of tomato genome loci controlling tolerance and resistance to tomato brown rugose fruit virus. Plants 2021, 10, 179. [Google Scholar] [CrossRef] [PubMed]
- Jewehan, A.; Salem, N.; Tóth, Z.; Salamon, P.; Szabó, Z. Screening of Solanum (sections Lycopersicon and Juglandifolia) germplasm for reactions to the tomato brown rugose fruit virus (ToBRFV). J. Plant Dis. Prot. 2022, 129, 117–123. [Google Scholar] [CrossRef]
- Morca, A.F.; Çelik, A.; Coşkan, S.; Santosa, A.I.; Akbaş, B. Population analysis on tomato spotted wilt virus isolates inducing various symptoms on tomato, pepper, and Chenopodium album in Turkey. Physiol. Mol. Plant Pathol. 2022, 118, 101786. [Google Scholar] [CrossRef]
- Gibbs, A.J. Tobamovirus classification. In The Plant Viruses; Van Regenmortel, M.H.V., Fraenkel-Conrat, H., Eds.; Springer: Boston, MA, USA, 1986; pp. 167–180. [Google Scholar]
- Duarte, L.M.L.; Alexandre, M.A.V.; Rivas, E.B.; Cattai, M.B.; Soares, R.M.; Harakava, R.; Fernandes, F.M.C. Phylogenetic analysis of Tomato mosaic virus from Hemerocallis sp. and Impatiens hawkeri. Summa Phytopathol. 2007, 33, 409–413. [Google Scholar] [CrossRef]
- Listhihani, L.; Hidayat, S.H.; Wiyono, S.; Damayanti, T.A. Characteristic of Tobacco mosaic virus isolated from cucumber and tobacco collected from East Java, Indonesia. Biodiversitas 2019, 20, 2937–2942. [Google Scholar] [CrossRef]
- Pagán, I.; Firth, C.; Holmes, E.C. Phylogenetic analysis reveals rapid evolutionary dynamics in the plant RNA virus genus Tobamovirus. J. Mol. Evol. 2010, 71, 298–307. [Google Scholar] [CrossRef]
- Stobbe, A.H.; Melcher, U.; Palmer, M.W.; Roossinck, M.J.; Shen, G. Co-divergence and host-switching in the evolution of tobamoviruses. J. Gen. Virol. 2012, 93, 408–418. [Google Scholar] [CrossRef]
- Gibbs, A.J.; Wood, J.; Garcia-Arenal, F.; Ohshima, K.; Armstrong, J.S. Tobamoviruses have probably co-diverged with their eudicotyledonous hosts for at least 110 million years. Virus Evol. 2015, 1, vev019. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 22, 1547–1549. [Google Scholar] [CrossRef]
- Martin, D.P.; Murrell, B.; Golden, M.; Khoosal, A.; Muhire, B. RDP4: Detection and analysis of recombination patterns in virus genomes. Virus Evol. 2015, 1, vev003. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K. Estimation of the number of nucleotide substitutions when there are strong transition-transversion and G + C-content biases. Mol. Biol. Evol. 1992, 9, 678–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muhire, B.M.; Varsani, A.; Martin, D.P. (SDT: A virus classification tool based on pairwise sequence alignment and identity calculation. PLoS ONE 2014, 9, e108277. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Datamonkey. Available online: http://www.datamonkey.org (accessed on 25 October 2022).
- Fu, Y.X.; Li, W.H. Statistical tests of neutrality of mutations. Genetics 1993, 133, 693–709. [Google Scholar] [CrossRef] [PubMed]
- Tajima, F. Statistical method for testing the neutral mutation hypothesis by DNA polymorphism. Genetics 1989, 123, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Hudson, R.R. A new statistic for detecting genetic differentiation. Genetics 2000, 155, 2011–2014. [Google Scholar] [CrossRef] [PubMed]
- Çelik, A.; Santosa, A.I.; Gibbs, A.J.; Ertunç, F. Prunus necrotic ringspot virus in Turkey: An immigrant population. Arch. Virol. 2022, 167, 553–562. [Google Scholar] [CrossRef] [PubMed]
- Coşkan, S.; Morca, A.F.; Akbaş, B.; Çelik, A.; Santosa, A.I. Comprehensive surveillance and population study on plum pox virus in Ankara Province of Turkey. J. Plant Dis. Prot. 2022, 129, 981–991. [Google Scholar] [CrossRef]
- Mello, B. Estimating TimeTrees with MEGA and the TimeTree resource. Mol. Biol. Evol. 2018, 35, 2334–2342. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population | N | h | Hd | S | η | k | π | dS | dN | ω |
|---|---|---|---|---|---|---|---|---|---|---|
| Complete genome | 174 | 123 | 0.994 | 377 | 390 | 17.396 | 0.0028 | 0.0088 | 0.0011 | 0.125 |
| Phylogroup | ||||||||||
| Group 1 | 49 | 35 | 0.985 | 62 | 63 | 5.639 | 0.0009 | 0.0029 | 0.0004 | 0.1379 |
| Group 2 | 124 | 87 | 0.991 | 326 | 333 | 17.719 | 0.0029 | 0.0087 | 0.0012 | 0.1379 |
| Region 1 | ||||||||||
| Europe | 130 | 86 | 0.991 | 235 | 241 | 17.372 | 0.0028 | 0.0089 | 0.0011 | 0.1236 |
| ME and China | 34 | 28 | 0.988 | 132 | 134 | 12.882 | 0.0021 | 0.0059 | 0.0009 | 0.1525 |
| America | 10 | 10 | 1.000 | 49 | 50 | 14.667 | 0.0024 | 0.0081 | 0.0008 | 0.0988 |
| ORF1 | 174 | 112 | 0.989 | 283 | 292 | 12.789 | 0.0026 | 0.0085 | 0.0009 | 0.1059 |
| ORF2 | 174 | 103 | 0.984 | 204 | 205 | 8.236 | 0.0025 | 0.0081 | 0.0008 | 0.0988 |
| ORF3 (MP) | 174 | 52 | 0.933 | 60 | 63 | 3.674 | 0.0046 | 0.0121 | 0.0025 | 0.2066 |
| ORF4 (CP) | 185 | 34 | 0.646 | 39 | 40 | 0.994 | 0.0021 | 0.0069 | 0.0006 | 0.0869 |
| Phylogroup | ||||||||||
| Group 1 | 49 | 7 | 0.267 | 7 | 7 | 0.325 | 0.0007 | 0.0025 | 0.0001 | 0.04 |
| Group 2 | 134 | 25 | 0. 421 | 29 | 29 | 0.637 | 0.0013 | 0.0035 | 0.0007 | 0.2 |
| Region 1 | ||||||||||
| Europe | 131 | 21 | 0.668 | 23 | 24 | 0.987 | 0.0021 | 0.0072 | 0.0005 | 0.0694 |
| ME and China | 42 | 10 | 0.458 | 12 | 12 | 0.707 | 0.0015 | 0.0033 | 0.0009 | 0.2727 |
| America | 12 | 5 | 0.576 | 7 | 7 | 1.167 | 0.0024 | 0.0088 | 0.0005 | 0.0568 |
| Population | Fu and Li’s D * | Fu and Li’s F * | Tajima’s D |
|---|---|---|---|
| Complete genome | −5.56444 ** | −4.81135 ** | −2.40531 ** |
| Phylogroup | |||
| Group 1 | −2.18556 ns | −2.57257 * | −2.10472 * |
| Group 2 | −4.89980 ** | −4.48389 ** | −2.36401 ** |
| Region 1 | |||
| Europe | −3.60441 ** | −3.43963 ** | −1.99775 * |
| ME and China | −2.48265 ns | −2.86841 * | −2.28997 ** |
| America | −1.10835 ns | −1.17368 ns | −0.83168 ns |
| ORF1 | −5.33498 ** | −4.69009 ** | −2.41119 ** |
| ORF2 | −5.63445 ** | −4.93305 ** | −2.46251 ** |
| ORF3 (MP) | −3.27960 ** | −3.27892 ** | −2.03581 * |
| ORF4 (CP) | −6.29855 ** | −5.63807 ** | −2.51446 *** |
| Phylogroup | |||
| Group 1 | −3.41067 ** | −3.51604 ** | −2.10661 * |
| Group 2 | −5.37507 ** | −5.10880 ** | −2.58167 *** |
| Region 1 | |||
| Europe | −4.81828 ** | −4.56208 ** | −2.23084 ** |
| ME and China | −3.17348 * | −3.39447 ** | −2.27783 ** |
| America | −2.38877 * | −2.57986 * | −1.94368 * |
| Comparison | αKS* | αKST * | p-Value | αZ * | p-Value | Snn | p-Value | βFST |
|---|---|---|---|---|---|---|---|---|
| Complete genome | ||||||||
| Phylogroup | ||||||||
| All (n = 174)/Group 1(n = 49) | 2.5781 | 0.0421 | 0.0000 *** | 8.9495 | 0.0000 *** | 0.6405 | 0.7180 ns | 0.2551 |
| All (n = 174)/Group 2 (n = 124) | 2.7892 | 0.0066 | 0.0000 *** | 9.6674 | 0.0000 *** | 0.3144 | 1.0000 ns | 0.0281 |
| Group 1 (n = 49)/Group 2 (n = 124) | 2.5164 | 0.0988 | 0.0000 *** | 8.1317 | 0.0000 *** | 1.0000 | 0.0000 *** | 0.3987 |
| Region 1 | ||||||||
| Europe (n = 130)/ME and China (n = 34) | 2.6901 | 0.0279 | 0.0000 *** | 8.3888 | 0.0000 *** | 0.9415 | 0.0000 *** | 0.1291 |
| Europe (n = 130)/America (n = 10) | 2.7292 | 0.0151 | 0.0000 *** | 8.1349 | 0.0000 *** | 0.9929 | 0.0000 *** | 0.1836 |
| ME and China (n = 34)/America (n = 10) | 2.5216 | 0.0295 | 0.0000 *** | 5.7025 | 0.0000 *** | 0.9886 | 0.0000 *** | 0.1293 |
| ORF4 (CP) | ||||||||
| Phylogroup | ||||||||
| All (n = 185)/Group 1 (n = 49) | 0.4993 | 0.1687 | 0.0000 *** | 9.1645 | 0.0000 *** | 0.7642 | 0.0000 *** | 0.4498 |
| All (n = 185)/Group 2 (n = 134) | 0.4899 | 0.0369 | 0.0000 *** | 9.9451 | 0.0000 *** | 0.5081 | 0.6070 ns | 0.0748 |
| Group 1 (n = 49)/Group 2 (n = 134) | 0.3298 | 0.4091 | 0.0000 *** | 8.5045 | 0.0000 *** | 1.0000 | 0.0000 *** | 0.6758 |
| Region 1 | ||||||||
| Europe (n = 131)/ME and China (n = 42) | 0.5396 | 0.0494 | 0.0000 *** | 8.6861 | 0.0000 *** | 0.7149 | 0.0000 *** | 0.1451 |
| Europe (n = 131)/America (n = 12) | 0.5807 | 0.0175 | 0.0120 * | 8.3262 | 0.0220 * | 0.8674 | 0.0210 * | 0.1124 |
| ME and China (n = 42)/America (n = 12) | 0.4511 | 0.0065 | 0.2500 ns | 6.4269 | 0,2870 ns | 0.6580 | 0.3070 ns | −0.0005 |
| ToBRFV vs. | Identity (%) | |
|---|---|---|
| Nucleotide | Amino Acid | |
| PMMoV | 69–69.6 | 61.8–62.3 |
| RehMV | 80.1–81.5 | 84–85.9 |
| TMV | 81.2–81.7 | 85.3–86.7 |
| ToMV | 81–81.4 | 85.5–86.4 |
| ToMMV | 80.7–81–1 | 84.4–85.5 |
| BPMV | 75.4–76.1 | 71.4–72.1 |
| YTMMV | 64.1–64.5 | 57.9–58.4 |
| TMGMV | 65.1–65.5 | 62.5–63.1 |
| ObPV | 65–65.1 | 62.9–64 |
| PaMMV | 65.3–65.7 | 63.7–64.1 |
| Species | Mean Patristic Distance | Ratios of ToBRFV and Other Patristic Distances |
|---|---|---|
| PaMMV | 0.68 | 4.53 |
| ObPV | 0.68 | 4.53 |
| TMGMV | 0.53 | 3.53 |
| YTMMV | 0.45 | 3 |
| BPMV | 0.28 | 1.87 |
| ToBRFV | 0.15 | 1 |
| ToMMV | 0.12 | 0.8 |
| ToMV | 0.12 | 0.8 |
| TMV | 0.11 | 0.73 |
| RehMV | 0.10 | 0.67 |
| PMMoV | 0.10 | 0.67 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Çelik, A.; Coşkan, S.; Morca, A.F.; Santosa, A.I.; Koolivand, D. Insight into Population Structure and Evolutionary Analysis of the Emerging Tomato Brown Rugose Fruit Virus. Plants 2022, 11, 3279. https://doi.org/10.3390/plants11233279
Çelik A, Coşkan S, Morca AF, Santosa AI, Koolivand D. Insight into Population Structure and Evolutionary Analysis of the Emerging Tomato Brown Rugose Fruit Virus. Plants. 2022; 11(23):3279. https://doi.org/10.3390/plants11233279
Chicago/Turabian StyleÇelik, Ali, Sevgi Coşkan, Ali Ferhan Morca, Adyatma Irawan Santosa, and Davoud Koolivand. 2022. "Insight into Population Structure and Evolutionary Analysis of the Emerging Tomato Brown Rugose Fruit Virus" Plants 11, no. 23: 3279. https://doi.org/10.3390/plants11233279