
Contributions of Beneficial Microorganisms in Soil Remediation and Quality Improvement of Medicinal Plants


Abstract
:1. Introduction
2. Review Methodology
3. Soil Problems in Medicinal Plant Cultivation
3.1. Continuous Cropping Obstacle
3.2. Soil Hardening and Salinization
3.3. Soil Acidification
3.4. Contamination of Pesticides, Heavy Metals and Organic Pollutants in Soil
4. Roles of Microorganisms in Soil Bioremediation and the Potential Mechanisms
4.1. Regulation of Soil Properties and Fertility
4.2. Degradation of Pesticides and Organic Pollutants in Soil
4.3. Removal of Heavy Metals from Soil
5. Function of Soil Microorganisms in Growth Promotion and Quality Improvement of Medicinal Plants and the Potential Mechanisms
5.1. Microorganisms Enhance the Environmental Stress Resistance and Growth of Medicinal Plants
5.2. Microorganisms Promote the Accumulation of Active Ingredients in Medicinal Plants
5.3. Soil Microorganisms can Enhance the Disease Resistance of Medicinal Plants
5.4. Microbes Alleviate the Toxicity and Accumulation of Soil Pollutants in Medicinal Plants
6. Development and Application of Microbial Inoculants for Medicinal Plants
7. Conclusions and Future Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Wang, W.Y.; Zhou, H.; Wang, Y.F.; Sang, B.S.; Liu, L. Current policies and measures on the development of traditional Chinese medicine in China. Pharmacol. Res. 2021, 163, 105187. [Google Scholar] [CrossRef] [PubMed]
- Astutik, S.; Pretzsch, J.; Kimengsi, J.N. Asian medicinal plants’ production and utilization potentials: A review. Sustainability 2019, 11, 5483. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.; Tang, X.; Jia, Z.; Li, C.; Ma, J.; Zhang, J. The effects of ecological factors on the main medicinal components of Dendrobium officinale under different cultivation modes. Forests 2020, 11, 94. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Jiang, D.; Yang, M.; Ma, T.; Ding, F.; Hao, M.; Chen, Y.; Zhang, C.; Zhang, X.; Li, M. Influence of the environment on the distribution and quality of Gentiana dahurica Fisch. Front. Plant Sci. 2021, 12, 706822. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Mou, D.; Luo, L.; Zhong, W.; Duan, L.; Zou, X. Different cultivation environments affect the yield, bacterial community and metabolites of Cordyceps cicadae. Front. Microbiol. 2021, 12, 669785. [Google Scholar] [CrossRef]
- Dong, Z.; Rao, M.P.N.; Liao, T.; Li, L.; Liu, Y.; Xiao, M.; Mohamad, O.A.A.; Tian, Y.; Li, W. Diversity and function of rhizosphere microorganisms between wild and cultivated medicinal plant Glycyrrhiza uralensis Fisch under different soil conditions. Arch. Microbiol. 2021, 203, 3657–3665. [Google Scholar] [CrossRef] [PubMed]
- Lang, T.; Pan, L.; Liu, B.; Guo, T.; Hou, X. Vegetation characteristics and response to the soil properties of three medicinal plant communities in Altay Prefecture, China. Sustainability 2020, 12, 10306. [Google Scholar] [CrossRef]
- Xin, A.; Li, X.; Jin, H.; Yang, X.; Zhao, R.; Liu, J.; Qin, B. The accumulation of reactive oxygen species in root tips caused by autotoxic allelochemicals-A significant factor for replant problem of Angelica sinensis (Oliv.) Diels. Ind. Crop. Prod. 2019, 138, 111432. [Google Scholar] [CrossRef]
- Vasickova, J.; Hvezdova, M.; Kosubova, P.; Hofman, J. Ecological risk assessment of pesticide residues in arable soils of the Czech Republic. Chemosphere 2019, 216, 479–487. [Google Scholar] [CrossRef]
- Zhang, Y.; Zheng, Y.; Xia, P.; Xun, L.; Liang, Z. Impact of continuous Panax notoginseng plantation on soil microbial and biochemical properties. Sci. Rep. 2019, 9, 13205. [Google Scholar] [CrossRef]
- Pang, Z.; Mao, X.; Xia, Y.; Xiao, J.; Wang, X.; Xu, P.; Liu, G. Multiomics reveals the effect of root rot on Polygonati rhizome and identifies pathogens and biocontrol strain. Microbiol. Spectr. 2022, 10, e02385-21. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Pandey, A.K.; Kim, S.; Singh, S.P.; Chaturvedi, P.; Varjani, S. Critical review on microbial community during in-situ bioremediation of heavy metals from industrial wastewater. Environ. Technol. Innov. 2021, 24, 101826. [Google Scholar] [CrossRef]
- Geng, Y.; Jiang, L.; Jiang, H.; Wang, L.; Peng, Y.; Wang, C.; Shi, X.; Gu, J.; Wang, Y.; Zhu, J.; et al. Assessment of heavy metals, fungicide quintozene and its hazardous impurity residues in medical Panax notoginseng (Burk) F.H. Chen root. Biomed. Chromatogr. 2019, 33, e4378. [Google Scholar] [CrossRef] [PubMed]
- The Ministry of Environmental Protection; The Ministry of Land and Resources. Report on the National General Survey of Soil Contamination. 2014. Available online: https://www.mee.gov.cn/gkml/sthjbgw/qt/201404/W020140417558995804588.pdf (accessed on 22 September 2022).
- Ara, T.; Nisa, W.U.; Aziz, R.; Rafiq, M.T.; Gill, R.A.; Hayat, M.T.; Afridi, U. Health risk assessment of hexachlorocyclohexane in soil, water and plants in the agricultural area of Potohar region, Punjab, Pakistan. Environ. Geochem. Health 2021, 43, 1–17. [Google Scholar] [CrossRef]
- Fu, Y.; Dou, X.; Lu, Q.; Qin, J.; Luo, J.; Yang, M. Comprehensive assessment for the residual characteristics and degradation kinetics of pesticides in Panax notoginseng and planting soil. Sci. Total Environ. 2020, 714, 136718. [Google Scholar] [CrossRef]
- Swamy, M.K.; Nalina, N.; Nalina, D.; Akhtar, M.S.; Purushotham, B. Heavy metal stress and tolerance in plants mediated by rhizospheric microbes. In Salt Stress, Microbes, and Plant Interactions: Causes and Solution; Akhtar, M., Ed.; Springer: Singapore, 2019; pp. 181–198. [Google Scholar] [CrossRef]
- Upton, R.N.; Bach, E.M.; Hofmockel, K.S. Spatio-temporal microbial community dynamics within soil aggregates. Soil Biol. Biochem. 2019, 132, 58–68. [Google Scholar] [CrossRef]
- Kui, L.; Chen, B.; Chen, J.; Sharifi, R.; Dong, Y.; Zhang, Z.; Miao, J. A comparative analysis on the structure and function of the Panax notoginseng rhizosphere microbiome. Front. Microbiol. 2021, 12, 673512. [Google Scholar] [CrossRef] [PubMed]
- Genre, A.; Lanfranco, L.; Perotto, S.; Bonfante, P. Unique and common traits in mycorrhizal symbioses. Nat. Rev. Microbiol. 2020, 18, 649–660. [Google Scholar] [CrossRef]
- Vejan, P.; Abdullah, R.; Khadiran, T.; Ismail, S.; Nasrulhaq, B.A. Role of plant growth promoting rhizobacteria in agricultural sustainability—A review. Molecules 2016, 21, 573. [Google Scholar] [CrossRef] [Green Version]
- Khan, I.; Aftab, M.; Shakir, S.; Ali, M.; Qayyum, S.; Rehman, M.U.; Haleem, K.S.; Touseef, I. Mycoremediation of heavy metal (Cd and Cr)-polluted soil through indigenous metallotolerant fungal isolates. Environ. Monit. Assess. 2019, 191, 585. [Google Scholar] [CrossRef]
- Kour, D.; Rana, K.L.; Sheikh, I.; Kumar, V.; Yadav, A.N.; Dhaliwal, H.S.; Saxena, A.K. Alleviation of drought stress and plant growth promotion by Pseudomonas libanensis EU-LWNA-33, a drought-adaptive phosphorus-solubilizing bacterium. Proc. Nat. Acad. Sci. India Sect. B Biol. Sci. 2019, 90, 785–795. [Google Scholar] [CrossRef]
- Chaudhary, D.K.; Kim, J. Novosphingobium naphthae sp. nov., from oil-contaminated soil. Int. J. Syst. Evol. Microbiol. 2016, 66, 3170–3176. [Google Scholar] [CrossRef] [PubMed]
- Kalaimurugan, D.; Balamuralikrishnan, B.; Durairaj, K.; Vasudhevan, P.; Shivakumar, M.S.; Kaul, T.; Chang, S.W.; Ravindran, B.; Venkatesan, S. Isolation and characterization of heavy-metal-resistant bacteria and their applications in environmental bioremediation. Int. J. Environ. Sci. Technol. 2019, 17, 1455–1462. [Google Scholar] [CrossRef]
- Zhang, X.; Gao, Y.; Zang, P.; Zhao, Y.; He, Z.; Zhu, H.; Song, S.; Zhang, L. Study on the simultaneous degradation of five pesticides by Paenibacillus polymyxa from Panax ginseng and the characteristics of their products. Ecotoxicol. Environ. Saf. 2019, 168, 415–422. [Google Scholar] [CrossRef]
- Alawiye, T.; Babalola, O. Metagenomic insight into the community structure and functional genes in the sunflower rhizosphere microbiome. Agriculture 2021, 11, 167. [Google Scholar] [CrossRef]
- Brunel, C.; Pouteau, R.; Dawson, W.; Pester, M.; Ramirez, K.S.; Van Kleunen, M. Towards unraveling macroecological patterns in rhizosphere microbiomes. Trends Plant Sci. 2020, 25, 1017–1029. [Google Scholar] [CrossRef]
- Saleem, M.; Hu, J.; Jousset, A. More than the sum of its parts: Microbiome biodiversity as a driver of plant growth and soil health. Annu. Rev. Ecol. Evol. Syst. 2019, 50, 145–168. [Google Scholar] [CrossRef]
- Cao, P.; Wang, G.; Wei, X.; Chen, S.; Han, J. How to improve CHMs quality: Enlighten from CHMs ecological cultivation. Chin. Herb. Med. 2021, 13, 301–312. [Google Scholar] [CrossRef]
- Haider, F.U.; Cai, L.; Coulter, J.A.; Cheema, S.A.; Wu, J.; Zhang, R.; Ma, W.; Farooq, M. Cadmium toxicity in plants: Impacts and remediation strategies. Ecotoxicol. Environ. Saf. 2021, 211, 111887. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Dai, S.; Wang, B.; Jiang, Y.; Ma, Y.; Pan, L.; Wu, K.; Huang, X.; Zhang, J.; Cai, Z.; et al. Autotoxic ginsenoside disrupts soil fungal microbiomes by stimulating potentially pathogenic microbes. Appl. Environ. Microbiol. 2020, 86, e00130-20. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Zhang, Q.; Chen, Y.; Su, Y.; Sun, S.; Chen, G. Different crop rotation systems change the rhizosphere bacterial community structure of Astragalus membranaceus (Fisch) Bge. var. mongholicus (Bge.) Hsiao. Appl. Soil Ecol. 2021, 166, 104003. [Google Scholar] [CrossRef]
- Pang, Z.; Tayyab, M.; Kong, C.; Liu, Q.; Liu, Y.; Hu, C.; Huang, J.; Weng, P.; Islam, W.; Lin, W.; et al. Continuous sugarcane planting negatively impacts soil microbial community structure, soil fertility, and sugarcane agronomic parameters. Microorganisms 2021, 9, 2008. [Google Scholar] [CrossRef]
- Gao, Z.; Hu, Y.; Han, M.; Xu, J.; Wang, X.; Liu, L.; Tang, Z.; Jiao, W.; Jin, R.; Liu, M.; et al. Effects of continuous cropping of sweet potatoes on the bacterial community structure in rhizospheric soil. BMC Microbiol. 2021, 21, 102. [Google Scholar] [CrossRef]
- Pang, Z.; Dong, F.; Liu, Q.; Lin, W.; Hu, C.; Yuan, Z. Soil metagenomics reveals effects of continuous sugarcane cropping on the structure and functional pathway of rhizospheric microbial community. Front. Microbiol. 2021, 12, 627569. [Google Scholar] [CrossRef] [PubMed]
- Xia, F.; Wang, L.; Chen, J.; Fu, M.; Wang, G.; Yan, Y.; Cui, L. Variations of microbial community in Aconitum carmichaeli Debx. rhizosphere soilin a short-term continuous cropping system. J. Microbiol. 2021, 59, 481–490. [Google Scholar]
- Liu, H.; Niu, M.; Zhu, S.; Zhang, F.; Liu, Q.; Liu, Y.; Liu, R.; Zhang, Y. Effect study of continuous monoculture on the quality of Salvia miltiorrhiza Bge Roots. Biomed. Res. Int. 2020, 2020, 4284385. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Guo, R.; Li, S.; Chen, Y.; Li, Z.; He, P.; Huang, X.; Huang, K. Effects of continuous cropping on soil, senescence, and yield of Tartary buckwheat. Agron. J. 2021, 113, 5102–5113. [Google Scholar] [CrossRef]
- Guo, K.; He, X.; Yan, Z.; Li, X.; Ren, X.; Pan, L.; Qin, B. Allelochemicals from the rhizosphere soil of cultivated Astragalus hoantchy. J. Agric. Food Chem. 2016, 64, 3345–3352. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Lu, L.-Y.; Hu, L.-Y.; Cao, W.; Sun, K.; Sun, Q.-B.; Siddikee, A.; Shi, R.-H.; Dai, C.-C. Evidence for the involvement of auxin, ethylene and ROS signaling during primary root inhibition of Arabidopsis by the allelochemical benzoic acid. Plant Cell Physiol. 2018, 59, 1889–1904. [Google Scholar] [CrossRef] [Green Version]
- Tan, G.; Liu, Y.; Peng, S.; Yin, H.; Meng, D.; Tao, J.; Gu, Y.; Li, J.; Yang, S.; Xiao, N.; et al. Soil potentials to resist continuous cropping obstacle: Three field cases. Environ. Res. 2021, 200, 111319. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Yan, Z.; He, X.; Li, X.; Qin, B. Allelochemicals from rhizosphere soils of Glycyrrhiza uralensis Fisch: Discovery of the autotoxic compounds of a traditional herbal medicine. Ind. Crop. Prod. 2017, 97, 302–307. [Google Scholar] [CrossRef]
- Li, X.; Lewis, E.E.; Liu, Q.; Li, H.; Bai, C.; Wang, Y. Effects of long-term continuous cropping on soil nematode community and soil condition associated with replant problem in strawberry habitat. Sci. Rep. 2016, 6, 30466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Wang, Z.; Niu, J.; Dang, K.; Zhang, S.; Wang, S.; Wang, Z. Changes in physicochemical properties, enzymatic activities, and the microbial community of soil significantly influence the continuous cropping of Panax quinquefolius L. (American ginseng). Plant Soil 2021, 463, 427–446. [Google Scholar] [CrossRef]
- Zhang, B.; Li, X.; Wang, F.; Li, M.; Zhang, J.; Gu, L.; Zhang, L.; Tu, W.; Zhang, Z. Assaying the potential autotoxins and microbial community associated with Rehmannia glutinosa replant problems based on its ‘autotoxic circle’. Plant Soil 2016, 407, 307–322. [Google Scholar] [CrossRef] [Green Version]
- Lei, H.; Liu, A.; Hou, Q.; Zhao, Q.; Guo, J.; Wang, Z. Diversity patterns of soil microbial communities in the Sophora flavescens rhizosphere in response to continuous monocropping. BMC Microbiol. 2020, 20, 272. [Google Scholar] [CrossRef]
- Wei, X.; Wang, X.; Cao, P.; Gao, Z.; Chen, A.J.; Han, J. Microbial community changes in the rhizosphere soil of healthy and rusty Panax ginseng and discovery of pivotal fungal genera associated with rusty roots. Biomed Res. Int. 2020, 2020, 8018525. [Google Scholar] [CrossRef] [Green Version]
- Zhu, B.; Wu, J.; Ji, Q.; Wu, W.; Dong, S.; Yu, J.; Zhang, Q.; Qin, L. Diversity of rhizosphere and endophytic fungi in Atractylodes macrocephala during continuous cropping. PeerJ 2020, 8, e8905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Z.; Han, M.; Hu, Y.; Li, Z.; Liu, C.; Wang, X.; Tian, Q.; Jiao, W.; Hu, J.; Liu, L.; et al. Effects of continuous cropping of sweet potato on the fungal community structure in rhizospheric soil. Front. Microbiol. 2019, 10, 2269. [Google Scholar] [CrossRef] [PubMed]
- Kulmatov, R.; Mirzaev, J.; Abuduwaili, J.; Karimov, B. Challenges for the sustainable use of water and land resources under a changing climate and increasing salinization in the Jizzakh irrigation zone of Uzbekistan. J. Arid Land 2020, 12, 90–103. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Sun, D.; Tang, Y.; Zhu, R.; Li, X.; Gruda, N.; Dong, J.; Duan, Z. Plastic shed soil salinity in China: Current status and next steps. J. Clean. Prod. 2021, 296, 126453. [Google Scholar] [CrossRef]
- Wu, R.; Sun, H.; Xue, J.; Yan, D.; Liu, Y.; Gui, D.; Wang, X.; Yang, J. Acceleration of soil salinity accumulation and soil degradation due to greenhouse cultivation: A survey of farmers’ practices in China. Environ. Monit. Assess. 2020, 192, 399. [Google Scholar] [CrossRef]
- Figueiredo, P.G.; Bicudo, S.J.; Chen, S.; Fernandes, A.M.; Tanamati, F.Y.; Djabou-Fondjo, A.S.M. Effects of tillage options on soil physical properties and cassava-dry-matter partitioning. Field Crop. Res. 2017, 204, 191–198. [Google Scholar] [CrossRef] [Green Version]
- Rengasamy, P. World salinization with emphasis on Australia. J. Exp. Bot. 2006, 57, 1017–1023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Etesami, H.; Beattie, G.A. Mining halophytes for plant growth-promoting halotolerant bacteria to enhance the salinity tolerance of non-halophytic crops. Front. Microbiol. 2018, 9, 148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bian, L.; Wang, J.; Liu, J.; Han, B. Spatiotemporal changes of soil salinization in the Yellow River Delta of China from 2015 to 2019. Sustainability 2021, 13, 822. [Google Scholar] [CrossRef]
- Huang, J.; Hartemink, A.E. Soil and environmental issues in sandy soils. Earth-Sci. Rev. 2020, 208, 103295. [Google Scholar] [CrossRef]
- Buvaneshwari, S.; Riotte, J.; Sekhar, M.; Sharma, A.K.; Helliwell, R.; Kumar, M.; Braun, J.J.; Ruiz, L. Potash fertilizer promotes incipient salinization in groundwater irrigated semi-arid agriculture. Sci. Rep. 2020, 10, 3691. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.P.; Rekha, P.D.; Arun, A.B.; Shen, F.T.; Lai, W.A.; Young, C.C. Phosphate solubilizing bacteria from subtropical soil and their tricalcium phosphate solubilizing abilities. Appl. Soil Ecol. 2006, 34, 33–41. [Google Scholar] [CrossRef]
- Elhanafi, L.; Houhou, M.; Rais, C.; Mansouri, I.; Elghadraoui, L.; Greche, H. Impact of excessive nitrogen fertilization on the biochemical quality, phenolic compounds, and antioxidant power of Sesamum indicum L Seeds. J. Food Qual. 2019, 2019, 9428092. [Google Scholar] [CrossRef] [Green Version]
- Ameixa, O.M.C.C.; Marques, B.; Fernandes, V.S.; Soares, A.M.V.M.; Calado, R.; Lillebø, A.I. Dimorphic seeds of Salicornia ramosissima display contrasting germination responses under different salinities. Ecol. Eng. 2016, 87, 120–123. [Google Scholar] [CrossRef]
- Mahajan, M.; Sharma, S.; Kumar, P.; Pal, P.K. Foliar application of KNO3 modulates the biomass yield, nutrient uptake and accumulation of secondary metabolites of Stevia rebaudiana under saline conditions. Ind. Crop. Prod. 2020, 145, 112102. [Google Scholar] [CrossRef]
- Mohanavelu, A.; Naganna, S.R.; Al-Ansari, N. Irrigation induced salinity and sodicity hazards on soil and groundwater: An overview of its causes, impacts and mitigation strategies. Agriculture 2021, 11, 983. [Google Scholar] [CrossRef]
- Ashraf, M.; Foolad, M.R. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Mansour, M.M.F. Protection of plasma membrane of onion epidermal cells by glycinebetaine and proline against NaCl stress. Plant Physiol. Biochem. 1998, 36, 767–772. [Google Scholar] [CrossRef]
- Ketchum, R.E.B.; Warren, R.S.; Klima, L.J.; Lopez-Gutiérrez, F.; Nabors, M.W. The mechanism and regulation of proline accumulation in suspension cell cultures of the halophytic grass Distichlis spicata L. J. Plant Physiol. 1991, 137, 368–374. [Google Scholar] [CrossRef]
- Kubotera, H. Analysis of problems in certain soils of the Kyushu Okinawa region for suitable management. Soil Sci. Plant Nutr. 2020, 66, 15–20. [Google Scholar] [CrossRef]
- Wahlström, E.M.; Kristensen, H.L.; Thomsen, I.K.; Labouriau, R.; Pulido-Moncada, M.; Nielsen, J.A.; Munkholm, L.J. Subsoil compaction effect on spatio-temporal root growth, reuse of biopores and crop yield of spring barley. Eur. J. Agron. 2021, 123, 126225. [Google Scholar] [CrossRef]
- Gaab, T. Effect of soil compaction and N fertilization on soil pore characteristics and physical quality of sandy loam soil under red clover/grass sward. Soil Tillage Res. 2014, 144, 8–19. [Google Scholar]
- Pandey, B.K.; Huang, G.; Bhosale, R.; Hartman, S.; Sturrock, C.J.; Jose, L.; Martin, O.C.; Karady, M.; Voesenek, L.A.C.J.; Ljung, K.; et al. Plant roots sense soil compaction through restricted ethylene diffusion. Science 2021, 371, 276–280. [Google Scholar] [CrossRef]
- Zhang, H.; Voroney, R.P.; Price, G.W.; White, A.J. Sulfur-enriched biochar as a potential soil amendment and fertiliser. Soil Res. 2017, 55, 93–99. [Google Scholar] [CrossRef]
- Iturri, L.A.; Buschiazzo, D.E. Light acidification in N-fertilized loess soils along a climosequence affected chemical and mineralogical properties in the short term. Catena 2016, 139, 92–98. [Google Scholar] [CrossRef]
- Wu, Z.; Sun, X.; Sun, Y.; Yan, J.; Zhao, Y.; Chen, J. Soil acidification and factors controlling topsoil pH shift of cropland in central China from 2008 to 2018. Geoderma 2022, 408, 115586. [Google Scholar] [CrossRef]
- Li, Y.; Sun, J.; Tian, D.; Wang, J.; Ha, D.; Qu, Y.; Jing, G.; Niu, S. Soil acid cations induced reduction in soil respiration under nitrogen enrichment and soil acidification. Sci. Total Environ. 2018, 615, 1535–1546. [Google Scholar] [CrossRef] [PubMed]
- Cecchini, G.; Andreetta, A.; Marchetto, A.; Carnicelli, S. Atmospheric deposition control of soil acidification in central Italy. Catena 2019, 182, 104102. [Google Scholar] [CrossRef]
- Huang, J.; Zhou, K.; Zhang, W.; Liu, J.; Ding, X.; Cai, X.; Mo, J. Sulfur deposition still contributes to forest soil acidification in the Pearl River Delta, South China, despite the control of sulfur dioxide emission since 2001. Environ. Sci. Pollut. Res. Int. 2019, 26, 12928–12939. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Qiu, Y.; Cheng, L.; Wang, Y.; Liu, L.; Tu, C.; Bowman, D.C.; Burkey, K.O.; Bian, X.; Zhang, W.; et al. Atmospheric CO2 enrichment and reactive nitrogen inputs interactively stimulate soil cation losses and acidification. Environ. Sci. Technol. 2018, 52, 6895–6902. [Google Scholar] [CrossRef] [PubMed]
- Raza, S.; Miao, N.; Wang, P.; Ju, X.; Chen, Z.; Zhou, J.; Kuzyakov, Y.; Na, M. Dramatic loss of inorganic carbon by nitrogen-induced soil acidification in Chinese croplands. Glob. Chang. Biol. 2020, 26, 3738–3751. [Google Scholar] [CrossRef]
- Cai, Z.; Wang, B.; Xu, M.; Zhang, H.; He, X.; Zhang, L.; Gao, S. Intensified soil acidification from chemical N fertilization and prevention by manure in an 18-year field experiment in the red soil of southern China. J. Soils Sediments 2014, 15, 260–270. [Google Scholar] [CrossRef]
- Hao, T.; Zhu, Q.; Zeng, M.; Shen, J.; Shi, X.; Liu, X.; Zhang, F.; de Vries, W. Impacts of nitrogen fertilizer type and application rate on soil acidification rate under a wheat-maize double cropping system. J. Environ. Manag. 2020, 270, 110888. [Google Scholar] [CrossRef]
- Hao, T.; Zhu, Q.; Zeng, M.; Shen, J.; Shi, X.; Liu, X.; Zhang, F.; de Vries, W. Quantification of the contribution of nitrogen fertilization and crop harvesting to soil acidification in a wheat-maize double cropping system. Plant Soil 2019, 434, 167–184. [Google Scholar] [CrossRef]
- Zhao, Q.; Xiong, W.; Xing, Y.; Sun, Y.; Lin, X.; Dong, Y. Long-term coffee monoculture alters soil chemical properties and microbial communities. Sci. Rep. 2018, 8, 6116. [Google Scholar] [CrossRef] [PubMed]
- Coventry, D.R.; Slattery, W.J. Acidification of soil associated with lupins grown in a crop rotation in north-eastern Victoria. Aust. J. Agric. Res. 1991, 42, 391–397. [Google Scholar] [CrossRef]
- Zhang, Y.; He, X.; Zhao, J.; Zhang, Y.; Shi, X. Soil acidification under long-term tobacco plantation results in alterations of mineralogical properties in an Alisol. Arch. Agron. Soil Sci. 2016, 62, 1033–1040. [Google Scholar] [CrossRef]
- Yu, Y.; Yang, J.; Zeng, S.; Wu, D.; Jacobs, D.F.; Sloan, J.L. Soil pH, organic matter, and nutrient content change with the continuous cropping of Cunninghamia lanceolata plantations in South China. J. Soils Sediments 2016, 17, 2230–2238. [Google Scholar] [CrossRef]
- Zhang, Y.; He, X.; Liang, H.; Zhao, J.; Zhang, Y.; Xu, C.; Shi, X. Long-term tobacco plantation induces soil acidification and soil base cation loss. Environ. Sci. Pollut. Res. Int. 2016, 23, 5442–5450. [Google Scholar] [CrossRef]
- Golez, N.V.; Kyuma, K. Influence of pyrite oxidation and soil acidification on some essential nutrient elements. Aquac. Eng. 1997, 16, 107–124. [Google Scholar] [CrossRef]
- Kang, Y.; Zhu, S.; Li, G.; Jiang, Y.; Teng, W.; Luo, M.; Wei, J.; Cao, F.; Wang, Z.; Huang, J. Phosphorus applied to the root half without Al3+ exposure can alleviate Al toxicity on the other root half of the same eucalyptus seedling. J. Plant Nutr. 2020, 44, 829–844. [Google Scholar] [CrossRef]
- Zarif, N.; Khan, A.; Wang, Q. Linking soil acidity to P fractions and exchangeable base cations under increased N and P fertilization of mono and mixed plantations in Northeast China. Forests 2020, 11, 1274. [Google Scholar] [CrossRef]
- Babourina, O.; Hawkins, B.; Lew, R.R.; Newman, I.; Shabala, S. K+ transport by Arabidopsis root hairs at low pH. Funct. Plant Biol. 2001, 28, 637–643. [Google Scholar] [CrossRef]
- Chen, L.; Guo, L.; Zhou, Q.; Liu, M.; Zhan, S.; Pan, X.; Zeng, Y. Response of soil fertility and Cu and Cd availability to biochar application on paddy soils with different acidification levels. Biomass Conv. Bioref. 2020, 12, 1493–1502. [Google Scholar] [CrossRef]
- Zhang, F.; Jin, Q.; Peng, H.; Zhu, T. Soil acidification in moso bamboo (Phyllostachys edulis) forests and changes of soil metal ions (Cu, Pb) concentration. Arch. Agron. Soil Sci. 2021, 67, 1799–1808. [Google Scholar] [CrossRef]
- Liu, C.; Liang, M.; Nie, Y.; Tang, J.; Siddique, K.H.M. The conversion of tropical forests to rubber plantations accelerates soil acidification and changes the distribution of soil metal ions in topsoil layers. Sci. Total Environ. 2019, 696, 134082. [Google Scholar] [CrossRef]
- Zhou, X.; Khashi, U.R.M.; Liu, J.; Wu, F. Soil acidification mediates changes in soil bacterial community assembly processes in response to agricultural intensification. Environ. Microbiol. 2021, 23, 4741–4755. [Google Scholar] [CrossRef] [PubMed]
- Xing, W.; Lu, X.; Ying, J.; Lan, Z.; Chen, D.; Bai, Y. Disentangling the effects of nitrogen availability and soil acidification on microbial taxa and soil carbon dynamics in natural grasslands. Soil Biol. Biochem. 2022, 164, 108495. [Google Scholar] [CrossRef]
- Chen, Z.; Maltz, M.R.; Zhang, Y.; O’Brien, B.J.; Neff, M.; Wang, Y.; Cao, J. Plantations of Cinnamomum camphora (Linn) Presl with distinct soil bacterial communities mitigate soil acidity within polluted locations in Southwest China. Forests 2021, 12, 657. [Google Scholar] [CrossRef]
- Muneer, M.A.; Hou, W.; Li, J.; Huang, X.; ur Rehman Kayani, M.; Cai, Y.; Yang, W.; Wu, L.; Ji, B.; Zheng, C. Soil pH: A key edaphic factor regulating distribution and functions of bacterial community along vertical soil profiles in red soil of pomelo orchard. BMC Microbiol. 2022, 22, 38. [Google Scholar] [CrossRef]
- Li, J.; Zheng, T.; Liu, C. Soil acidification enhancing the growth and metabolism inhibition of PFOS and Cr(VI) to bacteria involving oxidative stress and cell permeability. Environ. Pollut. 2021, 275, 116650. [Google Scholar] [CrossRef] [PubMed]
- Grube, A.; Donaldson, D.; Kiely, T.; Wu, L. Pesticides Industry Sales and Usage: 2006 and 2007 Market Estimates. 2011. Available online: https://www.epa.gov/sites/default/files/2015-10/documents/market_estimates2007.pdf (accessed on 22 September 2022).
- Kumar, S.; Kaushik, G.; Dar, M.A.; Nimesh, S.; Lopez-Chuken, U.J.; Villarreal-Chiu, J.F. Microbial degradation of organophosphate pesticides; A review. Pedosphere 2018, 28, 190–208. [Google Scholar] [CrossRef]
- Kaur, I.; Gaur, V.K.; Regar, R.K.; Roy, A.; Srivastava, P.K.; Gaur, R.; Manickam, N.; Barik, S.K. Plants exert beneficial influence on soil microbiome in a HCH contaminated soil revealing advantage of microbe-assisted plant-based HCH remediation of a dumpsite. Chemosphere 2021, 280, 130690. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, K.S.; Gul, P.; Gul, M.M. Efficient fungal and bacterial facilitated remediation of thiencarbazone methyl in the environment. Environ. Res. 2020, 188, 109811. [Google Scholar] [CrossRef]
- Wang, L.; Wang, J.; Zhu, L.; Wang, J. Toxic effects of oxytetracycline and copper, separately or combined, on soil microbial biomasses. Environ. Geochem. Health 2018, 40, 763–776. [Google Scholar] [CrossRef] [PubMed]
- Jampani, M.; Huelsmann, S.; Liedl, R.; Sonkamble, S.; Ahmed, S.; Amerasinghe, P. Spatio-temporal distribution and chemical characterization of groundwater quality of a wastewater irrigated system: A case study. Sci. Total Environ. 2018, 636, 1089–1098. [Google Scholar] [CrossRef] [PubMed]
- Sikka, R.; Nayyar, V.; Sidhu, S.S. Monitoring of Cd pollution in soils and plants irrigated with untreated sewage water in some industrialized cities of Punjab, India. Environ. Monit. Assess. 2009, 154, 53–64. [Google Scholar] [CrossRef]
- Shaheen, S.M.; Shams, M.S.; Khalifa, M.R.; El-Dali, M.A.; Rinklebe, J. Various soil amendments and environmental wastes affect the (im) mobilization and phytoavailability of potentially toxic elements in a sewage effluent irrigated sandy soil. Ecotoxicol. Environ. Saf. 2017, 142, 375–387. [Google Scholar] [CrossRef] [PubMed]
- Qin, G.; Niu, Z.; Yu, J.; Li, Z.; Ma, J.; Xiang, P. Soil heavy metal pollution and food safety in China: Effects, sources and removing technology. Chemosphere 2021, 267, 129205. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Luan, Y.; Ning, Y.; Wang, L. Effects and mechanisms of microbial remediation of heavy metals in soil: A critical review. Appl. Sci. 2018, 8, 1336. [Google Scholar] [CrossRef] [Green Version]
- Taha, S.M.; Gadalla, S.A. Development of an efficient method for multi residue analysis of 160 pesticides in herbal plant by ethyl acetate hexane mixture with direct injection to GC-MS/MS. Talanta 2017, 174, 767–779. [Google Scholar] [CrossRef]
- Durgnat, J.M.; Heuser, J.; Andrey, D.; Perrin, C. Quality and safety assessment of ginseng extracts by determination of the contents of pesticides and metals. Food Addit. Contam. 2005, 22, 1224–1230. [Google Scholar] [CrossRef]
- Kumar, N.; Kulsoom, M.; Shukla, V.; Kumar, D.; Priyanka; Kumar, S.; Tiwari, J.; Dwivedi, N. Profiling of heavy metal and pesticide residues in medicinal plants. Environ. Sci. Pollut. Res. Int. 2018, 25, 29505–29510. [Google Scholar] [CrossRef]
- Harris, E.S.; Cao, S.; Littlefield, B.A.; Craycroft, J.A.; Scholten, R.; Kaptchuk, T.; Fu, Y.; Wang, W.; Liu, Y.; Chen, H.; et al. Heavy metal and pesticide content in commonly prescribed individual raw Chinese herbal medicines. Sci. Total Environ. 2011, 409, 4297–4305. [Google Scholar] [CrossRef] [Green Version]
- Maitlo, A.A.; Jatoi, W.B.; Memon, A.F.; Soomro, A.H.; Bhayo, M.S. Assessment of Zinc, Lead, Chromium, and Cobalt in commonly consumed herbal medicines in Sindh, Pakistan. Biol. Trace Elem. Res. 2021, 199, 2366–2374. [Google Scholar] [CrossRef]
- Singh, S.; Singh, A.; Bashri, G.; Prasad, S.M. Impact of Cd stress on cellular functioning and its amelioration by phytohormones: An overview on regulatory network. Plant Growth Regul. 2016, 80, 253–263. [Google Scholar] [CrossRef]
- Scoccianti, V.; Crinelli, R.; Tirillini, B.; Mancinelli, V.; Speranza, A. Uptake and toxicity of Cr(III) in celery seedlings. Chemosphere 2006, 64, 1695–1703. [Google Scholar] [CrossRef] [PubMed]
- Ahsan, M.; Younis, A.; Jaskani, M.J.; Tufail, A.; Riaz, A.; Schwinghamer, T.; Tariq, U.; Nawaz, F. Heavy metal accumulation imparts structural differences in fragrant Rosa species irrigated with marginal quality water. Ecotoxicol. Environ. Saf. 2018, 159, 240–248. [Google Scholar] [CrossRef]
- Liu, L.; Li, S.; Guo, J.; Li, N.; Jiang, M.; Li, X. Low temperature tolerance is depressed in wild-type and abscisic acid-deficient mutant barley grown in Cd-contaminated soil. J. Hazard. Mater. 2022, 430, 128489. [Google Scholar] [CrossRef]
- Wang, Q.; Zhang, J.; Zhao, B.; Xin, X.; Zhang, C.; Zhang, H. The influence of long-term fertilization on cadmium (Cd) accumulation in soil and its uptake by crops. Environ. Sci. Pollut. Res. Int. 2014, 21, 10377–10385. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Kim, S.U.; Jung, K.Y.; Lee, S.; Choi, Y.D.; Owens, V.N.; Kumar, S.; Yun, S.W.; Hong, C.O. Cadmium phytoavailability from 1976 through 2016: Changes in soil amended with phosphate fertilizer and compost. Sci. Total Environ. 2021, 762, 143132. [Google Scholar] [CrossRef] [PubMed]
- Pizzol, M.; Smart, J.C.R.; Thomsen, M. External costs of cadmium emissions to soil: A drawback of phosphorus fertilizers. J. Clean. Prod. 2014, 84, 475–483. [Google Scholar] [CrossRef]
- Zhang, S.; Yao, H.; Lu, Y.; Yu, X.; Wang, J.; Sun, S.; Liu, M.; Li, D.; Li, Y.-F.; Zhang, D. Uptake and translocation of polycyclic aromatic hydrocarbons (PAHs) and heavy metals by maize from soil irrigated with wastewater. Sci. Rep. 2017, 7, 12165. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.; Liu, Y.; Yang, K.; Zhu, L.; White, J.C.; Lin, D. Synergistic remediation of PCB-contaminated soil with nanoparticulate zero-valent iron and alfalfa: Targeted changes in the root metabolite-dependent microbial community. Environ. Sci. Nano 2021, 8, 986–999. [Google Scholar] [CrossRef]
- Sakshi; Singh, S.K.; Haritash, A.K. Polycyclic aromatic hydrocarbons: Soil pollution and remediation. Int. J. Environ. Sci. Technol. 2019, 16, 6489–6512. [Google Scholar] [CrossRef]
- Elgallal, M.; Fletcher, L.; Evans, B. Assessment of potential risks associated with chemicals in wastewater used for irrigation in arid and semiarid zones: A review. Agric. Water Manag. 2016, 177, 419–431. [Google Scholar] [CrossRef]
- Zhang, X.; Zhong, T.; Liu, L.; Ouyang, X. Impact of soil heavy metal pollution on food safety in China. PLoS ONE 2015, 10, e0135182. [Google Scholar] [CrossRef] [Green Version]
- Karami-Mohajeri, S.; Abdollahi, M. Toxic influence of organophosphate, carbamate, and organochlorine pesticides on cellular metabolism of lipids, proteins, and carbohydrates: A systematic review. Hum. Exp. Toxicol. 2011, 30, 1119–1140. [Google Scholar] [CrossRef]
- Zhang, X.; Zhong, T.; Chen, D.; Cheng, M.; Liu, L.; Zhang, X.; Li, X. Assessment of arsenic (As) occurrence in arable soil and its related health risk in China. Environ. Geochem. Health 2016, 38, 691–702. [Google Scholar] [CrossRef] [PubMed]
- Shen, T.; Liu, L.; Li, Y.; Wang, Q.; Dai, J.; Wang, R. Long-term effects of untreated wastewater on soil bacterial communities. Sci. Total Environ. 2019, 646, 940–950. [Google Scholar] [CrossRef]
- Zhang, M.; Teng, Y.; Xu, Z.; Wang, J.; Christie, P.; Luo, Y. Cumulative effects of repeated chlorothalonil application on soil microbial activity and community in contrasting soils. J. Soils Sediments 2016, 16, 1754–1763. [Google Scholar] [CrossRef]
- Zhang, C.; Zhou, T.; Zhu, L.; Juhasz, A.; Du, Z.; Li, B.; Wang, J.; Wang, J.; Sun, Y. Response of soil microbes after direct contact with pyraclostrobin in fluvo-aquic soil. Environ. Pollut. 2019, 255, 113164. [Google Scholar] [CrossRef]
- Ren, G.; Ren, W.; Teng, Y.; Li, Z. Evident bacterial community changes but only slight degradation when polluted with pyrene in a red soil. Front. Microbiol. 2015, 6, 22. [Google Scholar] [CrossRef] [Green Version]
- Estrada, G.A.; Baldani, V.L.D.; De Oliveira, D.M.; Urquiaga, S.; Baldani, J.I. Selection of phosphate-solubilizing diazotrophic Herbaspirillum and Burkholderia strains and their effect on rice crop yield and nutrient uptake. Plant Soil 2013, 369, 115–129. [Google Scholar] [CrossRef]
- Defez, R.; Andreozzi, A.; Bianco, C. The overproduction of indole-3-acetic acid (IAA) in endophytes upregulates nitrogen fixation in both bacterial cultures and inoculated rice plants. Microb. Ecol. 2017, 74, 441–452. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.-B.; Du, Z.; Bai, L.; Tian, C.; Zhang, Y.; Xie, J.-Y.; Wang, T.; Liu, X.; Chen, X.; Cheng, Q.; et al. Comparative genomic analysis of N2-fixing and non-N2-fixing Paenibacillus spp.: Organization, evolution and expression of the nitrogen fixation genes. PLoS Genet. 2014, 10, e1004231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarikhani, M.R.; Oustan, S.; Ebrahimi, M.; Aliasgharzad, N. Isolati Isolation and identification of potassium-releasing bacteria in soil and assessment of their ability to release potassium for plants. Eur. J. Soil Sci. 2018, 69, 1078–1086. [Google Scholar] [CrossRef]
- Sun, F.; Ou, Q.; Wang, N.; Guo, Z.X.; Ou, Y.; Li, N.; Peng, C. Isolation and identification of potassium-solubilizing bacteria from Mikania micrantha rhizospheric soil and their effect on M. micrantha plants. Glob. Ecol. Conserv. 2020, 23, e01141. [Google Scholar] [CrossRef]
- Yaghoubi Khanghahi, M.; Pirdashti, H.; Rahimian, H.; Nematzadeh, G.; Sepanlou, M.G. Potassium solubilising bacteria (KSB) isolated from rice paddy soil: From isolation, identification to K use efficiency. Symbiosis 2017, 76, 13–23. [Google Scholar] [CrossRef]
- Jiang, H.; Li, S.; Wang, T.; Chi, X.; Qi, P.; Chen, G. Interaction between halotolerant phosphate-solubilizing bacteria (Providencia rettgeri Strain TPM23) and rock phosphate improves soil biochemical properties and peanut growth in saline soil. Front. Microbiol. 2021, 12, 777351. [Google Scholar] [CrossRef]
- Ji, C.; Liu, Z.; Hao, L.; Song, X.; Wang, C.; Liu, Y.; Li, H.; Li, C.; Gao, Q.; Liu, X. Effects of Enterobacter cloacae HG-1 on the nitrogen-fixing community structure of wheat rhizosphere soil and on salt tolerance. Front. Plant Sci. 2020, 11, 1094. [Google Scholar] [CrossRef]
- Allito, B.B.; Ewusi-Mensah, N.; Logah, V. Legume-rhizobium strain specificity enhances nutrition and nitrogen fixation in faba bean (Vicia faba L.). Agronomy 2020, 10, 826. [Google Scholar] [CrossRef]
- Martins, M.R.; Jantalia, C.P.; Reis, V.M.; Döwich, I.; Polidoro, J.C.; Alves, B.J.R.; Boddey, R.M.; Urquiaga, S. Impact of plant growth-promoting bacteria on grain yield, protein content, and urea-15 N recovery by maize in a Cerrado Oxisol. Plant Soil 2018, 422, 239–250. [Google Scholar] [CrossRef] [Green Version]
- Lu, P.; Liu, H.; Liu, A. Biodegradation of dicofol by Microbacterium sp. D-2 isolated from pesticide-contaminated agricultural soil. Appl. Biol. Chem. 2019, 62, 72. [Google Scholar] [CrossRef] [Green Version]
- Hongsawat, P.; Vangnai, A.S. Biodegradation pathways of chloroanilines by Acinetobacter baylyi strain GFJ2. J. Hazard. Mater. 2011, 186, 1300–1307. [Google Scholar] [CrossRef]
- Kamanavalli, C.M.; Ninnekar, H.Z. Biodegradation of DDT by a Pseudomonas species. Curr. Microbiol. 2004, 48, 10–13. [Google Scholar]
- Chanika, E.; Georgiadou, D.; Soueref, E.; Karas, P.; Karanasios, E.; Tsiropoulos, N.G.; Tzortzakakis, E.A.; Karpouzas, D.G. Isolation of soil bacteria able to hydrolyze both organophosphate and carbamate pesticides. Bioresour. Technol. 2011, 102, 3184–3192. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Li, J.; Zhang, Y.; Wu, X.; Zhou, Z.; Huang, Y.; Zhao, Y.; Mishra, S.; Bhatt, P.; Chen, S. Characterization of a novel glyphosate-degrading bacterial species, Chryseobacterium sp. Y16C, and evaluation of its effects on microbial communities in glyphosate-contaminated soil. J. Hazard. Mater. 2022, 432, 128689. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Song, P.; Wang, G.; Wang, J.; Zhu, L.; Wang, J. Responses of atrazine degradation and native bacterial community in soil to Arthrobacter sp. strain HB-5. Ecotoxicol. Environ. Saf. 2018, 159, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Karuppiah, V.; Li, Y.; Pandian, S.; Kumaran, S.; Chen, J. Role of cytochrome P450 genes of Trichoderma atroviride T23 on the resistance and degradation of dichlorvos. Chemosphere 2022, 290, 133173. [Google Scholar] [CrossRef]
- Yadav, U.; Kushwaha, S.; Anand, V.; Kumar, S.; Prakash, O.; Singh, P.C. Chlorpyrifos degradation by plant growth-promoting Alcaligenes faecalis bacteria isolated from oil-contaminated soil. Bioremediat. J. 2021, 25, 80–88. [Google Scholar] [CrossRef]
- Ali, M.; Naqvi, T.A.; Kanwal, M.; Rasheed, F.; Hameed, A.; Ahmed, S. Detection of the organophosphate degrading gene opdA in the newly isolated bacterial strain Bacillus pumilus W1. Ann. Microbiol. 2012, 62, 233–239. [Google Scholar] [CrossRef]
- Aswathi, A.; Pandey, A.; Sukumaran, R.K. Rapid degradation of the organophosphate pesticide-Chlorpyrifos by a novel strain of Pseudomonas nitroreducens AR-3. Bioresour. Technol. 2019, 292, 122025. [Google Scholar] [CrossRef]
- Loh, K.; Wu, T. Cometabolic transformation of 2-Chlorophenol and 4-Chlorophenol in the presence of phenol by Pseudomonas putida. Can. J. Chem. Eng. 2006, 84, 356–367. [Google Scholar] [CrossRef]
- Dash, D.M.; Osborne, J.W. Biodegradation of monocrotophos by a plant growth promoting Bacillus aryabhattai (VITNNDJ5) strain in artificially contaminated soil. Int. J. Environ. Sci. Technol. 2020, 17, 1475–1490. [Google Scholar] [CrossRef]
- Li, R.; Dörfler, U.; Munch, J.C.; Schroll, R. Enhanced degradation of isoproturon in an agricultural soil by a Sphingomonas sp. strain and a microbial consortium. Chemosphere 2017, 168, 1169–1176. [Google Scholar] [CrossRef] [PubMed]
- Muendo, B.M.; Shikuku, V.O.; Lalah, J.O.; Getenga, Z.M.; Wandiga, S.O.; Rothballer, M. Enhanced degradation of diuron by two Bacillus species isolated from diuron contaminated sugarcane and pineapple-cultivated soils in Kenya. Appl. Soil Ecol. 2021, 157, 103721. [Google Scholar] [CrossRef]
- Zhao, H.; Geng, Y.; Chen, L.; Tao, K.; Hou, T. Biodegradation of cypermethrin by a novel Catellibacterium sp. strain CC-5 isolated from contaminated soil. Can. J. Microbiol. 2013, 59, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, K.S. Environmental contaminant 2-chloro-N-(2,6-diethylphenyl)-N-(methoxymethyl)acetamide remediation via Xanthomonas axonopodis and Aspergillus niger. Environ. Res. 2020, 182, 109117. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhang, Y.; Hou, Z.; Wang, X.; Wang, J.; Lu, Z.; Zhao, X.; Sun, F.; Pan, H. Biodegradation potential of deltamethrin by the Bacillus cereus strain Y1 in both culture and contaminated soil. Int. Biodeterior. Biodegrad. 2016, 106, 53–59. [Google Scholar] [CrossRef]
- Altowayti, W.; Algaifi, H.A.; Bakar, S.A.; Shahir, S. The adsorptive removal of As (III) using biomass of arsenic resistant Bacillus thuringiensis strain WS3: Characteristics and modelling studies. Ecotoxicol. Environ. Saf. 2019, 172, 176–185. [Google Scholar] [CrossRef]
- Afegbua, S.L.; Batty, L.C. Effect of plant growth promoting bacterium; Pseudomonas putida UW4 inoculation on phytoremediation efficacy of monoculture and mixed culture of selected plant species for PAH and lead spiked soils. Int. J. Phytoremediat. 2019, 21, 200–208. [Google Scholar] [CrossRef]
- Hussain, A.; Amna; Kamran, M.A.; Javed, M.T.; Hayat, K.; Farooq, M.A.; Ali, N.; Ali, M.; Manghwar, H.; Jan, F.; et al. Individual and combinatorial application of Kocuria rhizophila and citric acid on phytoextraction of multi-metal contaminated soils by Glycine max L. Environ. Exp. Bot. 2019, 159, 23–33. [Google Scholar] [CrossRef]
- Huang, H.; Zhao, Y.; Fan, L.; Jin, Q.; Yang, G.; Xu, Z. Improvement of manganese phytoremediation by Broussonetia papyrifera with two plant growth promoting (PGP) Bacillus species. Chemosphere 2020, 260, 127614. [Google Scholar] [CrossRef]
- Kumar, A.; Voropaeva, O.; Maleva, M.; Panikovskaya, K.; Borisova, G.; Rajkumar, M.; Bruno, L.B. Bioaugmentation with copper tolerant endophyte Pseudomonas lurida strain EOO26 for improved plant growth and copper phytoremediation by Helianthus annuus. Chemosphere 2021, 266, 128983. [Google Scholar] [CrossRef]
- Wani, P.A.; Wahid, S.; Singh, R.; Kehinde, A.M. Antioxidant and chromium reductase assisted chromium (VI) reduction and Cr (III) immobilization by the rhizospheric Bacillus helps in the remediation of Cr (VI) and growth promotion of soybean crop. Rhizosphere 2018, 6, 23–30. [Google Scholar] [CrossRef]
- Haider, F.U.; Coulter, J.A.; Cheema, S.A.; Farooq, M.; Wu, J.; Zhang, R.; Shuaijie, G.; Liqun, C. Co-application of biochar and microorganisms improves soybean performance and remediate cadmium-contaminated soil. Ecotoxicol. Environ. Saf. 2021, 214, 112112. [Google Scholar] [CrossRef] [PubMed]
- Qadir, M.; Hussain, A.; Shah, M.; Lee, I.J.; Iqbal, A.; Irshad, M.; Ismail; Sayyed, A.; Husna; Ahmad, A.; et al. Comparative assessment of chromate bioremediation potential of Pantoea conspicua and Aspergillus niger. J. Hazard. Mater. 2022, 424, 127314. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Guo, H.; He, Z.; Yang, C.; Yu, T.; Chai, Q.; Lu, L. Interaction of Lolium perenne and Hyphomicrobium sp. GHH enhances the removal of 17α-ethinyestradiol (EE2) from soil. J. Soils Sediments 2018, 19, 1297–1305. [Google Scholar] [CrossRef]
- Ping, L.; Zhang, C.; Zhang, C.; Zhu, Y.; He, H.; Wu, M.; Tang, T.; Li, Z.; Zhao, H. Isolation and characterization of pyrene and benzo[a]pyrene-degrading Klebsiella pneumonia PL1 and its potential use in bioremediation. Appl. Microbiol. Biotechnol. 2014, 98, 3819–3828. [Google Scholar] [CrossRef] [PubMed]
- Smułek, W.; Sydow, M.; Zabielska-Matejuk, J.; Kaczorek, E. Bacteria involved in biodegradation of creosote PAH-A case study of long-term contaminated industrial area. Ecotoxicol. Environ. Saf. 2020, 187, 109843. [Google Scholar] [CrossRef]
- Masakorala, K.; Yao, J.; Cai, M.; Chandankere, R.; Yuan, H.; Chen, H. Isolation and characterization of a novel phenanthrene (PHE) degrading strain Psuedomonas sp. USTB-RU from petroleum contaminated soil. J. Hazard. Mater. 2013, 263, 2493–2500. [Google Scholar] [CrossRef]
- Cunliffe, M.; Kertesz, M.A. Effect of Sphingobium yanoikuyae B1 inoculation on bacterial community dynamics and polycyclic aromatic hydrocarbon degradation in aged and freshly PAH-contaminated soils. Environ. Pollut. 2006, 144, 228–237. [Google Scholar] [CrossRef]
- Wang, T.; Su, D.; Wang, X.; He, Z. Adsorption-degradation of polycyclic aromatic hydrocarbons in soil by immobilized mixed bacteria and its effect on microbial communities. J. Agric. Food Chem. 2020, 68, 14907–14916. [Google Scholar] [CrossRef]
- DeBruyn, J.M.; Mead, T.J.; Sayler, G.S. Horizontal transfer of PAH catabolism genes in Mycobacterium: Evidence from comparative genomics and isolated pyrene-degrading bacteria. Environ. Sci. Technol. 2012, 46, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Dionisi, H.M.; Chewning, C.S.; Morgan, K.H.; Menn, F.M.; Easter, J.P.; Sayler, G.S. Abundance of dioxygenase genes similar to Ralstonia sp. strain U2 nagAc is correlated with naphthalene concentrations in coal tar-contaminated freshwater sediments. Appl. Environ. Microbiol. 2004, 70, 3988–3995. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.; Singh, S.N. Biodegradation of benzo(a)pyrene mediated by catabolic enzymes of bacteria. Int. J. Environ. Sci. Technol. 2013, 11, 1571–1580. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.Y.; Chen, D.Z.; Zhu, R.Y.; Chen, J.M. Substrate interactions during the biodegradation of BTEX and THF mixtures by Pseudomonas oleovorans DT4. Bioresour. Technol. 2011, 102, 6644–6649. [Google Scholar] [CrossRef]
- Wu, X.; Gu, Y.; Wu, X.; Zhou, X.; Zhou, H.; Amanze, C.; Shen, L.; Zeng, W. Construction of a tetracycline degrading bacterial consortium and its application evaluation in laboratory-scale soil remediation. Microorganisms 2020, 8, 292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, B.; Ju, X.; Chang, J.; Ge, Y.; Vitousek, P.M. Integrated reactive nitrogen budgets and future trends in China. Proc. Natl. Acad. Sci. USA 2015, 112, 8792–8797. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; He, Y.; Zhou, W.; Ai, L.; Liu, H.; Chen, L.; Xie, Y. Effects of continuous cropping of Codonopsis tangshen on rhizospheric soil bacterial community as determined by pyrosequencing. Diversity 2021, 13, 317. [Google Scholar] [CrossRef]
- Ling, D.; Huang, Q.; Ouyang, Y. Impacts of simulated acid rain on soil enzyme activities in a latosol. Ecotoxicol. Environ. Saf. 2010, 73, 1914–1918. [Google Scholar] [CrossRef]
- Coban, O.; De Deyn, G.B.; van der Ploeg, M. Soil microbiota as game-changers in restoration of degraded lands. Science 2022, 375, abe0725. [Google Scholar] [CrossRef]
- Majeed, A.; Muhammad, Z.; Ahmad, H. Plant growth promoting bacteria: Role in soil improvement, abiotic and biotic stress management of crops. Plant Cell Rep. 2018, 37, 1599–1609. [Google Scholar] [CrossRef]
- Liang, C.; Amelung, W.; Lehmann, J.; Kastner, M. Quantitative assessment of microbial necromass contribution to soil organic matter. Glob. Chang. Biol. 2019, 25, 3578–3590. [Google Scholar] [CrossRef] [PubMed]
- Shahrajabian, M.H.; Sun, W.; Cheng, Q. The importance of Rhizobium, Agrobacterium, Bradyrhizobium, Herbaspirillum, Sinorhizobium in sustainable agricultural production. Not. Bot. Horti Agrobot. 2021, 49, 12183. [Google Scholar] [CrossRef]
- Taha, K.; Berraho, E.B.; El Attar, I.; Dekkiche, S.; Aurag, J.; Béna, G. Rhizobium laguerreae is the main nitrogen-fixing symbiont of cultivated lentil (Lens culinaris) in Morocco. Syst. Appl. Microbiol. 2018, 41, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, J.T.; Hu, Y.; Wiig, J.A.; Rees, D.C.; Ribbe, M.W. Structure of precursor-bound NifEN: A nitrogenase FeMo cofactor maturase/insertase. Science 2011, 331, 91–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nonaka, A.; Yamamoto, H.; Kamiya, N.; Kotani, H.; Yamakawa, H.; Tsujimoto, R.; Fujita, Y. Accessory proteins of the nitrogenase assembly, NifW, NifX/NafY, and NifZ, are essential for diazotrophic growth in the nonheterocystous cyanobacterium Leptolyngbya boryana. Front. Microbiol. 2019, 10, 495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fay, A.W.; Blank, M.A.; Rebelein, J.G.; Lee, C.C.; Ribbe, M.W.; Hedman, B.; Hodgson, K.O.; Hu, Y. Assembly scaffold NifEN: A structural and functional homolog of the nitrogenase catalytic component. Proc. Natl. Acad. Sci. USA 2016, 113, 9504–9508. [Google Scholar] [CrossRef] [Green Version]
- Alemneh, A.A.; Zhou, Y.; Ryder, M.H.; Denton, M.D. Mechanisms in plant growth-promoting rhizobacteria that enhance legume-rhizobial symbioses. J. Appl. Microbiol 2020, 129, 1133–1156. [Google Scholar] [CrossRef]
- Lodwig, E.M.; Hosie, A.H.; Bourdes, A.; Findlay, K.; Allaway, D.; Karunakaran, R.; Downie, J.A.; Poole, P.S. Amino-acid cycling drives nitrogen fixation in the legume-Rhizobium symbiosis. Nature 2003, 422, 722–726. [Google Scholar] [CrossRef]
- Lindstrom, K.; Mousavi, S.A. Effectiveness of nitrogen fixation in rhizobia. Microb. Biotechnol. 2020, 13, 1314–1335. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.-J.; Radhakrishnan, R.; Kang, S.-M.; You, Y.-H.; Jeong, E.-J.; Kim, J.-G.; Lee, I.-J. Plant growth promoting effect of Bacillus amyloliquefaciens H-2-5 on crop plants and influence on physiological changes in soybean under soil salinity. Physiol. Mol. Biol. Plants 2017, 23, 571–580. [Google Scholar] [CrossRef]
- Wu, X.; Rensing, C.; Han, D.; Xiao, K.Q.; Dai, Y.; Tang, Z.; Liesack, W.; Peng, J.; Cui, Z.; Zhang, F. Genome-resolved metagenomics reveals distinct phosphorus acquisition strategies between soil microbiomes. mSystems 2022, 7, e0110721. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Cui, Z.; Peng, J.; Zhang, F.; Liesack, W. Genome-resolved metagenomics identifies the particular genetic traits of phosphate-solubilizing bacteria in agricultural soil. ISME Commun. 2022, 2, 17. [Google Scholar] [CrossRef]
- Tewari, S.; Pooniya, V.; Sharma, S. Next generation bioformulation prepared by amalgamating Bradyrhizobium, cell free culture supernatant, and exopolysaccharides enhances the indigenous rhizospheric rhizobial population, nodulation, and productivity of pigeon pea. Appl. Soil Ecol. 2020, 147, 103363. [Google Scholar] [CrossRef]
- Adnan, M.; Shah, Z.; Fahad, S.; Arif, M.; Alam, M.; Khan, I.A.; Mian, I.A.; Basir, A.; Ullah, H.; Arshad, M.; et al. Phosphate-solubilizing bacteria nullify the antagonistic effect of soil calcification on bioavailability of phosphorus in alkaline soils. Sci. Rep. 2017, 7, 16131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, S.; Feng, B.; Xiao, C.; Wang, Q.; Chi, R. Phosphate-solubilizing microorganisms to enhance phytoremediation of excess phosphorus pollution in phosphate mining wasteland soil. Bioremediat. J. 2021, 25, 271–281. [Google Scholar] [CrossRef]
- Saeed, Q.; Wang, X.; Haider, F.U.; Kucerik, J.; Mumtaz, M.Z.; Holatko, J.; Naseem, M.; Kintl, A.; Ejaz, M.; Naveed, M.; et al. Rhizosphere bacteria in plant growth promotion, biocontrol, and bioremediation of contaminated sites: A comprehensive review of effects and mechanisms. Int. J. Mol. Sci. 2021, 22, 10529. [Google Scholar] [CrossRef]
- Zheng, W.; Morris, E.K.; Lehmann, A.; Rillig, M.C. Interplay of soil water repellency, soil aggregation and organic carbon. A meta-analysis. Geoderma 2016, 283, 39–47. [Google Scholar] [CrossRef]
- Rashid, M.I.; Mujawar, L.H.; Shahzad, T.; Almeelbi, T.; Ismail, I.M.; Oves, M. Bacteria and fungi can contribute to nutrients bioavailability and aggregate formation in degraded soils. Microbiol. Res. 2016, 183, 26–41. [Google Scholar] [CrossRef]
- Caesar-TonThat, T.C.; Stevens, W.B.; Sainju, U.M.; Caesar, A.J.; West, M.; Gaskin, J.F. Soil-aggregating bacterial community as affected by irrigation, tillage, and cropping system in the Northern Great Plains. Soil Sci. 2014, 179, 11–20. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.; He, X.; Zou, Y.; He, K.; Sun, Y.; Cao, M. Spatial distribution of glomalin-related soil protein and its relationships with root mycorrhization, soil aggregates, carbohydrates, activity of protease and β-glucosidase in the rhizosphere of Citrus unshiu. Soil Biol. Biochem. 2012, 45, 181–183. [Google Scholar] [CrossRef]
- Henao, L.J.; Mazeau, K. Molecular modelling studies of clay-exopolysaccharide complexes: Soil aggregation and water retention phenomena. Mater. Sci. Eng. C 2009, 29, 2326–2332. [Google Scholar] [CrossRef]
- Kallenbach, C.M.; Frey, S.D.; Grandy, A.S. Direct evidence for microbial-derived soil organic matter formation and its ecophysiological controls. Nat. Commun. 2016, 7, 13630. [Google Scholar] [CrossRef] [Green Version]
- Six, J.; Bossuyt, H.; Degryze, S.; Denef, K. A history of research on the link between (micro)aggregates, soil biota, and soil organic matter dynamics. Soil Tillage Res. 2004, 79, 7–31. [Google Scholar] [CrossRef]
- Yang, Y.; Xie, H.; Mao, Z.; Bao, X.; He, H.; Zhang, X.; Liang, C. Fungi determine increased soil organic carbon more than bacteria through their necromass inputs in conservation tillage croplands. Soil Biol. Biochem. 2022, 167, 108587. [Google Scholar] [CrossRef]
- Xie, M.; Zhang, S.; Cui, Z.; Cao, X. Distribution characteristics and risk assessment of polycyclic aromatic hydrocarbons in soils of a steel enterprise in East China. Bull. Environ. Contam. Toxicol. 2021, 106, 873–877. [Google Scholar] [CrossRef]
- Dar, M.A.; Kaushik, G.; Villarreal-Chiu, J.F. Pollution status and bioremediation of chlorpyrifos in environmental matrices by the application of bacterial communities: A review. J. Environ. Manag. 2019, 239, 124–136. [Google Scholar] [CrossRef]
- Bhadbhade, B.J.; Sarnaik, S.S.; Kanekar, P.P. Biomineralization of an organophosphorus pesticide, Monocrotophos, by soil bacteria. J. Appl. Microbiol. 2002, 93, 224–234. [Google Scholar] [CrossRef]
- Abraham, J.; Silambarasan, S.; Logeswari, P. Simultaneous degradation of organophosphorus and organochlorine pesticides by bacterial consortium. J. Taiwan Inst. Chem. Eng. 2014, 45, 2590–2596. [Google Scholar] [CrossRef]
- Lovecka, P.; Pacovska, I.; Stursa, P.; Vrchotova, B.; Kochankova, L.; Demnerova, K. Organochlorinated pesticide degrading microorganisms isolated from contaminated soil. New Biotechnol. 2015, 32, 26–31. [Google Scholar] [CrossRef]
- Renault, H.; Bassard, J.E.; Hamberger, B.; Werck-Reichhart, D. Cytochrome P450-mediated metabolic engineering: Current progress and future challenges. Curr. Opin. Plant Biol. 2014, 19, 27–34. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Huo, X.; Zhou, J.; Zhang, Q.; Wang, W. Metabolic activation mechanism of 2,2′,3,3′,6,6′-hexachlorobiphenyl (PCB136) by cytochrome P450 2B6: A QM/MM approach. Sci. Total Environ. 2021, 776, 145579. [Google Scholar] [CrossRef] [PubMed]
- Chadha, S.; Mehetre, S.T.; Bansal, R.; Kuo, A.; Aerts, A.; Grigoriev, I.V.; Druzhinina, I.S.; Mukherjee, P.K. Genome-wide analysis of cytochrome P450s of Trichoderma spp.: Annotation and evolutionary relationships. Fungal Biol. Biotechnol. 2018, 5, 12. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Lin, Z.; Pang, S.; Bhatt, P.; Chen, S. Insights into the biodegradation of lindane (gamma-Hexachlorocyclohexane) using a microbial system. Front. Microbiol. 2020, 11, 522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Debruyn, J.M.; Chewning, C.S.; Sayler, G.S. Comparative quantitative prevalence of Mycobacteria and functionally abundant nidA, nahAc, and nagAc dioxygenase genes in coal tar contaminated sediments. Environ. Sci. Technol. 2007, 41, 5426–5432. [Google Scholar] [CrossRef] [PubMed]
- Nzila, A. Update on the cometabolism of organic pollutants by bacteria. Environ. Pollut. 2013, 178, 474–482. [Google Scholar] [CrossRef] [PubMed]
- Aktas, O.; Cecen, F. Cometabolic bioregeneration of activated carbons loaded with 2-chlorophenol. Bioresour. Technol. 2009, 100, 4604–4610. [Google Scholar] [CrossRef]
- Gu, D.; Xiang, X.; Wu, Y.; Zeng, J.; Lin, X. Synergy between fungi and bacteria promotes polycyclic aromatic hydrocarbon cometabolism in lignin-amended soil. J. Hazard. Mater. 2022, 425, 127958. [Google Scholar] [CrossRef]
- Zulfiqar, U.; Farooq, M.; Hussain, S.; Maqsood, M.; Hussain, M.; Ishfaq, M.; Ahmad, M.; Anjum, M.Z. Lead toxicity in plants: Impacts and remediation. J. Environ. Manag. 2019, 250, 109557. [Google Scholar] [CrossRef]
- Redha, A.; Al-Hasan, R.; Afzal, M. Synergistic and concentration-dependent toxicity of multiple heavy metals compared with single heavy metals in Conocarpus lancifolius. Environ. Sci. Pollut. Res. Int. 2021, 28, 23258–23272. [Google Scholar] [CrossRef]
- Huang, X.H.; Zhu, F.; Yan, W.D.; Chen, X.Y.; Wang, G.J.; Wang, R.J. Effects of Pb and Zn toxicity on chlorophyll fluorescence and biomass production of Koelreuteria paniculata and Zelkova schneideriana young plants. Photosynthetica 2019, 57, 688–697. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.S.; Kormoker, T.; Idris, A.M.; Proshad, R.; Kabir, M.H.; Ustaoğlu, F. Plant-microbe-metal interactions for heavy metal bioremediation: A review. Crop Pasture Sci. 2021, 73, 181–201. [Google Scholar] [CrossRef]
- Khan, I.; Ali, M.; Aftab, M.; Shakir, S.; Qayyum, S.; Haleem, K.S.; Tauseef, I. Mycoremediation: A treatment for heavy metal-polluted soil using indigenous metallotolerant fungi. Environ. Monit. Assess. 2019, 191, 622. [Google Scholar] [CrossRef]
- Fakhar, A.; Gul, B.; Gurmani, A.R.; Khan, S.M.; Ali, S.; Sultan, T.; Chaudhary, H.J. Heavy metal remediation and resistance mechanism of Aeromonas, Bacillus, and Pseudomonas: A review. Crit. Rev. Environ. Sci. Technol. 2022, 52, 1868–1914. [Google Scholar] [CrossRef]
- Brady, D.; Duncan, J.R. Cation loss during accumulation of heavy metal cations by Saccharomyces cerevisiae. Biotechnol. Lett. 1994, 16, 543–548. [Google Scholar] [CrossRef]
- Khalid, M.; Ur-Rahman, S.; Hassani, D.; Hayat, K.; Pei, Z.; Nan, H. Advances in fungal-assisted phytoremediation of heavy metals; a review. Pedosphere 2021, 31, 475–495. [Google Scholar] [CrossRef]
- Dong, L.; Xu, J.; Zhang, L.; Cheng, R.; Wei, G.; Su, H.; Yang, J.; Qian, J.; Xu, R.; Chen, S. Rhizospheric microbial communities are driven by Panax ginseng at different growth stages and biocontrol bacteria alleviates replanting mortality. Acta Pharm. Sin. B 2018, 8, 272–282. [Google Scholar] [CrossRef]
- Sun, Z.; Yang, L.; Han, M.; Han, Z.; Yang, L.; Cheng, L.; Yang, X.; Lv, Z.-L. Biological control ginseng grey mold and plant colonization by antagonistic bacteria isolated from rhizospheric soil of Panax ginseng Meyer. Biol. Control 2019, 138, 104048. [Google Scholar] [CrossRef]
- Lee, B.D.; Dutta, S.; Ryu, H.; Yoo, S.J.; Suh, D.S.; Park, K. Induction of systemic resistance in Panax ginseng against Phytophthora cactorum by native Bacillus amyloliquefaciens HK34. J. Ginseng Res. 2015, 39, 213–220. [Google Scholar] [CrossRef] [Green Version]
- Durairaj, K.; Velmurugan, P.; Park, J.; Chang, W.; Park, Y.; Senthilkumar, P.; Choi, K.-M.; Lee, J.-H.; Oh, B.-T. An investigation of biocontrol activity Pseudomonas and Bacillus strains against Panax ginseng root rot fungal phytopathogens. Biol. Control 2018, 125, 138–146. [Google Scholar] [CrossRef]
- Kang, J.P.; Huo, Y.; Yang, D.U.; Yang, D.C. Influence of the plant growth promoting Rhizobium panacihumi on aluminum resistance in Panax ginseng. J. Ginseng Res. 2021, 45, 442–449. [Google Scholar] [CrossRef]
- Dong, L.; Xu, J.; Li, Y.; Fang, H.; Niu, W.; Li, X.; Zhang, Y.; Ding, W.; Chen, S. Manipulation of microbial community in the rhizosphere alleviates the replanting issues in Panax ginseng. Soil Biol. Biochem. 2018, 125, 64–74. [Google Scholar] [CrossRef]
- Liu, N.; Shao, C.; Sun, H.; Liu, Z.; Guan, Y.; Wu, L.; Zhang, L.; Pan, X.; Zhang, Z.; Zhang, Y.; et al. Arbuscular mycorrhizal fungi biofertilizer improves American ginseng (Panax quinquefolius L.) growth under the continuous cropping regime. Geoderma 2020, 363, 114155. [Google Scholar] [CrossRef]
- Amanifar, S.; Khodabandeloo, M.; Mohsenifard, E.; Askari, M.S.; Ashrafi, M. Alleviation of salt stress and changes in glycyrrhizin accumulation by arbuscular mycorrhiza in liquorice (Glycyrrhiza glabra) grown under salinity stress. Environ. Exp. Bot. 2019, 160, 25–34. [Google Scholar] [CrossRef]
- Liu, J.; Wu, L.; Wei, S.; Xiao, X.; Su, C.; Jiang, P.; Song, Z.; Wang, T.; Yu, Z. Effects of arbuscular mycorrhizal fungi on the growth, nutrient uptake and glycyrrhizin production of licorice (Glycyrrhiza uralensis Fisch). Plant Growth Regul. 2007, 52, 29–39. [Google Scholar] [CrossRef]
- Yu, M.; Xie, W.; Zhang, X.; Zhang, S.; Wang, Y.; Hao, Z.; Chen, B. Arbuscular mycorrhizal fungi can compensate for the loss of indigenous microbial communities to support the growth of liquorice (Glycyrrhiza uralensis Fisch.). Plants 2019, 9, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, C.; Wang, W.; Hou, J. Plant growth and soil microbial impacts of enhancing licorice with inoculating dark septate endophytes under drought stress. Front. Microbiol. 2019, 10, 2277. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Zhou, L.; Wu, J. Promotion of Salvia miltiorrhiza hairy root growth and tanshinone production by polysaccharide-protein fractions of plant growth-promoting rhizobacterium Bacillus cereus. Process Biochem. 2010, 45, 1517–1522. [Google Scholar] [CrossRef]
- Wei, X.; Cao, P.; Wang, G.; Han, J. Microbial inoculant and garbage enzyme reduced cadmium (Cd) uptake in Salvia miltiorrhiza (Bge.) under Cd stress. Ecotoxicol. Environ. Saf. 2020, 192, 110311. [Google Scholar] [CrossRef]
- Geneva, M.P.; Stancheva, I.V.; Boychinova, M.M.; Mincheva, N.H.; Yonova, P.A. Effects of foliar fertilization and arbuscular mycorrhizal colonization on Salvia officinalis L. growth, antioxidant capacity, and essential oil composition. J. Sci. Food Agric. 2010, 90, 696–702. [Google Scholar] [CrossRef]
- Wang, J.W.; Zhang, Z.; Tan, R.X. Stimulation of artemisinin production in Artemisia annua hairy roots by the elicitor from the endophytic Colletotrichum sp. Biotechnol. Lett. 2001, 23, 857–860. [Google Scholar] [CrossRef]
- Domokos, E.; Bíró-Janka, B.; Bálint, J.; Molnár, K.; Fazakas, C.; Jakab-Farkas, L.; Domokos, J.; Albert, C.; Mara, G.; Balog, A. Arbuscular mycorrhizal fungus rhizophagus irregularis influences Artemisia annua plant parameters and artemisinin content under different soil types and cultivation methods. Microorganisms 2020, 8, 899. [Google Scholar] [CrossRef] [PubMed]
- Awasthi, A.; Bharti, N.; Nair, P.; Singh, R.; Shukla, A.K.; Gupta, M.M.; Darokar, M.P.; Kalra, A. Synergistic effect of Glomus mosseae and nitrogen fixing Bacillus subtilis strain Daz26 on artemisinin content in Artemisia annua L. Appl. Soil Ecol. 2011, 49, 125–130. [Google Scholar] [CrossRef]
- Chen, J.; Li, N.; Chang, J.; Ren, K.; Zhou, J.; Yang, G. Taxonomic structure of rhizosphere bacterial communities and its association with the accumulation of alkaloidal metabolites in Sophora flavescens. Front. Microbiol. 2021, 12, 781316. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Meng, X.; Jiao, Y.; Guo, B.; Sui, X.; Ma, S.; Chen, W.; Singh, R. Bradyrhizobium arachidis mediated enhancement of (oxy)matrine content in the medicinal legume Sophora flavescens. Lett. Appl. Microbiol. 2021, 72, 570–577. [Google Scholar] [CrossRef]
- Khalilpour, M.; Mozafari, V.; Abbaszadeh-Dahaji, P. Tolerance to salinity and drought stresses in pistachio (Pistacia vera L.) seedlings inoculated with indigenous stress-tolerant PGPR isolates. Sci. Hortic. 2021, 289, 110440. [Google Scholar] [CrossRef]
- Pankaj, U.; Singh, D.N.; Mishra, P.; Gaur, P.; Babu, C.S.V.; Shanker, K.; Verma, R.K. Autochthonous halotolerant plant growth-promoting rhizobacteria promote bacoside A yield of Bacopa monnieri (L.) Nash and phytoextraction of salt-affected soil. Pedosphere 2020, 30, 671–683. [Google Scholar] [CrossRef]
- Attarzadeh, M.; Balouchi, H.; Rajaie, M.; Movahhedi, D.M.; Salehi, A. Growth and nutrient content of Echinacea purpurea as affected by the combination of phosphorus with arbuscular mycorrhizal fungus and Pseudomonas florescent bacterium under different irrigation regimes. J. Environ. Manag. 2019, 231, 182–188. [Google Scholar] [CrossRef]
- Barnawal, D.; Maji, D.; Bharti, N.; Chanotiya, C.S.; Kalra, A. ACC deaminase-containing Bacillus subtilis reduces stress ethylene-induced damage and improves mycorrhizal colonization and rhizobial nodulation in Trigonella foenum-graecum under drought stress. J. Plant Growth Regul. 2013, 32, 809–822. [Google Scholar] [CrossRef]
- Dadrasan, M.; Chaichi, M.R.; Pourbabaee, A.A.; Yazdani, D.; Keshavarz-Afshar, R. Deficit irrigation and biological fertilizer influence on yield and trigonelline production of fenugreek. Ind. Crop. Prod. 2015, 77, 156–162. [Google Scholar] [CrossRef]
- Mishra, B.K.; Meena, K.K.; Dubey, P.N.; Aishwath, O.P.; Kant, K.; Sorty, A.M.; Bitla, U. Influence on yield and quality of fennel (Foeniculum vulgare Mill.) grown under semi-arid saline soil, due to application of native phosphate solubilizing rhizobacterial isolates. Ecol. Eng. 2016, 97, 327–333. [Google Scholar] [CrossRef]
- Tian, Y.; Liu, Y.; Yue, L.; Uwaremwe, C.; Zhao, X.; Zhou, Q.; Wang, Y.; Wang, R. Bacterial inoculant and sucrose amendments improve the growth of Rheum palmatum L. by reprograming its metabolite composition and altering its soil microbial community. Int. J. Mol. Sci. 2022, 23, 1694. [Google Scholar] [CrossRef] [PubMed]
- Benson, A.; Joe, M.M.; Karthikeyan, B.; Sa, T.; Rajasekaran, C. Role of Achromobacter xylosoxidans AUM54 in micropropagation of endangered medicinal plant Naravelia zeylanica (L.) DC. J. Plant Growth Regul. 2014, 33, 202–213. [Google Scholar] [CrossRef]
- Li, Y.; Liu, Y.; Zhang, H.; Yang, Y.; Wei, G.; Li, Z. The composition of root-associated bacteria and fungi of Astragalus mongholicus and their relationship with the bioactive ingredients. Front. Microbiol. 2021, 12, 642730. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zheng, L.; Zheng, Y.; Xue, S.; Zhang, J.; Huang, P.; Zhao, Y.; Hao, X.; He, Z.; Hu, Z.; et al. Insight into the assembly of root-associated microbiome in the medicinal plant Polygonum cuspidatum. Ind. Crop. Prod. 2020, 145, 112163. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, M.; Li, Y.; Wu, A.; Huang, J. Effects of arbuscular mycorrhizal fungi on growth and nitrogen uptake of Chrysanthemum morifolium under salt stress. PLoS ONE 2018, 13, e0196408. [Google Scholar] [CrossRef]
- Barnawal, D.; Pandey, S.S.; Bharti, N.; Pandey, A.; Ray, T.; Singh, S.; Chanotiya, C.S.; Kalra, A. ACC deaminase-containing plant growth-promoting rhizobacteria protect Papaver somniferum from downy mildew. J. Appl. Microbiol. 2017, 122, 1286–1298. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Jiang, Q.; Liu, X.; Liu, L.; Ding, W. Plant growth promoting rhizobacteria alleviate aluminum toxicity and ginger bacterial wilt in acidic continuous cropping soil. Front. Microbiol. 2020, 11, 569512. [Google Scholar] [CrossRef]
- Tripathi, P.; Yadav, R.; Das, P.; Singh, A.; Singh, R.P.; Kandasamy, P.; Kalra, A.; Khare, P. Endophytic bacterium CIMAP-A7 mediated amelioration of atrazine induced phyto-toxicity in Andrographis paniculata. Environ. Pollut. 2021, 287, 117635. [Google Scholar] [CrossRef]
- Khan, K.; Pankaj, U.; Verma, S.K.; Gupta, A.K.; Singh, R.P.; Verma, R.K. Bio-inoculants and vermicompost influence on yield, quality of Andrographis paniculata, and soil properties. Ind. Crop. Prod. 2015, 70, 404–409. [Google Scholar] [CrossRef]
- Lima, C.S.; Santos, H.R.S.; Albuquerque, U.P.D.; Silva, F.S.B.D. Mycorrhizal symbiosis increase the level of total foliar phenols and tannins in Commiphora leptophloeos (Mart.) J.B. Gillett seedlings. Ind. Crop. Prod. 2017, 104, 28–32. [Google Scholar] [CrossRef]
- Toussaint, J.; Smith, F.A.; Smith, S.E. Arbuscular mycorrhizal fungi can induce the production of phytochemicals in sweet basil irrespective of phosphorus nutrition. Mycorrhiza 2007, 17, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.Y.; Sun, K.; Chen, F.; Yuan, J.; Li, X.; Dai, C.C. Endophytic Pseudomonas induces metabolic flux changes that enhance medicinal sesquiterpenoid accumulation in Atractylodes lancea. Plant Physiol. Biochem. 2018, 130, 473–481. [Google Scholar] [CrossRef] [PubMed]
- Mañero, F.J.G.; Algar, E.; Martín Gómez, M.S.; Saco Sierra, M.D.; Solano, B.R. Elicitation of secondary metabolism in Hypericum perforatum by rhizosphere bacteria and derived elicitors in seedlings and shoot cultures. Pharm. Biol. 2012, 50, 1201–1209. [Google Scholar] [CrossRef] [PubMed]
- Zubek, S.; Mielcarek, S.; Turnau, K. Hypericin and pseudohypericin concentrations of a valuable medicinal plant Hypericum perforatum L. are enhanced by arbuscular mycorrhizal fungi. Mycorrhiza 2012, 22, 149–156. [Google Scholar] [CrossRef] [Green Version]
- Ghorbanpour, M. Role of plant growth promoting rhizobacteria on antioxidant enzyme activities and tropane alkaloids production of Hyoscyamus niger under water deficit stress. Turk. J. Biol. 2013, 37, 350–360. [Google Scholar] [CrossRef]
- Jami, N.; Rahimi, A.; Naghizadeh, M.; Sedaghati, E. Investigating the use of different levels of Mycorrhiza and Vermicompost on quantitative and qualitative yield of saffron (Crocus sativus L.). Sci. Hortic. 2020, 262, 109027. [Google Scholar] [CrossRef]
- Bharti, N.; Barnawal, D.; Shukla, S.; Tewari, S.K.; Katiyar, R.S.; Kalra, A. Integrated application of Exiguobacterium oxidotolerans, Glomus fasciculatum, and vermicompost improves growth, yield and quality of Mentha arvensis in salt-stressed soils. Ind. Crop. Prod. 2016, 83, 717–728. [Google Scholar] [CrossRef]
- Singh, S.; Tripathi, A.; Maji, D.; Awasthi, A.; Vajpayee, P.; Kalra, A. Evaluating the potential of combined inoculation of Trichoderma harzianum and Brevibacterium halotolerans for increased growth and oil yield in Mentha arvensis under greenhouse and field conditions. Ind. Crop. Prod. 2019, 131, 173–181. [Google Scholar] [CrossRef]
- Liu, S.; Garcia-Palacios, P.; Tedersoo, L.; Guirado, E.; van der Heijden, M.; Wagg, C.; Chen, D.; Wang, Q.; Wang, J.; Singh, B.K.; et al. Phylotype diversity within soil fungal functional groups drives ecosystem stability. Nat. Ecol. Evol. 2022, 6, 900–909. [Google Scholar] [CrossRef]
- Ling, N.; Wang, T.; Kuzyakov, Y. Rhizosphere bacteriome structure and functions. Nat. Commun. 2022, 13, 836. [Google Scholar] [CrossRef]
- Aslam, M.M.; Okal, E.J.; Idris, A.L.; Qian, Z.; Xu, W.; Karanja, J.K.; Wani, S.H.; Yuan, W. Rhizosphere microbiomes can regulate plant drought tolerance. Pedosphere 2022, 32, 61–74. [Google Scholar] [CrossRef]
- Wang, C.; Li, Y.; Li, M.; Zhang, K.; Ma, W.; Zheng, L.; Xu, H.; Cui, B.; Liu, R.; Yang, Y.; et al. Functional assembly of root-associated microbial consortia improves nutrient efficiency and yield in soybean. J. Integr. Plant Biol. 2021, 63, 1021–1035. [Google Scholar] [CrossRef] [PubMed]
- Oldroyd, G.E.D.; Leyser, O. A plant’s diet, surviving in a variable nutrient environment. Science 2020, 368, eaba0196. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Shao, J.; Fu, Y.; Chen, Y.; Wang, H.; Xu, Z.; Feng, H.; Xun, W.; Liu, Y.; Zhang, N.; et al. The volatile cedrene from Trichoderma guizhouense modulates Arabidopsis root development through auxin transport and signalling. Plant Cell Environ. 2022, 45, 969–984. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Chen, A.; Xie, K.; Yang, X.; Luo, Z.; Chen, J.; Zeng, D.; Ren, Y.; Yang, C.; Wang, L.; et al. Functional analysis of the OsNPF4.5 nitrate transporter reveals a conserved mycorrhizal pathway of nitrogen acquisition in plants. Proc. Natl. Acad. Sci. USA 2020, 117, 16649–16659. [Google Scholar] [CrossRef]
- Chen, C.; Wang, M.; Zhu, J.; Tang, Y.; Zhang, H.; Zhao, Q.; Jing, M.; Chen, Y.; Xu, X.; Jiang, J.; et al. Long-term effect of epigenetic modification in plant-microbe interactions: Modification of DNA methylation induced by plant growth-promoting bacteria mediates promotion process. Microbiome 2022, 10, 36. [Google Scholar] [CrossRef]
- Wu, H.; Qin, X.; Wang, J.; Wu, L.; Chen, J.; Fan, J.; Zheng, L.; Tangtai, H.; Arafat, Y.; Lin, W.; et al. Rhizosphere responses to environmental conditions in Radix pseudostellariae under continuous monoculture regimes. Agric. Ecosyst. Environ. 2019, 270–271, 19–31. [Google Scholar] [CrossRef]
- Wu, H.; Xia, J.; Qin, X.; Wu, H.; Zhang, S.; Zhao, Y.; Rensing, C.; Lin, W. Underlying mechanism of wild Radix pseudostellariae in tolerance to disease under the natural forest cover. Front. Microbiol. 2020, 11, 1142. [Google Scholar] [CrossRef]
- Gauri, S.S.; Mandal, S.M.; Dey, S.; Pati, B.R. Biotransformation of p-coumaric acid and 2,4-dichlorophenoxy acetic acid by Azotobacter sp. strain SSB81. Bioresour. Technol. 2012, 126, 350–353. [Google Scholar] [CrossRef]
- Zhang, B.; Weston, L.A.; Li, M.; Zhu, X.; Weston, P.A.; Feng, F.; Zhang, B.; Zhang, L.; Gu, L.; Zhang, Z. Rehmannia glutinosa replant issues: Root exudate-rhizobiome interactions clearly influence replant success. Front. Microbiol. 2020, 11, 1413. [Google Scholar] [CrossRef]
- Iqbal, N.; Khan, N.A.; Ferrante, A.; Trivellini, A.; Francini, A.; Khan, M. Ethylene role in plant growth, development and senescence: Interaction with other phytohormones. Front. Plant Sci. 2017, 8, 475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadeghi, H.; Ghanaatiyan, K. Probing the responses of four chicory ecotypes by ethylene accumulation and growth characteristics under drought stress. Ital. J. Agron. 2017, 12, 773. [Google Scholar] [CrossRef]
- Brunetti, C.; Saleem, A.R.; Della, R.G.; Emiliani, G.; De Carlo, A.; Balestrini, R.; Khalid, A.; Mahmood, T.; Centritto, M. Effects of plant growth-promoting rhizobacteria strains producing ACC deaminase on photosynthesis, isoprene emission, ethylene formation and growth of Mucuna pruriens (L.) DC. in response to water deficit. J. Biotechnol. 2021, 331, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Glick, B.R.; Nascimento, F.X. Pseudomonas 1-Aminocyclopropane-1-carboxylate (ACC) deaminase and its role in beneficial plant-microbe interactions. Microorganisms 2021, 9, 2467. [Google Scholar] [CrossRef] [PubMed]
- Ravanbakhsh, M.; Sasidharan, R.; Voesenek, L.A.; Kowalchuk, G.A.; Jousset, A. ACC deaminase-producing rhizosphere bacteria modulate plant responses to flooding. J. Ecol. 2017, 105, 979–986. [Google Scholar] [CrossRef]
- Orhan, F. Potential of halophilic/halotolerant bacteria in enhancing plant growth under salt stress. Curr. Microbiol. 2021, 78, 3708–3719. [Google Scholar] [CrossRef]
- Li, Y.; Kong, D.; Fu, Y.; Sussman, M.R.; Wu, H. The effect of developmental and environmental factors on secondary metabolites in medicinal plants. Plant Physiol. Biochem. 2020, 148, 80–89. [Google Scholar] [CrossRef]
- Liu, H.; Gu, H.; Ye, C.; Guo, C.; Zhu, Y.; Huang, H.; Liu, Y.; He, X.; Yang, M.; Zhu, S. Planting density affects Panax notoginseng growth and ginsenoside accumulation by balancing primary and secondary metabolism. Front. Plant Sci. 2021, 12, 628294. [Google Scholar] [CrossRef]
- Abdul, M.N.; Kumar, I.S.; Nadarajah, K. Elicitor and receptor molecules: Orchestrators of plant defense and immunity. Int. J. Mol. Sci. 2020, 21, 963. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Liang, W.; Lu, J.; Yao, L.; Wang, J.; Gao, W. Penicillium sp. YJM-2013 induces ginsenosides biosynthesis in Panax ginseng adventitious roots by inducing plant resistance responses. Chin. Herb. Med. 2020, 12, 257–264. [Google Scholar] [CrossRef]
- Chamkhi, I.; Benali, T.; Aanniz, T.; El Menyiy, N.; Guaouguaou, F.; El Omari, N.; El-Shazly, M.; Zengin, G.; Bouyahya, A. Plant-microbial interaction: The mechanism and the application of microbial elicitor induced secondary metabolites biosynthesis in medicinal plants. Plant Physiol. Biochem. 2021, 167, 269–295. [Google Scholar] [CrossRef] [PubMed]
- Ho, T.T.; Lee, J.D.; Jeong, C.S.; Paek, K.Y.; Park, S.Y. Improvement of biosynthesis and accumulation of bioactive compounds by elicitation in adventitious root cultures of Polygonum multiflorum. Appl. Microbiol. Biotechnol. 2018, 102, 199–209. [Google Scholar] [CrossRef]
- Zhou, X.; Wu, Y.; Wang, X.; Liu, B.; Xu, H. Salidroside production by hairy roots of Rhodiola sachalinensis obtained after transformation with Agrobacterium rhizogenes. Biol. Pharm. Bull. 2007, 30, 439–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, Y.; Guo, L.-P.; Chen, B.; Hao, Z.; Wang, J.-Y.; Huang, L.-Q.; Yang, G.; Cui, X.-M.; Yang, L.; Wu, Z.-X.; et al. Arbuscular mycorrhizal symbiosis and active ingredients of medicinal plants: Current research status and prospectives. Mycorrhiza 2013, 23, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, T.; Shukla, P. Bioinoculants for bioremediation applications and disease resistance: Innovative perspectives. Indian J. Microbiol. 2019, 59, 129–136. [Google Scholar] [CrossRef]
- Enebe, M.C.; Babalola, O.O. The impact of microbes in the orchestration of plants’ resistance to biotic stress: A disease management approach. Appl. Microbiol. Biotechnol. 2019, 103, 9–25. [Google Scholar] [CrossRef] [Green Version]
- Gao, M.; Xiong, C.; Gao, C.; Tsui, C.; Wang, M.-M.; Zhou, X.; Zhang, A.-M.; Cai, L. Disease-induced changes in plant microbiome assembly and functional adaptation. Microbiome 2021, 9, 187. [Google Scholar] [CrossRef]
- Pangesti, N.; Pineda, A.; Dicke, M.; van Loon, J.J.A.; Leiss, K.; Leiss, K. Variation in plant-mediated interactions between rhizobacteria and caterpillars: Potential role of soil composition. Plant Biol. 2015, 17, 474–483. [Google Scholar] [CrossRef]
- Beneduzi, A.; Ambrosini, A.; Passaglia, L.M. Plant growth-promoting rhizobacteria (PGPR): Their potential as antagonists and biocontrol agents. Genet. Mol. Biol. 2012, 35, 1044–1051. [Google Scholar] [CrossRef] [Green Version]
- Kloepper, J.W.; Ryu, C.; Zhang, S. Induced systemic resistance and promotion of plant growth by Bacillus spp. Phytopathology 2004, 94, 1259–1266. [Google Scholar] [CrossRef] [Green Version]
- Buttner, H.; Niehs, S.P.; Vandelannoote, K.; Cseresnyes, Z.; Dose, B.; Richter, I.; Gerst, R.; Figge, M.T.; Stinear, T.P.; Pidot, S.J.; et al. Bacterial endosymbionts protect beneficial soil fungus from nematode attack. Proc. Natl. Acad. Sci. USA 2021, 118, e2110669118. [Google Scholar] [CrossRef]
- Lamdan, N.L.; Shalaby, S.; Ziv, T.; Kenerley, C.M.; Horwitz, B.A. Secretome of Trichoderma interacting with maize roots: Role in induced systemic resistance. Mol. Cell. Proteomics 2015, 14, 1054–1063. [Google Scholar] [CrossRef] [PubMed]
- Su, P.; Tan, X.; Li, C.; Zhang, D.; Cheng, J.; Zhang, S.; Zhou, X.; Yan, Q.; Peng, J.; Zhang, Z.; et al. Photosynthetic bacterium Rhodopseudomonas palustris GJ-22 induces systemic resistance against viruses. Microb. Biotechnol. 2017, 10, 612–624. [Google Scholar] [CrossRef]
- Patel, S.; Thaker, V.; Kuvad, R.; Saraf, M. Role of lipopolysaccaride extracted from Alcaligenes faecalis as elicitor for the induction of plant defense against fusarium wilt. J. Plant Pathol. 2019, 102, 351–357. [Google Scholar] [CrossRef]
- Yin, C.; Casa Vargas, J.M.; Schlatter, D.C.; Hagerty, C.H.; Hulbert, S.H.; Paulitz, T.C. Rhizosphere community selection reveals bacteria associated with reduced root disease. Microbiome 2021, 9, 86. [Google Scholar] [CrossRef]
- Bozsoki, Z.; Gysel, K.; Hansen, S.B.; Lironi, D.; Kronauer, C.; Feng, F.; de Jong, N.; Vinther, M.; Kamble, M.; Thygesen, M.B.; et al. Ligand-recognizing motifs in plant LysM receptors are major determinants of specificity. Science 2020, 369, 663–670. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Hong, S.; Xiong, W.; Raza, W.; Shen, Z.; Wang, B.; Li, R.; Ruan, Y.; Shen, Q.; Dini-Andreote, F. Development of fungal-mediated soil suppressiveness against Fusarium wilt disease via plant residue manipulation. Microbiome 2021, 9, 200. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.R.; Jeon, C.W.; Cho, G.; Thomashow, L.S.; Weller, D.M.; Paik, M.J.; Lee, Y.B.; Kwak, Y.-S. Glutamic acid reshapes the plant microbiota to protect plants against pathogens. Microbiome 2021, 9, 244. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Xun, W.; Wang, X.; Tian, S.; Zhang, Y.; Li, D.; Zhou, Y.; Qin, Y.; Zhang, B.; Zhao, G.; et al. Root-secreted bitter triterpene modulates the rhizosphere microbiota to improve plant fitness. Nat. Plants 2022, 8, 887–896. [Google Scholar] [CrossRef]
- Santoyo, G. How plants recruit their microbiome? New insights into beneficial interactions. J. Adv. Res. 2021, 40, 45–58. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Wu, H.; Jiao, Y.; Zhang, Z.; Rensing, C.; Lin, W. The combination of biochar and PGPBs stimulates the differentiation in rhizosphere soil microbiome and metabolites to suppress soil-borne pathogens under consecutive monoculture regimes. GCB Bioenergy 2022, 14, 84–103. [Google Scholar] [CrossRef]
- Zhang, Y.; Gao, X.; Shen, Z.; Zhu, C.; Jiao, Z.; Li, R.; Shen, Q. Pre-colonization of PGPR triggers rhizosphere microbiota succession associated with crop yield enhancement. Plant Soil 2019, 439, 553–567. [Google Scholar] [CrossRef]
- Fira, D.; Dimkić, I.; Berić, T.; Lozo, J.; Stanković, S. Biological control of plant pathogens by Bacillus species. J. Biotechnol. 2018, 285, 44–55. [Google Scholar] [CrossRef] [PubMed]
- Jiang, G.; Zhang, Y.; Gan, G.; Li, W.; Wan, W.; Jiang, Y.; Yang, T.; Zhang, Y.; Xu, Y.; Wang, Y.; et al. Exploring rhizo-microbiome transplants as a tool for protective plant-microbiome manipulation. ISME Commun. 2022, 2, 10. [Google Scholar] [CrossRef]
- Wang, M.; Li, S.; Chen, S.; Meng, N.; Li, X.; Zheng, H.; Zhao, C.; Wang, D. Manipulation of the rhizosphere bacterial community by biofertilizers is associated with mitigation of cadmium phytotoxicity. Sci. Total. Environ. 2019, 649, 413–421. [Google Scholar] [CrossRef]
- Shabaan, M.; Asghar, H.N.; Akhtar, M.J.; Ali, Q.; Ejaz, M. Role of plant growth promoting rhizobacteria in the alleviation of lead toxicity to Pisum sativum L. Int. J. Phytoremediat. 2021, 23, 837–845. [Google Scholar] [CrossRef]
- de Fatima, P.D.; Barbosa, M.V.; Dos, S.J.; Pinto, F.A.; Siqueira, J.O.; Carneiro, M. Arbuscular mycorrhizal fungi favor the initial growth of Acacia mangium, Sorghum bicolor, and Urochloa brizantha in soil contaminated with Zn, Cu, Pb, and Cd. Bull. Environ. Contam. Toxicol. 2018, 101, 386–391. [Google Scholar] [CrossRef]
- Oliveira, K.O.; Silva, A.R.M.; da Silva, B.F.; Milagre, H.M.S.; Milagre, C.D.F. Insights into the microbial degradation pathways of the ioxynil octanoate herbicide. Biocatal. Agric. Biotechnol. 2018, 13, 258–264. [Google Scholar] [CrossRef] [Green Version]
- Pan, X.; Wang, S.; Shi, N.; Fang, H.; Yu, Y. Biodegradation and detoxification of chlorimuron-ethyl by Enterobacter ludwigii sp. CE-1. Ecotoxicol. Environ. Saf. 2018, 150, 34–39. [Google Scholar] [CrossRef]
- Gupta, M.; Mathur, S.; Sharma, T.K.; Rana, M.; Gairola, A.; Navani, N.K.; Pathania, R. A study on metabolic prowess of Pseudomonas sp. RPT 52 to degrade imidacloprid, endosulfan and coragen. J. Hazard. Mater. 2016, 301, 250–258. [Google Scholar] [CrossRef]
- Li, H.; Qiu, Y.; Yao, T.; Ma, Y.; Zhang, H.; Yang, X.; Li, C. Evaluation of seven chemical pesticides by mixed microbial culture (PCS-1): Degradation ability, microbial community, and Medicago sativa phytotoxicity. J. Hazard. Mater. 2020, 389, 121834. [Google Scholar] [CrossRef]
- Abdelhameed, R.E.; Metwally, R.A. Alleviation of cadmium stress by arbuscular mycorrhizal symbiosis. Int. J. Phytoremediat. 2019, 21, 663–671. [Google Scholar] [CrossRef]
- Yue, Z.; Chen, Y.; Chen, C.; Ma, K.; Tian, E.; Wang, Y.; Liu, H.; Sun, Z. Endophytic Bacillus altitudinis WR10 alleviates Cu toxicity in wheat by augmenting reactive oxygen species scavenging and phenylpropanoid biosynthesis. J. Hazard. Mater. 2021, 405, 124272. [Google Scholar] [CrossRef]
- Chen, B.; Xiao, X.; Zhu, Y.G.; Smith, F.A.; Xie, Z.M.; Smith, S.E. The arbuscular mycorrhizal fungus Glomus mosseae gives contradictory effects on phosphorus and arsenic acquisition by Medicago sativa Linn. Sci. Total Environ. 2007, 379, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Chen, Z.; Kowalchuk, G.A.; Xu, Z.; Fu, X.; Kuramae, E.E. Succession of the resident soil microbial community in response to periodic inoculations. Appl. Environ. Microbiol. 2021, 87, e00046-21. [Google Scholar] [CrossRef] [PubMed]
- Morya, R.; Salvachua, D.; Thakur, I.S. Burkholderia: An untapped but promising bacterial genus for the conversion of aromatic compounds. Trends Biotechnol. 2020, 38, 963–975. [Google Scholar] [CrossRef]
- Hayat, K.; Menhas, S.; Bundschuh, J.; Zhou, P.; Niazi, N.K.; Amna; Hussain, A.; Hayat, S.; Ali, H.; Wang, J.; et al. Plant growth promotion and enhanced uptake of Cd by combinatorial application of Bacillus pumilus and EDTA on Zea mays L. Int. J. Phytoremediat. 2020, 22, 1372–1384. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhang, N.; Zhang, Z.; Luo, J.; Shen, B.; Zhang, R.; Shen, Q. Antagonist Bacillus subtilis HJ5 controls Verticillium wilt of cotton by root colonization and biofilm formation. Biol. Fertil. Soils 2012, 49, 295–303. [Google Scholar] [CrossRef]
- Sun, X.; Xu, Z.; Xie, J.; Hesselberg-Thomsen, V.; Tan, T.; Zheng, D.; Strube, M.L.; Dragoš, A.; Shen, Q.; Zhang, R.; et al. Bacillus velezensis stimulates resident rhizosphere Pseudomonas stutzeri for plant health through metabolic interactions. ISME J. 2022, 16, 774–787. [Google Scholar] [CrossRef]
- Dan, A.; Zhang, N.; Qiu, R.; Li, C.; Wang, S.; Ni, Z. Accelerated biodegradation of p-tert-butylphenol in the Phragmites australis rhizosphere by phenolic root exudates. Environ. Exp. Bot. 2020, 169, 103891. [Google Scholar] [CrossRef]
- Xia, L.; Miao, Y.; Cao, A.; Liu, Y.; Liu, Z.; Sun, X.; Xue, Y.; Xu, Z.; Xun, W.; Shen, Q.; et al. Biosynthetic gene cluster profiling predicts the positive association between antagonism and phylogeny in Bacillus. Nat. Commun. 2022, 13, 1023. [Google Scholar] [CrossRef] [PubMed]




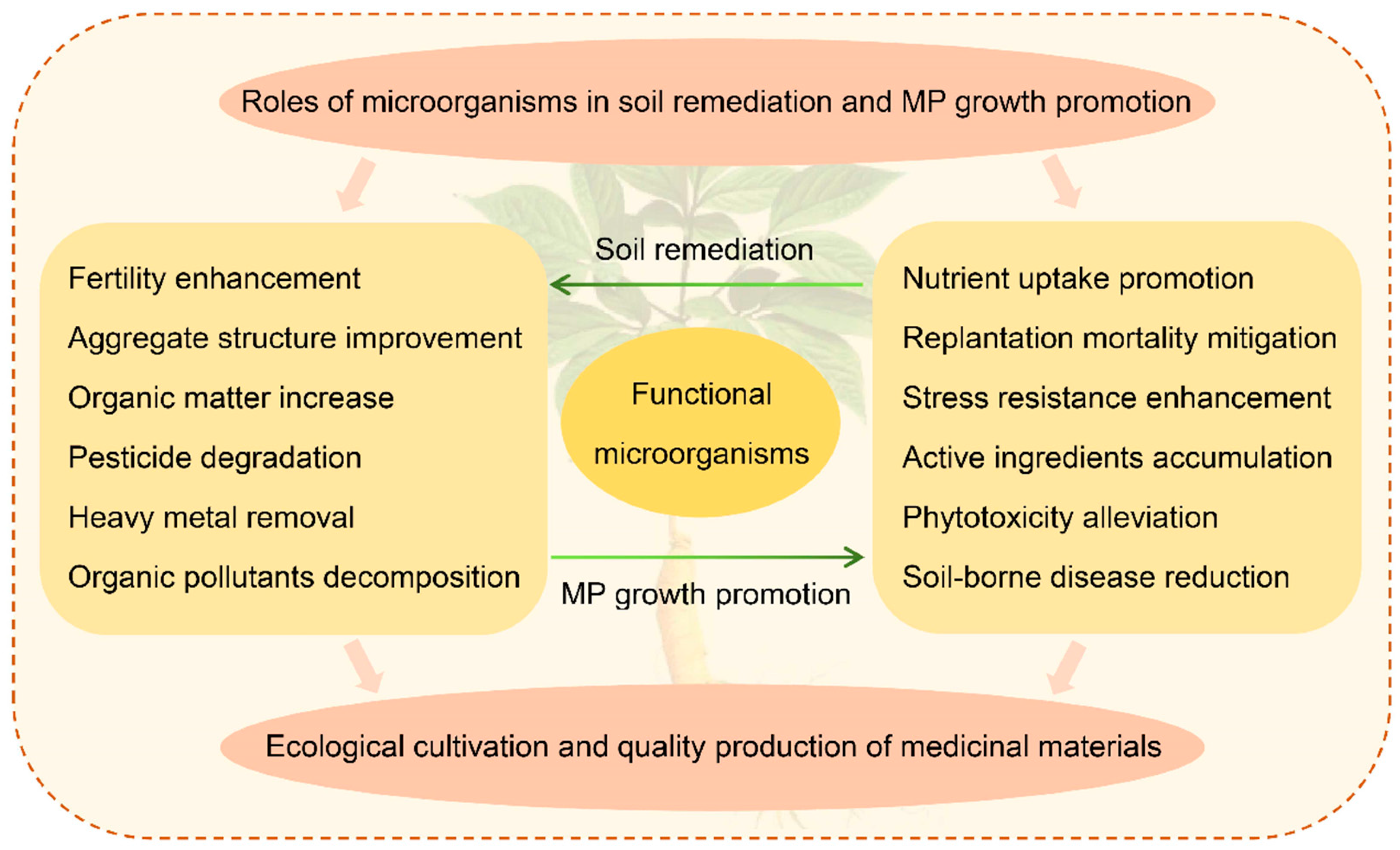{kind=link}
{kind=link}
{kind=link}
| Soil Issues | Strains/Microbes | Contributions | Reference |
|---|---|---|---|
| Nutrient deficiency and imbalance | Pseudomonas libanensis EU-LWNA-33 | Increasing P solubilization | [23] |
| Herbaspirillum sp. H18 and ZA15, Burkholderia vietaminensis AR114 | Increasing P solubilization and N use efficiency | [133] | |
| Enterobacter cloacae RCA25, Klebsiella variicola RCA26, Herbaspirillum seropedicae z67, Sinorhizobium fredii NGR234 | IAA production, improving N fixation | [134] | |
| Paenibacillus spp. | Catalyzing N2-fixing | [135] | |
| Pseudomonas sp. S10-3 | Increasing K solubilization | [136] | |
| Burkholderia spp. | Increasing K solubilization | [137] | |
| Pantoea agglomerans, Rahnella aquatilis, Pseudomonas orientalis | Increasing K solubilization, IAA production | [138] | |
| Providencia rettgeri TPM23 | Increasing available N, P and K contents | [139] | |
| Enterobacter cloacae HG-1 | Enhancing N fixation, producing iron carriers, ACC deaminase and plant hormones | [140] | |
| Rhizobium strains NSFBR-12, NSFBR-15 | Enhancing N fixation | [141] | |
| Azospirillum brasilense | Improving fertilizer-N recovery | [142] | |
| Pesticide residues | Microbacterium sp. D-2 | Dicofol degradation | [143] |
| Acinetobacter baylyi GFJ2 | Chloroanilines degradation | [144] | |
| Pseudomonas sp. | DDT degradation | [145] | |
| Pseudomonas putida, Acinetobacter rhizosphaerae | Hydrolysis of organophosphate and carbamate pesticides | [146] | |
| Chryseobacterium sp. Y16C | Glyphosate degradation | [147] | |
| Arthrobacter sp. HB-5 | Atrazine degradation | [148] | |
| Trichoderma atroviride T23 | Dichlorvos degradation | [149] | |
| Alcaligenes faecalis NBRI OSS2-5 | Chlorpyrifos degradation | [150] | |
| Bacillus pumilus W1 | Organophosphate degradation | [151] | |
| Pseudomonas nitroreducens AR-3 | Chlorpyrifos degradation | [152] | |
| Pseudomonas putida ATCC 49451 | Chlorophenols degradation | [153] | |
| Bacillus aryabhattai VITNNDJ5 | Monocrotophos degradation | [154] | |
| Sphingomonas sp. AK1 | Isoproturon degradation | [155] | |
| Bacillus spp. | Diuron degradation | [156] | |
| Catellibacterium sp. CC-5 | Cypermethrin degradation | [157] | |
| Xanthomonas axonopodis, Aspergillus niger | Alachlor degradation | [158] | |
| Bacillus cereus Y1 | Deltamethrin degradation | [159] | |
| Excessive heavy metals | Bacillus thuringiensis WS3 | As removal | [160] |
| Pseudomonas putida UW4 | PAH and Pb removal | [161] | |
| Kocuria rhizophila | Promoting phytoremediation of Cd, Cr, Cu and Ni-contaminated soil | [162] | |
| Bacillus cereus HM5, Bacillus thuringiensis HM7 | Improving Mn phytoremediation | [163] | |
| Pseudomonas lurida EOO26 | Improving Cu phytoremediation | [164] | |
| Bacillus subtilus MAI3 | Chromium (VI) reduction and Cr (III) immobilization | [165] | |
| Trichoderma harzianum, Bacillus subtilis | Cd bioavailability reduction | [166] | |
| Pantoea conspicua MT5, Aspergillus niger CRS3 | Reduction and detoxification of Cr (Ⅵ) | [167] | |
| Organic pollutants | Hyphomicrobium sp. GHH | 17α-ethinyestradiol removal | [168] |
| Klebsiella pneumonia PL1 | Pyrene and benzo(a)pyrene removal | [169] | |
| Pseudomonas mendocina, Brevundimonas olei, Serratia marcescens | Creosote PAH degradation | [170] | |
| Psuedomonas sp. USTB-RU | Phenanthrene degradation | [171] | |
| Sphingobium yanoikuyae B1 | PAH degradation | [172] | |
| Pseudomonas sp. SDR4, Mortierella alpina JDR7 | PAH degradation | [173] | |
| Mycobacterium spp. | Pyrene degradation | [174] | |
| Ralstonia sp. U2 | Naphthalene degradation | [175] | |
| Pseudomonas aeruginosa PSA5, Rhodococcus sp. NJ2 | Benzo(a)pyrene degradation | [176] | |
| Pseudomonas oleovorans DT4 | Tetrahydrofuran, benzene, toluene, ethylbenzene and xylene degradation | [177] | |
| Raoultella sp. XY-1, Pandoraea sp. XY-2 | Tetracycline degradation | [178] |
| Medicinal Plants | Strains/Microbes | Role | Reference |
|---|---|---|---|
| Panax ginseng | Paenibacillus polymyxa | Simultaneous degradation of fluazinam, BHC, PCNB, chlorpyrifos and DDT in roots, stems and leaves | [26] |
| Bacillus subtilis 50-1 | Replanting mortality alleviation | [229] | |
| Bacillus amyloliquefaciens SW-34 | Grey mold suppression | [230] | |
| Bacillus amyloliquefaciens HK34 | Foliar blight and root rot control | [231] | |
| Pseudomonas aeruginosa D4 and Bacillus Stratosphericus FW3 | Root rot fungal pathogen control | [232] | |
| Rhizobium panacihumi DCY116T | Al resistance enhancement | [233] | |
| Sphingobacterium sp. PG-1 | Toxic diisobutyl phthalate degradation, replanting issue alleviation, growth promotion | [234] | |
| P. quinquefolius | Rhizoglomus irregulare, Funneliformis mosseae and F. caledonium | Soil-borne pathogen control, nutrient acquisition improvement, continuous cropping obstacle mitigation | [235] |
| Glycyrrhiza glabra | Funneliformis mosseae | Salt stress alleviation, P and K-concentration increase, glycyrrhizin accumulation | [236] |
| G. uralensis | Glomus mosseae and G. veriforme | Growth promotion, P-acquisition improvement, glycyrrhizin accumulation | [237] |
| Rhizophagus irregularis | Compensation for the loss of indigenous microbial communities, growth and secondary metabolism promotion | [238] | |
| Acrocalymma vagum | Biomass increase under drought stress, glycyrrhizic acid and glycyrrhizin accumulation | [239] | |
| Salvia miltiorrhiza | Bacillus cereus | Hairy root growth promotion, tanshinone production increase | [240] |
| Bacillus amyloliquefaciens, B. licheniformis and Actinomyces bovis | Cd-uptake reduction, total tanshinones accumulation | [241] | |
| S. officinalis | Glomus intraradices | Essential oil yield and quality improvement | [242] |
| Artemisia annua | Colletotrichum sp. | Artemisinin production stimulation | [243] |
| Rizophagus irregularis | Artemisinin content and essential oil yield increase | [244] | |
| Glomus mosseae and Bacillus subtilis Daz26 | Growth, biomass yield, and artemisinin content enhancement | [245] | |
| Sophora flavescens | Actinobacteria and Chloroflexi | Matrine and oxymatrine accumulation | [246] |
| Bradyrhizobium arachidis CCBAU 051107T | Oxymatrine and matrine content enhancement | [247] | |
| Pistacia vera | Staphylococcus sciuri, Zobellella denitrificans and Arthrobacter endophyticus | Yield increase, photosynthesis promotion, water absorption promotion, performance improvement under salinity and drought stresses | [248] |
| Bacopa monnieri | Pseudomonas plecoglossicida KM233646, Acinetobacter calcoaceticus KM233647, Bacillus flexus KM233648 and B. safensis KM233652 | Growth promotion, bacoside A yield increase, saline soil reclamation | [249] |
| Echinacea purpurea | Pseudomonas fluorescens, AMF | Nutrient concentration increase, water absorption promotion, growth improvement, drought stress alleviation | [250] |
| Trigonella foenum-graecum | Bacillus subtilis LDR2 | Nodulation and AMF colonization enhancement, nutrient uptake improvement, growth promotion, drought-stress resistance enhancement | [251] |
| Azotobacter chroococcum and Peudomoance fluorescence | Yield and trigonelline production enhancement under deficit irrigation | [252] | |
| Foeniculum vulgare | Bacillus subtilis PSB-1 and PSB-36 | Seed yield and essential oil content increase, P-availability enhancement | [253] |
| Rheum palmatum | Bacillus amyloliquefaciens EZ99 | Increase of root fresh weight, active components content and K-availability | [254] |
| Naravelia zeylanica | Achromobacter xylosoxidans AUM54 | Growth promotion, survivability and stress tolerance increase | [255] |
| Astragalus mongholicus | Stenotrophomonas, Phyllobacterium and Inquilinus | Bioactive ingredients accumulation | [256] |
| Polygonum cuspidatum | Bacteroides, Acinetobacter, Erysipelatoclostridium and Achromobacter | Resveratrol accumulation | [257] |
| Chrysanthemum morifolium | Funneliformis mosseae and Diversispora versiformis | N-uptake enhancement, growth promotion under salt stress | [258] |
| Papaver somniferum | Pseudomonas putida | Growth and yield improvement, downy mildew tolerance enhancement | [259] |
| Zingiber officinale | Bacillus, Pseudomonas, Arthrobacter and Serratia | Al-toxicity and bacterial wilt alleviation | [260] |
| Andrographis paniculata | Bacillus sp. CIMAP-A7 | Atrazine-induced toxicity amelioration, oxidative stress amelioration | [261] |
| Azotobacter chrococcum, Bacillus megaterium, Pseudomonas monteilii, Glomus intraradices | Soil dehydrogenase, alkaline and acidic phosphatase activity improvement, growth promotion, pharmacological quality enhancement | [262] | |
| Commiphora leptophloeos | Gigaspora albida and Claroideoglomus etunicatum | Total foliar phenols and tannins increase | [263] |
| Ocimum basilicum | Glomus caledonium BEG 162 and G. mosseae NBR 1–2 | Rosmarinic and caffeic acids production enhancement | [264] |
| Atractylodes lancea | Pseudomonas fluorescens ALEB7B | Medicinal sesquiterpenoid accumulation, photosynthesis enhancement | [265] |
| Hypericum perforatum | Stenotrophomonas maltophilia N5.18 | Hypericin and pseudohypericin increase | [266] |
| Rhizophagus intraradices, Funneliformis constrictum, F. geosporum and F. mosseae | Photosynthetic activity stimulation, secondary metabolites production, hypericin and pseudohypericin concentration enhancement | [267] | |
| Hyoscyamus niger | Pseudomonas putida and P. fluorescens | Growth promotion under water deficit stress, alkaloid production increase | [268] |
| Crocus sativus | Rhizophagus intraradicse, Funneliformis mosseae, Rhizophagus irregutaris and Glomus caledonium | Photosynthesis promotion, increase of flower number, leaf dry weight and area and yield | [269] |
| Mentha arvensis | Exiguobacterium oxidotoleran, and Glomus fasciculatum | Growth promotion in salt-stressed soil, essential oil yield increase | [270] |
| Trichoderma harzianum and Brevibacterium halotolerans | Increase in plant growth, oil content, leaf-stem ratio, photosynthetic pigments and nutrient uptake | [271] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, G.; Ren, Y.; Bai, X.; Su, Y.; Han, J. Contributions of Beneficial Microorganisms in Soil Remediation and Quality Improvement of Medicinal Plants. Plants 2022, 11, 3200. https://doi.org/10.3390/plants11233200
Wang G, Ren Y, Bai X, Su Y, Han J. Contributions of Beneficial Microorganisms in Soil Remediation and Quality Improvement of Medicinal Plants. Plants. 2022; 11(23):3200. https://doi.org/10.3390/plants11233200
Chicago/Turabian StyleWang, Gang, Ying Ren, Xuanjiao Bai, Yuying Su, and Jianping Han. 2022. "Contributions of Beneficial Microorganisms in Soil Remediation and Quality Improvement of Medicinal Plants" Plants 11, no. 23: 3200. https://doi.org/10.3390/plants11233200