
Transcriptome Analysis of Genes Involved in Fatty Acid and Lipid Biosynthesis in Developing Walnut (Juglans regia L.) Seed Kernels from Qinghai Plateau

Abstract
:1. Introduction
2. Results
2.1. Morphological Characteristics and Oil Accumulation during J. regia Embryo Development
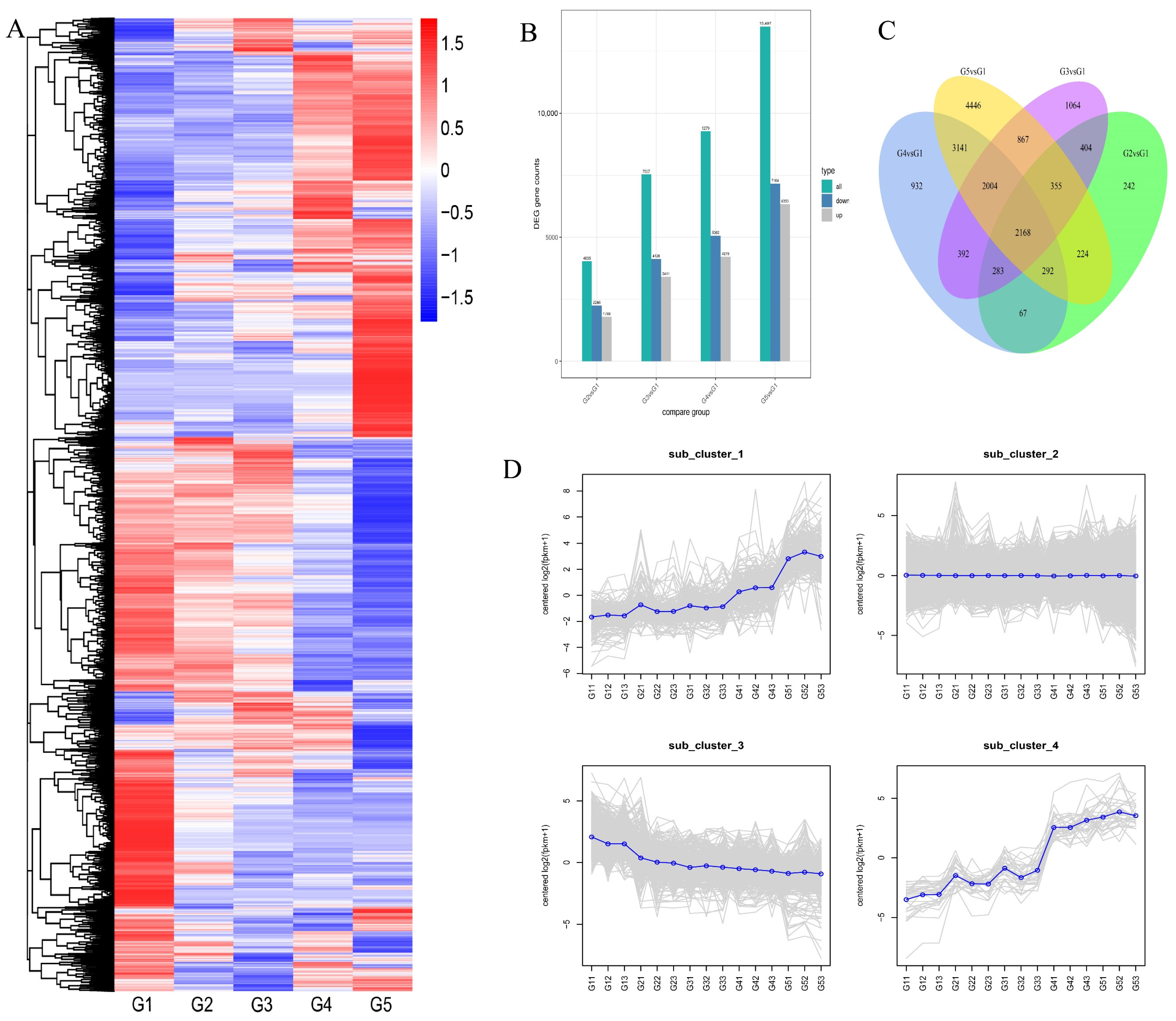
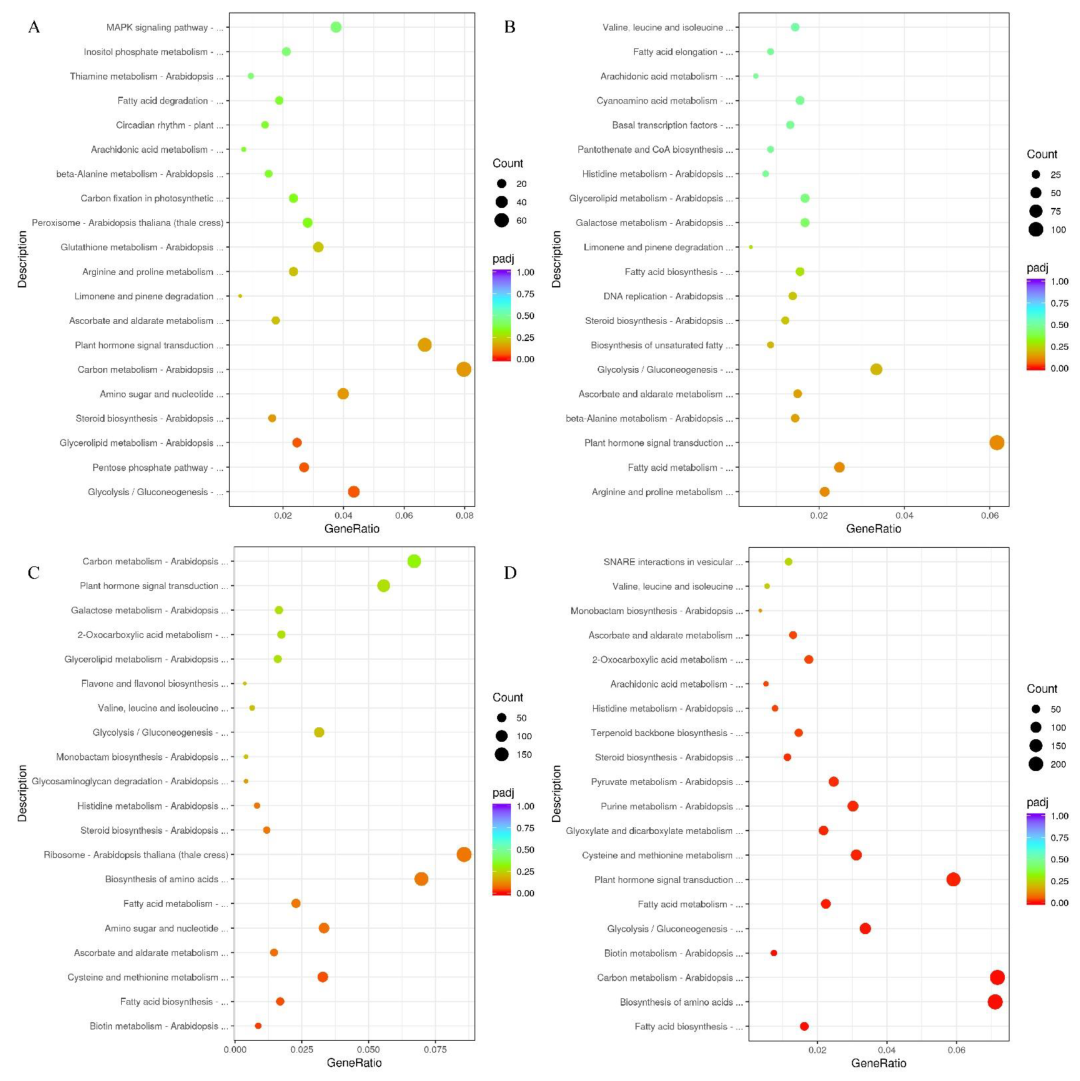
2.2. Identification of DEGs and Enrichment Analysis during Seed Development
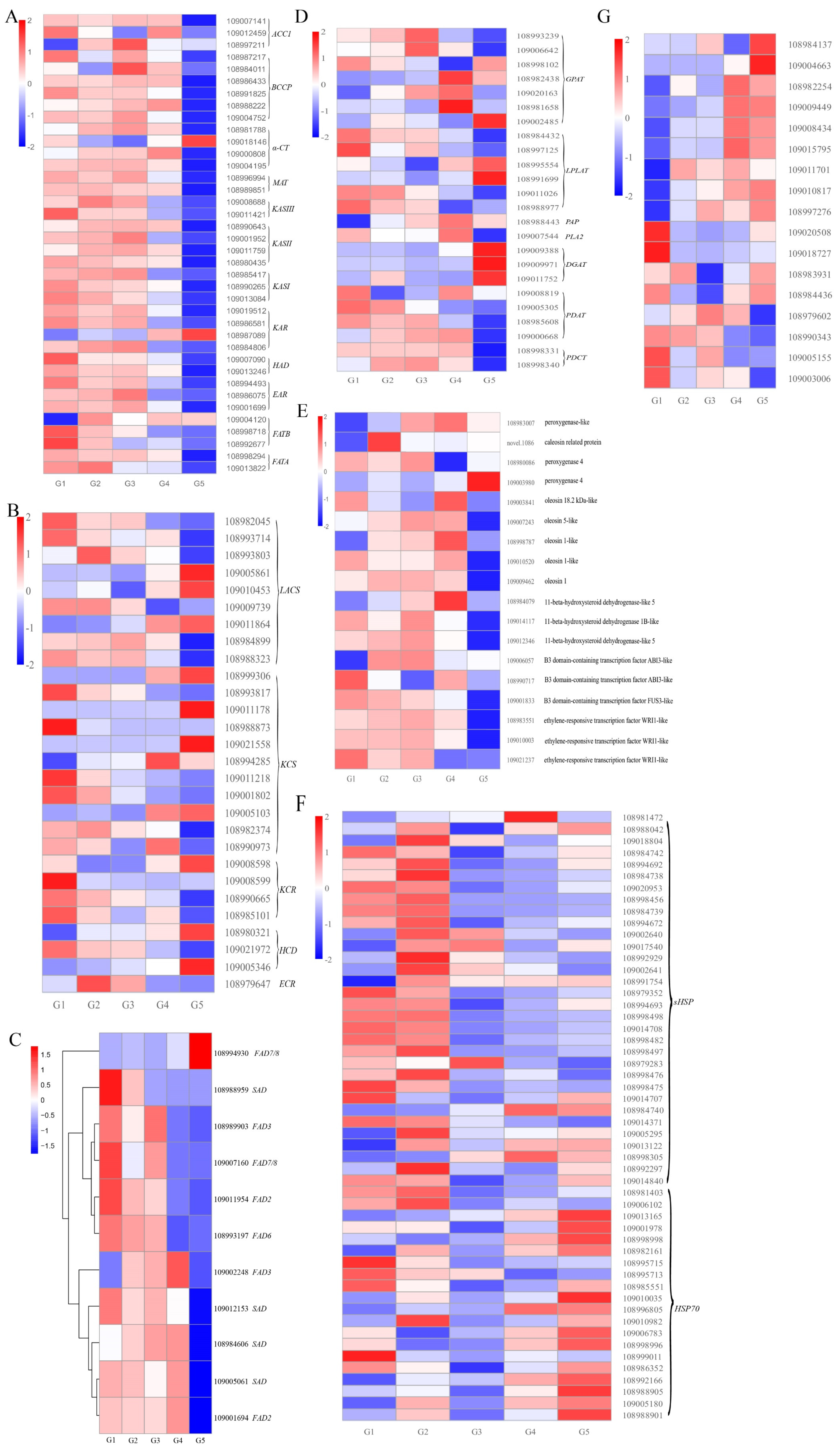
2.3. Analysis of DEGs for Lipid Metabolism
2.4. Identification and Expression Profiling of DEGs for Fatty Acid Biosynthesis, Elongation, and Desaturation
2.5. Identification and Expression Profiling of DEGs for TAG Assembly and Oil Accumulation
2.6. Identification and Expression Profiling of DEGs of HSPs and HSFs
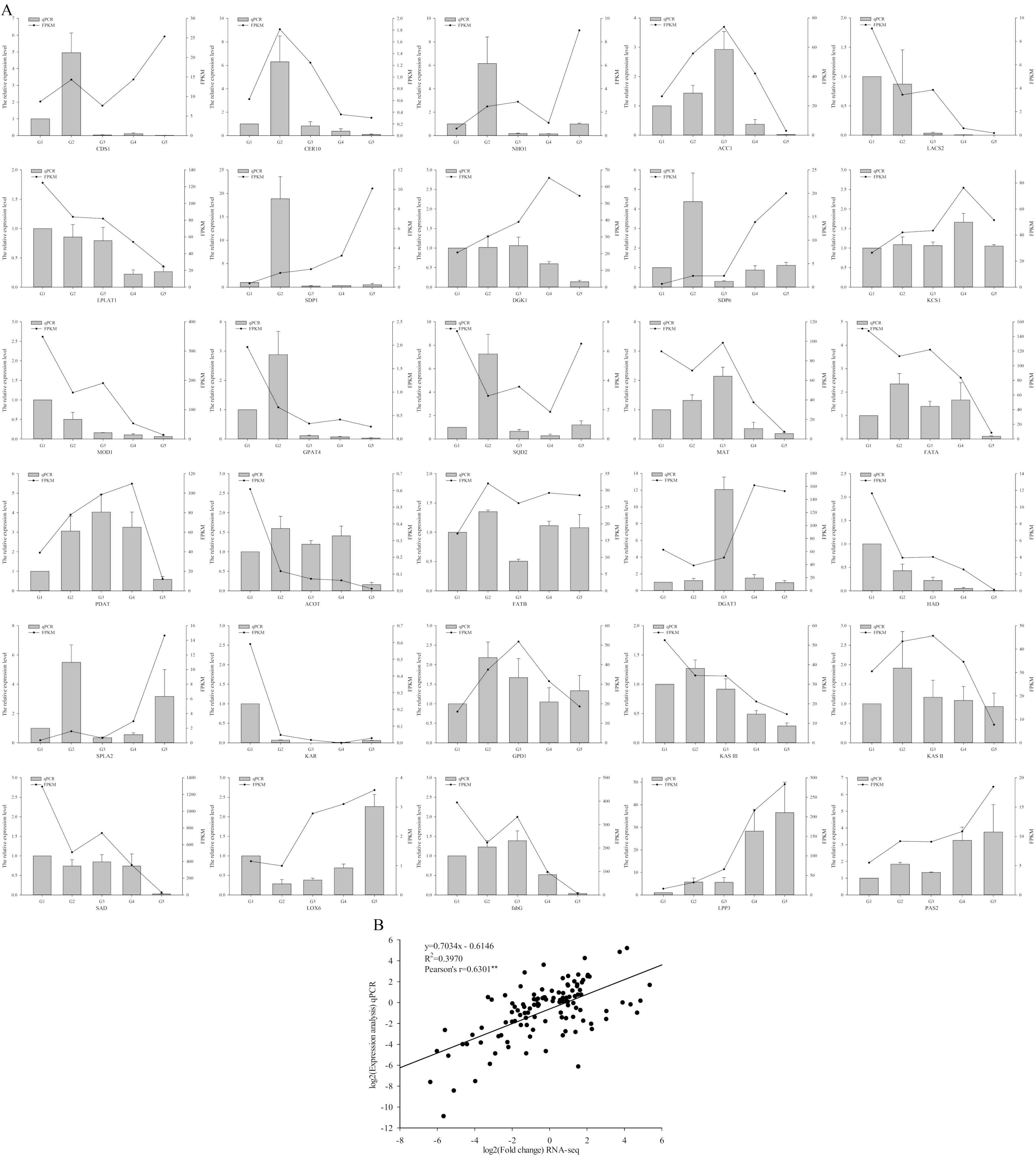
2.7. Validation of RNA-Seq Results by Quantitative Real-Time PCR (qRT-PCR)
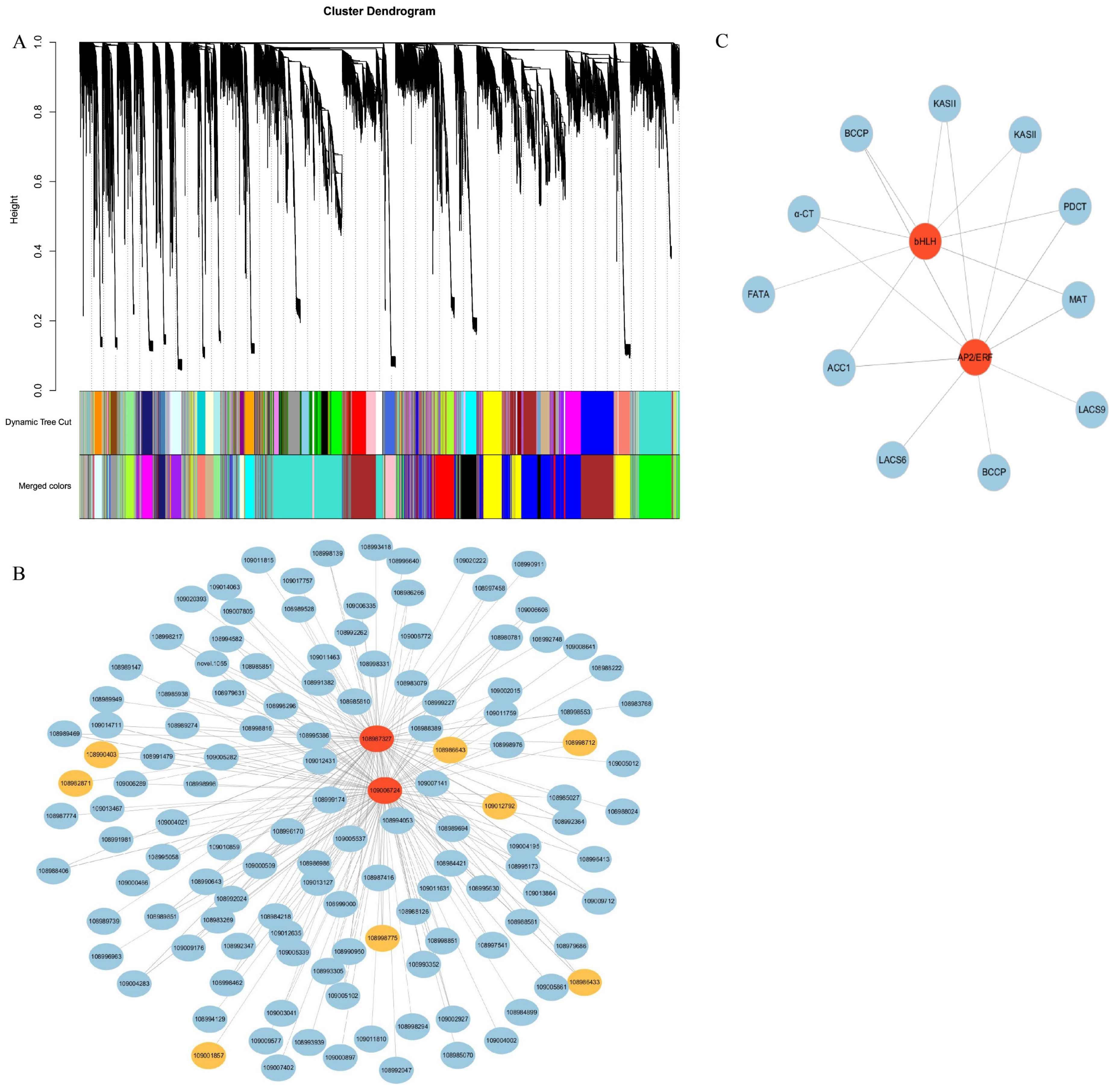
2.8. Weighted Gene Co-Expression Network Analysis
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Sampling and Fruit Growth Analysis
4.3. Kernel Oil Content and Fatty Acid Composition Detection
4.4. Total RNA Isolation, Transcriptome Sequencing Library Construction, and the Next Generation Sequencing
4.5. Analysis of Transcriptome, Quality Control and Clean Reads Mapping to the Reference Genome
4.6. Quantification of Gene Expression Level and Differential Expression Analysis
4.7. GO and KEGG Enrichment of DEGs
4.8. WGCNA
4.9. Quantitative Real-Time PCR and Correlation Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abdallah, I.B.; Tlili, N.; Martinez-Force, E.; Rubio, A.G.P.; Perez-Camino, M.C.; Albouchi, A.; Boukhchina, S. Content of carotenoids, tocopherols, sterols, triterpenic and aliphatic alcohols, and volatile compounds in six walnuts (Juglans regia L.) varieties. Food Chem. 2015, 173, 972–978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouabdallah, I.; Bouali, I.; Martinez-Force, E.; Albouchi, A.; Perez Camino, M.C.; Boukhchina, S. Composition of fatty acids, triacylglycerols and polar compounds of different walnut varieties (Juglans regia L.) from Tunisia. Nat. Prod. Res. 2014, 28, 1826–1833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruijun, W.; Shi, W.; Yijun, X.; Mengwuliji, T.; Lijuan, Z.; Yumin, W. Antitumor effects and immune regulation activities of a purified polysaccharide extracted from Juglan regia. Int. J. Biol. Macromol. 2015, 72, 771–775. [Google Scholar] [CrossRef] [PubMed]
- Djoussé, L.; Lu, B.; Gaziano, J.M. Effects of walnut consumption on endothelial function in people with type 2 diabetes: A randomized pilot trial. Curr. Nutr. Rep. 2016, 5, 1–8. [Google Scholar] [CrossRef]
- Kendall, C.W.C.; Esfahani, A.; Josse, A.R.; Augustin, L.S.A.; Vidgen, E.; Jenkins, D.J.A. The glycemic effect of nut-enriched meals in healthy and diabetic subjects. Nutr. Metab. Cardiovasc. Dis. 2011, 21, S34–S39. [Google Scholar] [CrossRef]
- Tindall, A.M.; Mclimans, C.J.; Petersen, K.S.; Kris-Etherton, P.M.; Lamendella, R. Walnuts and vegetable oils containing oleic acid differentially affect the gut microbiota and associations with cardiovascular risk factors: Follow-up of a randomized, controlled, feeding trial in adults at risk for cardiovascular disease. J. Nutr. 2020, 150, 806–817. [Google Scholar] [CrossRef] [Green Version]
- Martínez, M.L.; Mattea, M.A.; Maestri, D.M. Varietal and crop year effects on lipid composition of walnut (Juglans regia) genotypes. J. Am. Oil Chem. Soc. 2006, 83, 791–796. [Google Scholar] [CrossRef]
- Amaral, J.S.; Casal, S.; Pereira, J.A.; Seabra, R.M.; Oliveira, B.P.P. Determination of sterol and fatty acid compositions, oxidative stability, and nutritional value of six walnut (Juglans regia L.) cultivars grown in Portugal. J. Agr. Food Chem. 2003, 51, 7698–7702. [Google Scholar] [CrossRef] [Green Version]
- Prasad, R.B.N. Walnuts and pecans. In Encyclopedia of Food Sciences and Nutrition, 2nd ed.; Caballero, B., Ed.; Academic Press: Oxford, UK, 2003; pp. 6071–6079. ISBN 978-0-12-227055-0. [Google Scholar]
- Li, Q.; Yin, R.; Zhang, Q.; Wang, X.; Hu, X.; Gao, Z.; Duan, Z. Chemometrics analysis on the content of fatty acid compositions in different walnut (Juglans regia L.) varieties. Eur. Food Res. Technol. 2017, 243, 2235–2242. [Google Scholar] [CrossRef]
- Zwarts, L.; Savage, G.P.; Mcneil, D.L. Fatty acid content of New Zealand-grown walnuts (Juglans regia L.). Int. J. Food Sci. Nutr. 1999, 50, 189–194. [Google Scholar] [CrossRef]
- Doğan, A.; Celik, F.; Balta, F.; Javidipour, I.; Yaviç, A. Analysis of fatty acid profiles of pistachios (Pistacia vera L.) and native walnuts (Juglans regia L.) from Turkey. Asian J. Chem. 2010, 22, 517–521. [Google Scholar]
- Ozrenk, K.; Javidipour, I.; Yarilgac, T.; Balta, F.; Gundogdu, M. Fatty acids, tocopherols, selenium and total carotene of pistachios (P. vera L.) from Diyarbakir (Southeastern Turkey) and walnuts (J. regia L.) from Erzincan (Eastern Turkey). Food Sci. Technol. Int. 2012, 18, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Zhai, M.Z.; Wang, D.; Tao, X.D.; Wang, Z.Y. Fatty acid compositions and tocopherol concentrations in the oils of 11 varieties of walnut (Juglans regia L.) grown at Xinjiang, China. J. Hortic. Sci. Biotechnol. 2015, 90, 715–718. [Google Scholar] [CrossRef]
- Gao, P.; Jin, J.; Liu, R.; Jin, Q.; Wang, X. Chemical compositions of walnut (Juglans regia L.) oils from different cultivated regions in China. J. Am. Oil Chem. Soc. 2018, 95, 825–834. [Google Scholar] [CrossRef]
- Poggetti, L.; Ferfuia, C.; Chiabà, C.; Testolin, R.; Baldini, M. Kernel oil content and oil composition in walnut (Juglans regia L.) accessions from north-eastern Italy. J. Sci. Food Agr. 2018, 98, 955–962. [Google Scholar] [CrossRef] [Green Version]
- Gao, P.; Liu, R.; Jin, Q.; Wang, X. Comparative study of chemical compositions and antioxidant capacities of oils obtained from two species of walnut: Juglans regia and Juglans sigillata. Food Chem. 2019, 279, 279–287. [Google Scholar] [CrossRef]
- Combres, J.; Pallas, B.; Rouan, L.; Mialet-Serra, I.; Caliman, J.; Braconnier, S.; Soulié, J.; Dingkuhn, M. Simulation of inflorescence dynamics in oil palm and estimation of environment-sensitive phenological phases: A model based analysis. Funct. Plant Biol. FPB 2013, 40, 263–279. [Google Scholar] [CrossRef]
- Shen, Q.; Zhao, J.; Qiu, X.; Shang, Z.; Wang, X.; Yang, S.; Xu, J.; Chen, S. Research on influence of environment factors to yield and quality traits of Perilla frutescen. China J. Chin. Mater. Med. 2018, 43, 4033–4043. [Google Scholar] [CrossRef]
- Dabbou, S.; Sifi, S.; Rjiba, I.; Esposto, S.; Taticchi, A.; Servili, M.; Montedoro, G.F.; Hammami, M. Effect of pedoclimatic conditions on the chemical composition of the sigoise olive cultivar. Chem. Biodivers. 2010, 7, 898–908. [Google Scholar] [CrossRef]
- Primomo, V.S.; Falk, D.E.; Ablett, G.R.; Tanner, J.W.; Rajcan, I. Genotype × environment interactions, stability, and agronomic performance of soybean with altered fatty acid profiles. Crop Sci. 2002, 42, 37–44. [Google Scholar] [CrossRef]
- Lee, J.; Woolard, M.; Sleper, D.A.; Smith, J.R.; Pantalone, V.R.; Nyinyi, C.N.; Cardinal, A.; Shannon, J.G. Environmental effects on oleic acid in soybean seed oil of plant introductions with elevated oleic concentration. Crop Sci. 2009, 49, 1762–1768. [Google Scholar] [CrossRef]
- Greve, L.; Mcgranahan, G.; Hasey, J.; Snyder, R.; Kelly, K.; Goldhamer, D.; Labavitch, J. Variation in polyunsaturated fatty acids composition of persian walnut. J. Am. Soc. Hortic. Sci. 1992, 117, 518–522. [Google Scholar] [CrossRef]
- Tzen, J.; Cao, Y.; Laurent, P.; Ratnayake, C.; Huang, A. Lipids, proteins, and structure of seed oil bodies from diverse species. Plant Physiol. 1993, 101, 267–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bates, P.D.; Stymne, S.; Ohlrogge, J. Biochemical pathways in seed oil synthesis. Curr. Opin. Plant Biol. 2013, 16, 358–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, S.; Wang, X.; Yang, C.; Wang, J.; Wang, Y.; Wu, B.; Qiao, L.; Zhao, J.; Mohammad, P.; Zheng, X.; et al. Insights into walnut lipid metabolism from metabolome and transcriptome analysis. Front. Genet. 2021, 12, 1334. [Google Scholar] [CrossRef]
- Zhao, S.; Zhang, X.; Su, Y.; Chen, Y.; Liu, Y.; Sun, M.; Qi, G.; Ferrante, A. Transcriptome analysis reveals dynamic fat accumulation in the walnut kernel. Int. J. Genom. 2018, 2018, 1–13. [Google Scholar] [CrossRef]
- Zhao, X.; Yang, G.; Liu, X.; Yu, Z.; Peng, S. Integrated analysis of seed microRNA and mRNA transcriptome reveals important functional genes and microRNA-targets in the process of walnut (Juglans regia) seed oil accumulation. Int. J. Mol. Sci. 2020, 21, 9093. [Google Scholar] [CrossRef]
- Huang, R.; Zhou, Y.; Zhang, J.; Ji, F.; Pei, D. Transcriptome analysis of walnut (Juglans regia L.) embryos reveals key developmental stages and genes involved in lipid biosynthesis and polyunsaturated fatty acid metabolism. J. Agr. Food Chem. 2021, 69, 377–396. [Google Scholar] [CrossRef]
- Arondel, V.; Lemieux, B.; Hwang, I.; Gibson, S.; Goodman, H.; Somerville, C. Map-based cloning of a gene controlling omega-3 fatty acid desaturation in Arabidopsis. Science 1992, 258, 1353–1355. [Google Scholar] [CrossRef] [PubMed]
- Okuley, J.; Lightner, J.; Feldmann, K.; Yadav, N. Arabidopsis FAD2 gene encodes the enzyme that is essential for polyunsaturated lipid synthesis. Plant Cell 1994, 6, 147–158. [Google Scholar]
- Baud, S.; Lepiniec, L. Physiological and developmental regulation of seed oil production. Prog. Lipid Res. 2010, 49, 235–249. [Google Scholar] [CrossRef] [PubMed]
- St Hl, U.; St Lberg, K.; Stymne, S.; Ronne, H. A family of eukaryotic lysophospholipid acyltransferases with broad specificity. Febs Lett. 2008, 582, 305–309. [Google Scholar] [CrossRef] [PubMed]
- Bates, P.D.; Fatihi, A.; Snapp, A.R.; Carlsson, A.S.; Browse, J.; Lu, C. Acyl editing and headgroup exchange are the major mechanisms that direct polyunsaturated fatty acid flux into triacylglycerols. Plant Physiol. 2012, 160, 1530–1539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jasieniecka-Gazarkiewicz, K.; Demski, K.; Lager, I.; Stymne, S.; Banaś, A. Possible role of different yeast and plant lysophospholipid: Acyl-CoA acyltransferases (LPLATs) in acyl remodelling of phospholipids. Lipids 2016, 51, 15–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Pan, C.; Wang, B.; Xiao, Z.; Hu, Y.; Hu, G. Oil body observation in seed development and its analysis in seed of Juglans regia ‘Wen185’ and J. regia ‘Xinxin2’ in period of seed maturity. Sci. Agric. Sincia 2015, 48, 3899–3909. [Google Scholar]
- Chen, H.; Pan, C.; Wang, B.; Hu, Y.; Xiao, Z.; He, M. The relationship among nutrients’ accumulation and dynamic changes of fatty acids in seed development of walnut. J. Agric. Univ. Hebei 2016, 39, 57–62. [Google Scholar]
- Simsek, M. Chemical, mineral, and fatty acid compositions of various types of walnut (Juglans regia L.) in Turkey. Izv. Po Khimiya Bulg. Akad. Na Nauk. 2016, 48, 66–70. [Google Scholar]
- Geng, S.; Ning, D.; Ma, T.; Chen, H.; Zhang, Y.; Sun, X. Comprehensive analysis of the components of walnut kernel (Juglans regia L.) in China. J. Food Qual. 2021, 2021, 1–11. [Google Scholar] [CrossRef]
- Hao, B.Q.; Zhao, Z.H.; Cai, Y.; Wang, L.H.; Li, M.S.; Rui, Y.K. Effects of different environmental conditions on composition of walnut oil. Fresen. Environ. Bull. 2021, 30, 3130–3133. [Google Scholar]
- Hou, G.; Ablett, G.R.; Pauls, K.P.; Rajcan, I. Environmental effects on fatty acid levels in soybean seed oil. J. Am. Oil Chem. Soc. 2006, 83, 759–763. [Google Scholar] [CrossRef]
- Lin, P.; Wang, K.; Zhou, C.; Xie, Y.; Yao, X.; Yin, H. Seed transcriptomics analysis in Camellia oleifera uncovers genes associated with oil content and fatty acid composition. Int. J. Mol. Sci. 2018, 19, 118. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Zhang, T.; Zhang, Q.; Chen, M.; Wang, Z.; Zheng, B.; Xia, G.; Yang, X.; Huang, C.; Huang, Y. The mechanism of high contents of oil and oleic acid revealed by transcriptomic and lipidomic analysis during embryogenesis in Carya cathayensis Sarg. BMC Genom. 2016, 17, 113. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Wang, Y.; Huang, Y.; Cui, Y.; Hua, J. Gene network of oil accumulation reveals expression profiles in developing embryos and fatty acid composition in upland cotton. J. Plant Physiol. 2018, 228, 101–112. [Google Scholar] [CrossRef]
- Nan, S.; Zhang, L.; Hu, X.; Miao, X.; Han, X.; Fu, H. Transcriptomic analysis reveals key genes involved in oil and linoleic acid biosynthesis during Artemisia sphaerocephala seed development. In. J. Mol. Sci. 2021, 22, 8369. [Google Scholar] [CrossRef]
- Yu, S.; Zhang, X.; Huang, L.; Lyu, Y.; Zhang, Y.; Yao, Z.; Zhang, X.; Yuan, J.; Hu, Y. Transcriptomic analysis of α-linolenic acid content and biosynthesis in Paeonia ostii fruits and seeds. BMC Genom. 2021, 22, 297. [Google Scholar] [CrossRef]
- Banik, M.; Duguid, S.; Cloutier, S. Transcript profiling and gene characterization of three fatty acid desaturase genes in high, moderate, and low linolenic acid genotypes of flax (Linum usitatissimum L.) and their role in linolenic acid accumulation. Genome 2011, 54, 471–483. [Google Scholar] [CrossRef]
- Liao, B.N.; Hao, Y.J.; Lu, J.X.; Bai, H.Y.; Guan, L.; Zhang, T. Transcriptomic analysis of Perilla frutescens seed to insight into the biosynthesis and metabolic of unsaturated fatty acids. BMC Genom. 2018, 19, 213. [Google Scholar] [CrossRef] [Green Version]
- Banaś, W.; Sanchez Garcia, A.; Banaś, A.; Stymne, S. Activities of acyl-CoA:diacylglycerol acyltransferase (DGAT) and phospholipid:diacylglycerol acyltransferase (PDAT) in microsomal preparations of developing sunflower and safflower seeds. Planta 2013, 237, 1627–1636. [Google Scholar] [CrossRef] [Green Version]
- Ding, M.; Lou, H.; Chen, W.; Zhou, Y.; Zhang, Z.; Xiao, M.; Wang, Z.; Yang, Y.; Yang, L.; Zhang, F.; et al. Comparative transcriptome analysis of the genes involved in lipid biosynthesis pathway and regulation of oil body formation in Torreya grandis kernels. Ind. Crop. Prod. 2020, 145, 112051. [Google Scholar] [CrossRef]
- Woodfield, H.K.; Cazenave-Gassiot, A.; Haslam, R.P.; Guschina, I.A.; Wenk, M.R.; Harwood, J.L. Using lipidomics to reveal details of lipid accumulation in developing seeds from oilseed rape (Brassica napus L.). Biochim. Et Biophys. Acta (BBA)—Mol. Cell Biol. Lipids 2018, 1863, 339–348. [Google Scholar] [CrossRef]
- Meng, J.; Tang, Y.; Sun, J.; Zhao, D.; Zhang, K.; Tao, J. Identification of genes associated with the biosynthesis of unsaturated fatty acid and oil accumulation in herbaceous peony ‘Hangshao’ (Paeonia lactiflora ‘Hangshao’) seeds based on transcriptome analysis. BMC Genom. 2021, 22, 94. [Google Scholar] [CrossRef] [PubMed]
- Siloto, R.M.P.; Findlay, K.; Lopez-Villalobos, A.; Yeung, E.C.; Nykiforuk, C.L.; Moloney, M.M. The accumulation of olesins determines the size of seed oilbodies in Arabidopsis. Plant Cell 2006, 18, 1961–1974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Wen, H.; Jin, Q.; Yu, W.; Li, G.; Wu, M.; Bai, H.; Shen, L.; Wu, C. Comparative transcriptome analysis on candidate genes involved in lipid biosynthesis of developing kernels for three walnut cultivars in Xinjiang. Food Sci. Hum. Wellness 2022, 11, 1201–1214. [Google Scholar] [CrossRef]
- Mortazavi, A.; Williams, B.A.; Mccue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bray, N.L.; Pimentel, H.; Melsted, P.; Pachter, L. Near-optimal probabilistic RNA-seq quantification. Nat. Biotechnol. 2016, 34, 525–527. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Society. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Yu, G.; Wang, L.; Han, Y.; He, Q. ClusterProfiler: An R package for comparing biological themes among gene clusters. OMICS: A J. Integr. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using Real-Time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acid Type | Content | ||
|---|---|---|---|
| Saturated (SFA) | Myristic acid (14:0) | 0.0220 ± 0.0007 | |
| Palmitic acid (16:0) | 6.4067 ± 0.1109 | ||
| Heptadecanoic acid (17:0) | 0.0539 ± 0.0007 | ||
| Stearic acid (18:0) | 2.0400 ± 0.0829 | ||
| Arachidic acid (20:0) | 0.0863 ± 0.0075 | ||
| Docosanoic acid (22:0) | 0.0429 ± 0.0069 | ||
| Tetracosanoic acid (24:0) | 0.0195 ± 0.0138 | ||
| Unsaturated (UFA) | Monounsaturated (MUFA) | Palmitoleic acid (16:1) | 0.1217 ± 0.0074 |
| Heptadecenoic acid (17:1) | 0.0320 ± 0.0023 | ||
| Oleic acid (18:1) | 19.9333 ± 0.4190 | ||
| 11-Eicosenoic acid (20:1) | 0.1753 ± 0.0146 | ||
| Erucic acid (22:1) | 0.0270 ± 0.0259 | ||
| Nervonic acid (24:1) | 0.0150 ± 0.0111 | ||
| Polyunsaturated (PUFA) | Linoleic acid (18:2) | 57.6667 ± 0.7717 | |
| Linolenic acid (18:3) | 13.3333 ± 0.2867 | ||
| Eicosadienoic acid (20:2) | 0.0261 ± 0.0015 | ||
| SFA | 8.6713 ± 0.1688 | ||
| UFA | 91.3304 ± 0.1675 | ||
| MUFA | 20.3043 ± 0.4147 | ||
| PUFA | 71.0261 ± 0.5715 | ||
| MUFA/SFA | 2.3417 ± 0.0296 | ||
| PUFA/SFA | 8.1952 ± 0.2219 | ||
| MUFA/PUFA | 0.2859 ± 0.0081 | ||
| KEGGID | Description | Stage | GeneRatio | Count | Up | Down |
|---|---|---|---|---|---|---|
| ath00061 | Fatty acid biosynthesis | G2 vs. G1 | 10/853 | 10 | 3 | 7 |
| G3 vs. G1 | 27/1735 | 27 | 11 | 16 | ||
| G4 vs. G1 | 37/2191 | 37 | 8 | 29 | ||
| G5 vs. G1 | 50/3080 | 50 | 6 | 44 | ||
| ath00062 | Fatty acid elongation | G2 vs. G1 | 7/853 | 7 | 2 | 5 |
| G3 vs. G1 | 15/1735 | 15 | 4 | 11 | ||
| G4 vs. G1 | 17/2191 | 17 | 5 | 12 | ||
| G5 vs. G1 | 23/3080 | 23 | 9 | 14 | ||
| ath00100 | Steroid biosynthesis | G2 vs. G1 | 14/853 | 14 | 4 | 10 |
| G3 vs. G1 | 21/1735 | 21 | 6 | 15 | ||
| G4 vs. G1 | 26/2191 | 26 | 6 | 20 | ||
| G5 vs. G1 | 35/3080 | 35 | 8 | 27 | ||
| ath00561 | Glycerolipid metabolism | G2 vs. G1 | 21/853 | 21 | 10 | 11 |
| G3 vs. G1 | 29/1735 | 29 | 14 | 15 | ||
| G4 vs. G1 | 35/2191 | 35 | 14 | 21 | ||
| G5 vs. G1 | 44/3080 | 44 | 20 | 24 | ||
| ath00564 | Glycerophospholipid metabolism | G2 vs. G1 | 19/853 | 19 | 9 | 10 |
| G3 vs. G1 | 37/1735 | 37 | 15 | 22 | ||
| G4 vs. G1 | 46/2191 | 46 | 18 | 28 | ||
| G5 vs. G1 | 60/3080 | 60 | 25 | 35 | ||
| ath00590 | Arachidonic acid metabolism | G2 vs. G1 | 6/853 | 6 | 4 | 2 |
| G3 vs. G1 | 9/1735 | 9 | 5 | 4 | ||
| G4 vs. G1 | 10/2191 | 10 | 7 | 3 | ||
| G5 vs. G1 | 16/3080 | 16 | 11 | 5 | ||
| ath00592 | alpha-Linolenic acid metabolism | G2 vs. G1 | 11/853 | 11 | 7 | 4 |
| G3 vs. G1 | 21/1735 | 21 | 11 | 10 | ||
| G4 vs. G1 | 21/2191 | 21 | 13 | 8 | ||
| G5 vs. G1 | 33/3080 | 33 | 19 | 14 | ||
| ath01040 | Biosynthesis of unsaturated fatty acids | G2 vs. G1 | 7/853 | 7 | 3 | 4 |
| G3 vs. G1 | 15/1735 | 15 | 7 | 8 | ||
| G4 vs. G1 | 13/2191 | 13 | 6 | 7 | ||
| G5 vs. G1 | 18/3080 | 18 | 5 | 13 | ||
| ko00071 | Fatty acid degradation | G2 vs. G1 | 16/853 | 16 | 6 | 10 |
| G3 vs. G1 | 26/1735 | 26 | 10 | 16 | ||
| G4 vs. G1 | 29/2191 | 29 | 11 | 18 | ||
| G5 vs. G1 | 43/3080 | 43 | 22 | 21 | ||
| ko00591 | Linoleic acid metabolism | G2 vs. G1 | 2/853 | 2 | 2 | 0 |
| G3 vs. G1 | 2/1735 | 2 | 2 | 0 | ||
| G4 vs. G1 | 4/2191 | 4 | 3 | 1 | ||
| G5 vs. G1 | 7/3080 | 7 | 5 | 2 | ||
| ko00600 | Sphingolipid metabolism | G2 vs. G1 | 2/853 | 2 | 0 | 2 |
| G3 vs. G1 | 7/1735 | 7 | 1 | 6 | ||
| G4 vs. G1 | 10/2191 | 10 | 4 | 6 | ||
| G5 vs. G1 | 14/3080 | 14 | 7 | 7 | ||
| ko00073 | Cutin, suberine and wax biosynthesis | G2 vs. G1 | 2/853 | 2 | 1 | 1 |
| G3 vs. G1 | 5/1735 | 5 | 3 | 2 | ||
| G4 vs. G1 | 5/2191 | 5 | 2 | 3 | ||
| G5 vs. G1 | 7/3080 | 7 | 4 | 3 | ||
| ko00565 | Ether lipid metabolism | G2 vs. G1 | 6/853 | 6 | 2 | 4 |
| G3 vs. G1 | 9/1735 | 9 | 1 | 8 | ||
| G4 vs. G1 | 14/2191 | 14 | 5 | 9 | ||
| G5 vs. G1 | 21/3080 | 21 | 8 | 13 | ||
| ko00072 | Synthesis and degradation of ketone bodies | G2 vs. G1 | 2/853 | 2 | 2 | 0 |
| G3 vs. G1 | 4/1735 | 4 | 2 | 2 | ||
| G4 vs. G1 | 4/2191 | 4 | 2 | 2 | ||
| G5 vs. G1 | 6/3080 | 6 | 3 | 3 | ||
| ko01212 | Fatty acid metabolism | G2 vs. G1 | 18/853 | 18 | 6 | 12 |
| G3 vs. G1 | 43/1735 | 43 | 17 | 26 | ||
| G4 vs. G1 | 50/2191 | 50 | 13 | 37 | ||
| G5 vs. G1 | 69/3080 | 69 | 17 | 52 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, W.; Zhang, D.; Ma, Z. Transcriptome Analysis of Genes Involved in Fatty Acid and Lipid Biosynthesis in Developing Walnut (Juglans regia L.) Seed Kernels from Qinghai Plateau. Plants 2022, 11, 3207. https://doi.org/10.3390/plants11233207
Shi W, Zhang D, Ma Z. Transcriptome Analysis of Genes Involved in Fatty Acid and Lipid Biosynthesis in Developing Walnut (Juglans regia L.) Seed Kernels from Qinghai Plateau. Plants. 2022; 11(23):3207. https://doi.org/10.3390/plants11233207
Chicago/Turabian StyleShi, Wenjun, Defang Zhang, and Zhong Ma. 2022. "Transcriptome Analysis of Genes Involved in Fatty Acid and Lipid Biosynthesis in Developing Walnut (Juglans regia L.) Seed Kernels from Qinghai Plateau" Plants 11, no. 23: 3207. https://doi.org/10.3390/plants11233207