
Potassium Application Enhanced Plant Growth, Mineral Composition, Proximate and Phytochemical Content in Trachyandra divaricata Kunth (Sandkool)


Abstract
:1. Introduction
2. Results
2.1. Effect of Potassium and Pruning Levels on Plant Growth
2.1.1. Height and Number of Flower Buds
2.1.2. Fresh and Dry Weight of Flower Buds
2.1.3. Fresh and Dry Weight of Shoots
2.1.4. Fresh and Dry Weight of Roots
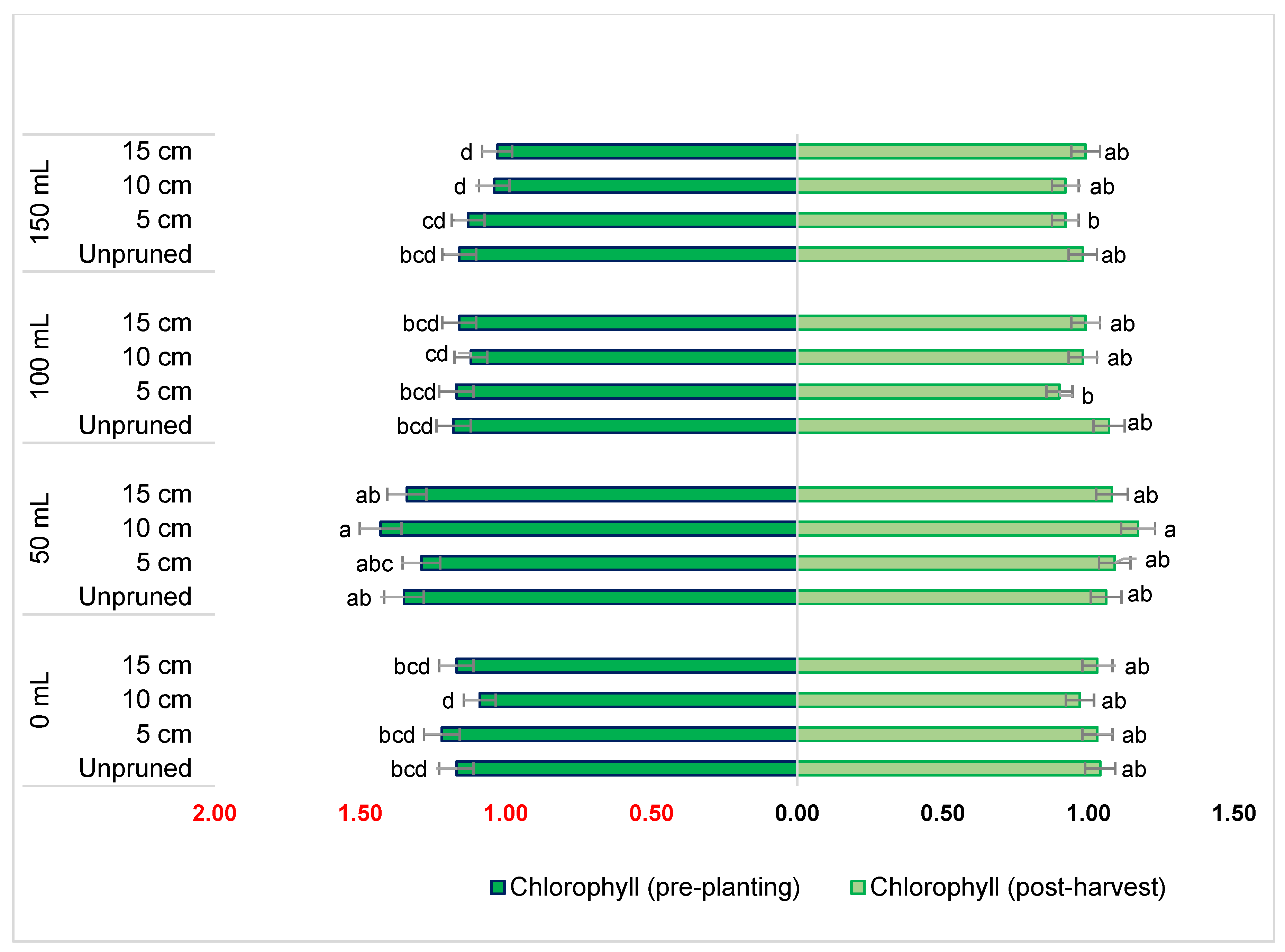
2.2. Effect of Potassium Dosage and Pruning Levels on Chlorophyll Content
2.3. Effect of Potassium and Pruning on Mineral Composition of Flower Buds of T. divaricata
2.3.1. Macronutrients
2.3.2. Micronutrients
2.4. Effect of Potassium and Pruning on Proximate Composition of Flower Buds
2.5. Effect of Potassium and Pruning Levels on Phytochemicals and Antioxidant Activity of Flower Buds
2.5.1. Total Polyphenols
2.5.2. Total Flavonol
2.5.3. FRAP Antioxidant Content
2.5.4. DPPH Antioxidant Content
2.5.5. TEAC/ABTS Activity
3. Materials and Methods
3.1. Experimental Location
3.2. Plant Preparation and Experimental Design
3.3. Treatments
3.4. Determination of Plant Growth
3.4.1. Shoot Length and the Number of Flower Buds
3.4.2. Plant Weight
3.5. Chlorophyll Content
3.6. Mineral Analysis
3.7. Proximate Analysis
3.7.1. Moisture Content
3.7.2. Crude Fibre Content
3.7.3. Crude Fat Content
3.7.4. Ash Content
3.7.5. Crude Protein
3.7.6. Neutral Detergent Fibre (NDF)
3.7.7. Non-Fibre Carbohydrate (NFC)
3.7.8. Energy Content
3.8. Phytochemicals and Antioxidant Assays
3.8.1. Sample Preparation
3.8.2. Polyphenol Assay
3.8.3. Estimation of Flavonol Content
3.8.4. Ferric Reducing Antioxidant Power (FRAP) Assay
3.8.5. DPPH Free Radical Scavenging Activity
3.8.6. ABTS Free Radical Scavenging Activity
3.9. Statistical Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Anju, T.; Prabhakar, P.; Sreedharan, S.; Kumar, A. Nutritional, Antioxidant and Dietary Potential of Some Traditional Leafy Vegetables Used in Ethnic Culinary Preparations. Food Control 2022, 141, 109161. [Google Scholar] [CrossRef]
- Sultanbawa, Y.; Sivakumar, D. Enhanced Nutritional and Phytochemical Profiles of Selected Underutilized Fruits, Vegetables, and Legumes. Curr. Opin. Food Sci. 2022, 46, 100853. [Google Scholar] [CrossRef]
- Leal Filho, W.; Barbir, J.; Gwenzi, J.; Ayal, D.; Simpson, N.P.; Adeleke, L.; Tilahun, B.; Chirisa, I.; Gbedemah, S.F.; Nzengya, D.M.; et al. The Role of Indigenous Knowledge in Climate Change Adaptation in Africa. Environ. Sci. Policy 2022, 136, 250–260. [Google Scholar] [CrossRef]
- Maroyi, A. Traditional Uses of Wild and Tended Plants in Maintaining Ecosystem Services in Agricultural Landscapes of the Eastern Cape Province in South Africa. J. Ethnobiol. Ethnomed. 2022, 18, 17. [Google Scholar] [CrossRef] [PubMed]
- Ntuli, N.R. Nutrient Content of Scarcely Known Wild Leafy Vegetables from Northern KwaZulu-Natal, South Africa. S. Afr. J. Bot. 2019, 127, 19–24. [Google Scholar] [CrossRef]
- Zulu, S.S.; Ngidi, M.; Ojo, T.; Hlatshwayo, S.I. Determinants of Consumers’ Acceptance of Indigenous Leafy Vegetables in Limpopo and Mpumalanga Provinces of South Africa. J. Ethn. Foods 2022, 9, 13. [Google Scholar] [CrossRef]
- Ngxabi, S.; Jimoh, M.O.; Kambizi, L.; Laubscher, C.P. Growth Characteristics, Phytochemical Contents, and Antioxidant Capacity of Trachyandra ciliata (L.f) Kunth Grown in Hydroponics under Varying Degrees of Salinity. Horticulturae 2021, 7, 244. [Google Scholar] [CrossRef]
- Heyligers, P.C. Dispersal of the Exotic Coastal Dune Plants Gladiolus Gueinzii and Trachyandra Divaricata in Australia. Cunninghamia 1999, 6, 315–330. [Google Scholar]
- Van Wyk, B.E. The Potential of South African Plants in the Development of New Food and Beverage Products. S. Afr. J. Bot. 2011, 77, 857–868. [Google Scholar] [CrossRef] [Green Version]
- De Vynck, J.C.; van Wyk, B.E.; Cowling, R.M. Indigenous Edible Plant Use by Contemporary Khoe-San Descendants of South Africa’s Cape South Coast. S. Afr. J. Bot. 2016, 102, 60–69. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J. Potassium Control of Plant Functions: Ecological and Agricultural Implications. Plants 2021, 10, 419. [Google Scholar] [CrossRef]
- Marques, D.J.; Bianchini, H.C.; da Silva Lobato, A.K.; da Silva, W.F. Potassium Fertilization in the Production of Vegetables and Fruits. In Potassium-Improvement of Quality in Fruits and Vegetables through Hydroponic Nutrient Management; InTech Open: London, UK, 2018. [Google Scholar] [CrossRef] [Green Version]
- Gelaye, Y.; Alemayehu, M.; Ademe, D. Potato (Solanum tuberosum L.) Growth and Quality as Influenced by Inorganic Fertilizer Rates in Northwestern Ethiopia. Int. J. Agron. 2022, 2022, 9476021. [Google Scholar] [CrossRef]
- Wang, Y.T.; Liu, R.L.; Huang, S.W.; Jin, J.Y. Effects of potassium application on flavor compounds of cherry tomato fruits. J. Plant Nutr. 2009, 32, 1451–1468. [Google Scholar] [CrossRef]
- Pandey, G.K.; Mahiwal, S. Potassium in Abiotic Stress. In Role of Potassium in Plants; Springer: Cham, Switzerland, 2020; pp. 45–49. [Google Scholar]
- Rani, P.; Saini, I.; Singh, N.; Kaushik, P.; Wijaya, L.; Al-Barty, A.; Darwish, H.; Noureldeen, A. Effect of Potassium Fertilizer on the Growth, Physiological Parameters, and Water Status of Brassica Juncea Cultivars under Different Irrigation Regimes. PLoS ONE 2021, 16, e0257023. [Google Scholar] [CrossRef] [PubMed]
- Mawarni, L.; Siahaan, M.D.A. Effect of Chicken Manure and Pruning on Kyuri Cucumber Plant. IOP Conf. Ser. Earth Environ. Sci. 2022, 977, 012043. [Google Scholar] [CrossRef]
- Nabi, J.; Narayan, I.S.; Malik, I.A.; Mir, I.S.; Wani, I.J.; Khan, I.F.; Bhat, I.T.; Shah, I.L.; Nisar, I.F.; Hussain, I.K.; et al. ZA Bhat Division of Floriculture and Landscaping Influence of Fertigation and Pruning Levels on Soil Status and Mineral Content of Parthenocarpic Cucumber under Polyhouse Conditions. Pharma Innov. J. 2022, SP-11, 605–609. [Google Scholar]
- Hartatik, S.; Hudah, M.; Soeparjono, S. Suharto Shoot Pruning and Potassium Application Effect on Cucumber (Cucumis sativus L.) Seeds Production and Quality. IOP Conf. Ser. Earth Environ. Sci. 2021, 709, 012067. [Google Scholar] [CrossRef]
- Hasanah, Y.; Rahmawati, N.; Nasution, K.H. Growth and Production of Cilembu Sweet Potatoes (Ipomoea batatas L.) Varieties in the Highlands with Potassium Fertilizer and Pruning Treatments. IOP Conf. Ser. Earth Environ. Sci. 2021, 782, 042027. [Google Scholar] [CrossRef]
- Viljoen, C.C.; Jimoh, M.O.; Laubscher, C.P. Studies of Vegetative Growth, Inflorescence Development and Eco-Dormancy Formation of Abscission Layers in Streptocarpus formosus (Gesneriaceae). Horticulturae 2021, 7, 120. [Google Scholar] [CrossRef]
- Jimoh, M.O.; Afolayan, A.J.; Lewu, F.B. Nutrients and Antinutrient Constituents of Amaranthus caudatus L. Cultivated on Different Soils. Saudi. J. Biol. Sci. 2020, 27, 3570–3580. [Google Scholar] [CrossRef]
- Muñoz-Arrieta, R.; Esquivel-Alvarado, D.; Alfaro-Viquez, E.; Alvarez-Valverde, V.; Krueger, C.G.; Reed, J.D. Nutritional and Bioactive Composition of Spanish, Valencia, and Virginia Type Peanut Skins. J. Food Compos. Anal. 2021, 98, 103816. [Google Scholar] [CrossRef]
- AOAC (Association of Official Analytical Chemist). Official Methods of Analysis; Latimer, G.W., Jr., Ed.; AOAC International: Washinton, DC, USA, 2016; ISBN 0935584870. [Google Scholar]
- Adegbaju, O.D.; Otunola, G.A.; Afolayan, A.J. Proximate, Mineral, Vitamin and Anti-Nutrient Content of Celosia Argentea at Three Stages of Maturity. S. Afr. J. Bot. 2019, 124, 372–379. [Google Scholar] [CrossRef]
- Yanu, P.; Jakmunee, J. Down Scaled Kjeldahl Digestion and Flow Injection Conductometric System for Determination of Protein Content in Some Traditional Northern Thai Foods. Food Chem. 2017, 230, 572–577. [Google Scholar] [CrossRef] [PubMed]
- USDA National Nutrient Database for Standard Reference Release. In Proceedings of the Full Report (All Nutrients) 11003, Amaranth Leaves, Raw, Beltsville, MD, USA, September 2008; pp. 1–3.
- FAO. Food Energy—Methods of Analysis and Conversion Factors. In Proceedings of the Report of a Technical Workshop on Food Energy, Rome, Italy, 3–6 December 2002; Tontisirin, K., Ed.; FAO: Rome, Italy, 2003; Volume 77, pp. 1–93. [Google Scholar]
- Tylutki, T.P.; Fox, D.G.; Durbal, V.M.; Tedeschi, L.O.; Russell, J.B.; van Amburgh, M.E.; Overton, T.R.; Chase, L.E.; Pell, A.N. Cornell Net Carbohydrate and Protein System: A Model for Precision Feeding of Dairy Cattle. Anim. Feed Sci. Technol. 2008, 143, 174–202. [Google Scholar] [CrossRef]
- Jimoh, M.O.; Afolayan, A.J.; Lewu, F.B. Antioxidant and Phytochemical Activities of Amaranthus Caudatus L. Harvested from Different Soils at Various Growth Stages. Sci. Rep. 2019, 9, 12965. [Google Scholar] [CrossRef] [Green Version]
- Espinoza, F.; Vidal, S.; Rautenbach, F.; Lewu, F.; Nchu, F. Effects of Beauveria bassiana (Hypocreales) on Plant Growth and Secondary Metabolites of Extracts of Hydroponically Cultivated Chive (Allium schoenoprasum L. [Amaryllidaceae]). Heliyon 2019, 5, e03038. [Google Scholar] [CrossRef] [Green Version]
- Mazza, G.; Fukumoto, L.; Delaquis, P.; Girard, B.; Ewert, B. Anthocyanins, Phenolics, and Color of Cabernet Franc, Merlot, and Pinot Noir Wines from British Columbia. J. Agric. Food Chem. 1999, 47, 4009–4017. [Google Scholar] [CrossRef] [PubMed]
- Benzie, I.F.F.; Strain, J.J. Ferric Reducing/Antioxidant Power Assay: Direct Measure of Total Antioxidant Activity of Biological Fluids and Modified Version for Simultaneous Measurement of Total Antioxidant Power and Ascorbic Acid Concentration. Methods Enzym. 1999, 299, 15–27. [Google Scholar] [CrossRef]
- Jimoh, M.A.; Idris, O.A.; Jimoh, M.O. Cytotoxicity, Phytochemical, Antiparasitic Screening, and Antioxidant Activities of Mucuna pruriens (Fabaceae). Plants 2020, 9, 1249. [Google Scholar] [CrossRef] [PubMed]
- Sogoni, A.; Jimoh, M.O.; Kambizi, L.; Laubscher, C.P. The Impact of Salt Stress on Plant Growth, Mineral Composition, and Antioxidant Activity in Tetragonia Decumbens Mill.: An Underutilized Edible Halophyte in South Africa. Horticulturae 2021, 7, 140. [Google Scholar] [CrossRef]
- Unuofin, J.O.; Otunola, G.A.; Afolayan, A.J. Phytochemical Screening and in Vitro Evaluation of Antioxidant and Antimicrobial Activities of Kedrostis africana (L.) Cogn. Asian Pac. J. Trop. Biomed. 2017, 7, 901–908. [Google Scholar] [CrossRef]
- Oosterhuis, D.M.; Loka, D.A.; Kawakami, E.M.; Pettigrew, W.T. The Physiology of Potassium in Crop Production. Adv. Agron. 2014, 126, 203–233. [Google Scholar] [CrossRef]
- Sanyal, S.K.; Rajasheker, G.; Kishor, P.B.K.; Kumar, S.A.; Kumari, P.H.; Saritha, K.V.; Rathnagiri, P.; Pandey, G.K. Role of Protein Phosphatases in Signaling, Potassium Transport, and Abiotic Stress Responses. In Protein Phosphatases and Stress Management in Plants; Springer: Cham, Switzerland, 2020; pp. 203–232. [Google Scholar] [CrossRef]
- Wanshnong, E.; Mounika, K.; Kumar, K.A.; Maiti, C.S. Effect of Different Levels of Potassium on Growth and Flowering of Papaya Var. Red Lady. Pharma Innov. J. 2022, 11, 1314–1317. [Google Scholar]
- Tränkner, M.; Tavakol, E.; Jákli, B. Functioning of Potassium and Magnesium in Photosynthesis, Photosynthate Translocation and Photoprotection. Physiol. Plant 2018, 163, 414–431. [Google Scholar] [CrossRef] [Green Version]
- Siddiqui, M.H.; Khan, M.N.; Mukherjee, S.; Alamri, S.; Basahi, R.A.; Al-Amri, A.A.; Alsubaie, Q.D.; Al-Munqedhi, B.M.A.; Ali, H.M.; Almohisen, I.A.A. Hydrogen Sulfide (H2S) and Potassium (K+) Synergistically Induce Drought Stress Tolerance through Regulation of H+-ATPase Activity, Sugar Metabolism, and Antioxidative Defense in Tomato Seedlings. Plant Cell Rep. 2021, 40, 1543–1564. [Google Scholar] [CrossRef]
- Jimoh, M.O.; Afolayan, A.J.; Lewu, F.B. Heavy Metal Uptake and Growth Characteristics of Amaranthus caudatus L. under Five Different Soils in a Controlled Environment. Not. Bot. Horti Agrobot. 2020, 48, 417–425. [Google Scholar] [CrossRef] [Green Version]
- Davarpanah, S.; Tehranifar, A.; Davarynejad, G.; Abadía, J.; Khorasani, R. Effects of Foliar Applications of Zinc and Boron Nano-Fertilizers on Pomegranate (Punica granatum Cv. Ardestani) Fruit Yield and Quality. Sci. Hortic. 2016, 210, 57–64. [Google Scholar] [CrossRef] [Green Version]
- Norozi, M.; ValizadehKaji, B.; Karimi, R.; Nikoogoftar Sedghi, M. Effects of Foliar Application of Potassium and Zinc on Pistachio (Pistacia vera L.) Fruit Yield. Int. J. Hortic. Sci. Technol. 2019, 6, 113–123. [Google Scholar] [CrossRef]
- Jiménez-Aguilar, D.M.; Grusak, M.A. Minerals, Vitamin C, Phenolics, Flavonoids and Antioxidant Activity of Amaranthus Leafy Vegetables. J. Food Compos. Anal. 2017, 58, 33–39. [Google Scholar] [CrossRef] [Green Version]
- Jimoh, M.O.; Afolayan, A.J.; Lewu, F.B. Suitability of Amaranthus Species for Alleviating Human Dietary Deficiencies. S. Afr. J. Bot. 2018, 115, 65–73. [Google Scholar] [CrossRef]
- Stein, A.J. Global Impacts of Human Mineral Malnutrition. Plant Soil. 2010, 335, 133–154. [Google Scholar] [CrossRef]
- Jobin, K.; Müller, D.N.; Jantsch, J.; Kurts, C. Sodium and Its Manifold Impact on Our Immune System. Trends Immunol. 2021, 42, 469–479. [Google Scholar] [CrossRef] [PubMed]
- Serna, J.; Bergwitz, C. Importance of Dietary Phosphorus for Bone Metabolism and Healthy Aging. Nutrients 2020, 12, 3001. [Google Scholar] [CrossRef] [PubMed]
- Ogundola, A.F.; Bvenura, C.; Afolayan, A.J. Nutrient and Antinutrient Compositions and Heavy Metal Uptake and Accumulation in S. Nigrum Cultivated on Different Soil Types. Sci. World J. 2018, 2018, 5703929. [Google Scholar] [CrossRef] [Green Version]
- Gopalakrishnan, L.; Doriya, K.; Kumar, D.S. Moringa Oleifera: A Review on Nutritive Importance and Its Medicinal Application. Food Sci. Hum. Wellness 2016, 5, 49–56. [Google Scholar] [CrossRef] [Green Version]
- Ghanbarzadeh, M.; Moazami, N.; Shahavi, M.H.; Mirdamadi, S. Study of Bioactive Compounds in Arthrospira Platensis MGH-1 Fortified with Micronutrients of Iron, Zinc, and Manganese. J. Appl. Phycol. 2022, 34, 2449–2462. [Google Scholar] [CrossRef]
- Kaur, M.; Kaur, R.; Gill, B.S. Mineral and Amino Acid Contents of Different Flaxseed Cultivars in Relation to Its Selected Functional Properties. J. Food Meas. Charact. 2017, 11, 500–511. [Google Scholar] [CrossRef]
- Radha; Kumar, M.; Puri, S.; Pundir, A.; Bangar, S.P.; Changan, S.; Choudhary, P.; Parameswari, E.; Alhariri, A.; Samota, M.K.; et al. Evaluation of Nutritional, Phytochemical, and Mineral Composition of Selected Medicinal Plants for Therapeutic Uses from Cold Desert of Western Himalaya. Plants 2021, 10, 1429. [Google Scholar] [CrossRef]
- Schroeder, B.O.; Bäckhed, F. Signals from the Gut Microbiota to Distant Organs in Physiology and Disease. Nat. Med. 2016, 22, 1079–1089. [Google Scholar] [CrossRef]
- Sultana, S. Nutritional and Functional Properties of Moringa Oleifera. Metabol. Open 2020, 8, 100061. [Google Scholar] [CrossRef]
- Bvenura, C.; Sivakumar, D. The Role of Wild Fruits and Vegetables in Delivering a Balanced and Healthy Diet. Food Res. Int. 2017, 99, 15–30. [Google Scholar] [CrossRef] [PubMed]
- Pinto, T.; Vilela, A.; Cosme, F. Chemical and Sensory Characteristics of Fruit Juice and Fruit Fermented Beverages and Their Consumer Acceptance. Beverages 2022, 8, 33. [Google Scholar] [CrossRef]
- Akbari, B.; Baghaei-Yazdi, N.; Bahmaie, M.; Mahdavi Abhari, F. The Role of Plant-Derived Natural Antioxidants in Reduction of Oxidative Stress. BioFactors 2022, 48, 611–633. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, S.H.; Salama, D.M.; El-Tanahy, A.; El-Bassiony, A.M. Effects of prolonged restriction in water supply and spraying with potassium silicate on growth and productivity of potato. Plant Arch. 2019, 19, 2585–2595. [Google Scholar]
- Zikalala, B.O.; Nkomo, M.; Araya, H.; Ngezimana, W.; Mudau, F.N. Nutritional Quality of Baby Spinach (Spinacia oleracea L.) as Affected by Nitrogen, Phosphorus and Potassium Fertilisation. S. Afr. J. Plant Soil 2017, 34, 79–86. [Google Scholar] [CrossRef]
- Bulgari, R.; Franzoni, G.; Ferrante, A. Biostimulants Application in Horticultural Crops under Abiotic Stress Conditions. Agronomy 2019, 9, 306. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| K2SO4 | Pruning | Plant Height | Flower Bud No. | Fresh Flower Buds (g) | Dry Flower Buds (g) |
|---|---|---|---|---|---|
| 0 M | Unpruned | 72.14 ± 1.85 a | 5.30 ± 0.63 ab | 16.14 ± 2.34 bc | 13.59 ± 2.16 b |
| 5 cm | 31.22 ± 0.77 efg | 2.80 ± 0.29 bcdef | 8.01 ± 2.31 e | 6.19 ± 2.11 d | |
| 10 cm | 30.29 ± 2.24 def | 4.60 ± 0.69 bc | 9.76 ± 2.65 e | 8.30 ± 2.43 c | |
| 15 cm | 34.23 ± 1.40 cd | 3.30 ± 0.58 bcdef | 15.83 ± 3.23 c | 13.71 ± 2.94 b | |
| 0.0072 M | Unpruned | 78.91 ± 1.65 a | 7.80 ± 0.49 a | 22.95 ± 3.44 a | 20.06 ± 3.14 a |
| 5 cm | 37.00 ± 1.14 de | 3.60 ± 0.37 bcde | 5.83 ± 1.16 f | 4.11 ± 094 de | |
| 10 cm | 44.27 ± 1.68 cd | 4.70 ± 0.78 bc | 17.75 ± 4.42 b | 14.31 ± 3.83 b | |
| 15 cm | 49.43 ± 1.57 bc | 7.60 ± 0.65 a | 22.09 ± 4.01 a | 19.54 ± 3.65 a | |
| 0.0144 M | Unpruned | 55.57 ± 2.28 b | 4.00 ± 0.52 bcd | 11.06 ± 2.22 de | 7.93 ± 1.85 c |
| 5 cm | 27.71 ± 0.85 efg | 1.80 ± 0.47 defg | 5.54 ± 2.32 f | 4.16 ± 1.86 de | |
| 10 cm | 25.50 ± 3.59 fgh | 2.70 ± 0.67 cdefg | 9.28 ± 1.92 e | 7.35 ± 1.66 c | |
| 15 cm | 27.52 ± 4.74 efg | 1.00 ± 0.45 fg | 4.95 ± 2.05 f | 3.64 ± 1.54 e | |
| 0.0216 M | Unpruned | 49.19 ± 2.43 bc | 2.70 ± 0.34 cdefg | 8.75 ± 1.11 e | 6.04 ± 0.85 d |
| 5 cm | 13.11 ± 2.80 i | 0.20 ± 0.13 g | 0.42 ± 0.42 g | 0.23 ± 0.23 f | |
| 10 cm | 15.29 ± 2.24 hi | 1.30 ± 0.54 g | 2.49 ± 1.28 g | 1.45 ± 0.45 f | |
| 15 cm | 21.51 ± 0.63 hi | 1.10 ± 0.28 g | 1.64 ± 0.5 g | 0.86 ± 0.30 f | |
| Two-way ANOVA F-Statistic | |||||
| K2SO4 | 17,272 * | 483.67 * | 4275 * | 3495 * | |
| Pruning | 34,194 * | 158.22 * | 1903 * | 1411 * | |
| K2SO4 × Pruning | 1582 * | 95.61 * | 1156 * | 1018 * | |
| K2SO4 | Pruning | Shoot Fresh Weight (g) | Shoot Dry Weight (g) | Root Fresh Weight (g) | Root Dry Weight (g) |
|---|---|---|---|---|---|
| 0 M | Unpruned | 93.48 ± 19.3 bc | 16.41 ± 2.3 b | 51.30 ± 2.39 bc | 27.89 ± 1.31 abcd |
| 5 cm | 60.4 ± 9.33 bcde | 6.72 ± 1.76 d | 30.98 ± 1.20 de | 21.27 ± 1.72 def | |
| 10 cm | 69.8 ± 12.5 cd | 17.27 ± 3.75 b | 30.86 ± 1.13 de | 22.80 ± 1.78 cde | |
| 15 cm | 114.5 ± 7.79 ab | 19.19 ± 4.8 ab | 46.06 ± 1.41 cd | 34.08 ± 2.25 abc | |
| 0.0072 M | Unpruned | 154.3 ± 12.6 a | 19.38 ± 3.76 a | 63.23 ± 2.68 a | 38.17 ± 3.81 a |
| 5 cm | 51.65 ± 4.96 d | 8.65 ± 2.14 cd | 30.98 ± 1.20 ef | 25.60 ± 0.84 bcd | |
| 10 cm | 91.0 ± 13.8 bc | 8.76 ± 2.51 cd | 21.22 ± 1.26 e | 34.86 ± 1.33 def | |
| 15 cm | 89.5 ± 16.0 bc | 15.80 ± 3.22B | 38.09 ± 2.31 de | 26.84 ± 2.85 abcd | |
| 0.0144 M | Unpruned | 97.9 ± 17.4 b | 11.13 ± 2.25 bc | 51.77 ± 3.05 bc | 35.78 ± 3.76 ab |
| 5 cm | 25.80 ± 5.56 f | 6.79 ± 2.38 d | 59.92 ± 3.30 ab | 21.27 ± 1.72 bcd | |
| 10 cm | 45.0 ± 12.4 e | 9.28 ± 1.92 cd | 30.50 ± 3.59 ef | 17.34 ± 2.60 def | |
| 15 cm | 50.0 ± 15.6 de | 4.95 ± 2.05 e | 22.93 ± 2.00 fg | 12.17 ± 2.83 efg | |
| 0.0216 M | Unpruned | 21.53 ± 4.10 f | 8.75 ± 1.11 cd | 32.88 ± 1.33 ef | 10.91 ± 1.67 fg |
| 5 cm | 5.91 ± 2.26 g | 0.42 ± 0.42 e | 14.32 ± 0.34 g | 2.62 ± 0.89 g | |
| 10 cm | 3.81 ± 1.82 g | 2.49 ± 1.28 e | 19.19 ± 0.58 g | 3.45 ± 1.57 g | |
| 15 cm | 4.47 ± 3.67 g | 1.64 ± 0.57 e | 24.49 ± 0.95 fg | 1.95 ± 1.41 g | |
| Two-way ANOVA F-Statistic | |||||
| K2SO4 | 18,136 * | 3541 * | 11,307 * | 13,755 * | |
| Pruning | 80,109 * | 1276 * | 8830 * | 3393 * | |
| K2SO4 × Pruning | 23,891 * | 1017 * | 9949 * | 2825 * | |
| (a) | ||||||||
| K2SO4 | Pruning | Ca (mg/100 g DW) | Mg (mg/100 g DW) | Na (mg/100 g DW) | P (mg/100 g DW) | N (mg/100 g DW) | K (mg/100 g DW) | K/Ca + Mg (mg/100 g DW) |
| 0 M | Unpruned | 610 ± 0.01 a | 340 ± 0.01 ab | 220 ± 0.00 c | 740 ± 0.01 ab | 3790 ± 0.07 b | 5700 ± 0.01 hi | 2530 ± 0.05 h |
| 5 cm | 480 ± 0.01 a | 340 ± 0.01 ab | 190 ± 0.00 cde | 700 ± 0.01 abc | 3630 ± 0.07 b | 5460 ± 0.02 ij | 2580 ± 0.09 h | |
| 10 cm | 650 ± 0.01 a | 370 ± 0.02 a | 180 ± 0.01 def | 700 ± 0.01 abc | 3800 ± 0.00 b | 5130 ± 0.01 jk | 2030 ± 0.017 i | |
| 15 cm | 640 ± 0.01 a | 350 ± 0.00 a | 210 ± 0.01 cd | 660 ± 0.02 cde | 3690 ± 0.05 b | 5130 ± 0.09 jk | 2160 ± 0.00 i | |
| 0.0072 M | Unpruned | 260 ± 0.01 a | 170 ± 0.01 e | 120 ± 0.01 h | 670 ± 0.01 cd | 3550 ± 0.08 b | 9460 ± 0.78 a | 9490 ± 0.02 a |
| 5 cm | 440 ± 0.01 a | 280 ± 0.28 bc | 130 ± 0.00 gh | 750 ± 0.01 a | 3690 ± 0.05 b | 6490 ± 0.03 ef | 3750 ± 0.05 g | |
| 10 cm | 250 ± 0.01 a | 210 ± 0.01 cde | 120 ± 0.00 gh | 710 ± 0.01 abc | 3770 ± 0.06 b | 7090 ± 0.04 cd | 5680 ± 0.14 b | |
| 15 cm | 390 ± 0.01 a | 250 ± 0.01 cd | 130 ± 0.01 gh | 740 ± 0.01 ab | 3700 ± 0.058 b | 7530 ± 0.07 b | 7530 ± 0.07 d | |
| 0.0144 M | Unpruned | 280 ± 0.01 a | 240 ± 0.01 cde | 270 ± 0.01 b | 670 ± 0.24 cd | 4300 ± 0.06 a | 6460 ± 0.03 ef | 4930 ± 0.04 d |
| 5 cm | 330 ± 0.01 a | 250 ± 0.01 cd | 140 ± 0.01 fgh | 660 ± 0.15 cde | 4340 ± 0.02 a | 5820 ± 0.08 h | 4180 ± 0.04 ef | |
| 10 cm | 310 ± 0.01 a | 260 ± 0.00 cd | 340 ± 0.01 a | 760 ± 0.01 a | 4470 ± 0.09 a | 7160 ± 0.07 bc | 4950 ± 0.01 d | |
| 15 cm | 270 ± 0.01 a | 230 ± 0.00 cde | 180 ± 0.01 cde | 620 ± 0.01 def | 4300 ± 0.06 b | 6730 ± 0.03 def | 5350 ± 0.02 c | |
| 0.0216 M | Unpruned | 230 ± 0.01 a | 220 ± 0.01 cde | 270 ± 0.01 b | 590 ± 0.01 ef | 2970 ± 0.03 cd | 6360 ± 0.04 fg | 3910 ± 0.06 fg |
| 5 cm | 700 ± 10.2 a | 210 ± 0.01 cde | 130 ± 0.01 gh | 680 ± 0.02 bcd | 3530 ± 0.145 b | 3530 ± 0.15 k | 4150 ± 0.02 ef | |
| 10 cm | 500 ± 0.88 a | 270 ± 0.03 bc | 300 ± 0.01 b | 560 ± 0.02 cde | 3170 ± 0.07 c | 6770 ± 0.13 de | 4400 ± 0.05 e | |
| 15 cm | 240 ± 0.01 a | 190 ± 0.01 de | 160 ± 0.01 efg | 550 ± 0.02 f | 2800 ± 0.06 d | 6070 ± 0.027 fg | 4760 ± 0.04 d | |
| Two-way ANOVA F-Statistic | ||||||||
| K2SO4 | 1.06 ns | 75.85 * | 137.70 * | 43.30 * | 181.01 * | 706.71 * | 2898 * | |
| Pruning | 0.96 ns | 5.40 * | 100.60 * | 19.48 * | 21.50 * | 228.54 * | 519 * | |
| K2SO4 × pruning | 0.95 ns | 4.72 * | 45.47 * | 11.32 * | 9.99 * | 92.39 * | 532 * | |
| (b) | ||||||||
| K2SO4 | Pruning | Zn (mg/100 g DW) | Mn (mg/100 g DW) | Cu (mg/100 g DW) | Fe (mg/100 g DW) | |||
| 0 M | Unpruned | 10.8 ± 1.15 bcd | 4.23 ± 1.20 def | 0.5. ± 0.00 a | 12.3 ± 0.88 abc | |||
| 5 cm | 8.9 ± 0.58 ef | 2.9 ± 0.88 jk | 0.373 ± 0.15 bc | 12.23 ± 0.67 bc | ||||
| 10 cm | 8.4. ± 1.15 fg | 3.36 ± 0.33 ij | 0.464 ± 0.31 ab | 8.9 ± 0.58 fg | ||||
| 15 cm | 10.2 ± 1.15 bcd | 3.8 ± 1.00 gh | 0.5 ± 0.00 a | 8.72 ± 0.64 fg | ||||
| 0.0072 M | Unpruned | 10.9 ± 1.53 bc | 6.7 ± 0.58 a | 0.2 ± 0.00 d | 9.23 ± 2.03 ef | |||
| 5 cm | 8.6 ± 0.88 f | 4.4 ± 1.15 de | 0 ± 0.00 f | 7.1 ± 0.58 h | ||||
| 10 cm | 7.3 ± 0.58 h | 4.9 ± 0.58 bc | 0.1 ± 0.00 e | 5.6 ± 1.53 i | ||||
| 15 cm | 11 ± 0.58 b | 5.2 ± 1.20 b | 0.1 ± 0.00 e | 8.2 ± 1.13 g | ||||
| 0.0144 M | Unpruned | 10.8 ± 1.53 bcd | 3.8 ± 0.58 gh | 0.4 ± 0.00 bc | 13 ± 0.58 ab | |||
| 5 cm | 10.1 ± 0.58 bcd | 4.2 ± 0.58 efg | 0.4 ± 0.00 bc | 8.1 ± 1.15 g | ||||
| 10 cm | 12.6 ± 0.88 a | 4.03 ± 0.35 efgh | 0.4 ± 0.00 bc | 12 ± 4.51 c | ||||
| 15 cm | 9.8 ± 0.58 de | 2.9 ± 0.58 k | 0.2 ± 0.00 d | 13.3 ± 2.65 a | ||||
| 0.0216 M | Unpruned | 10.16 ± 1.67 bcd | 3.9 ± 0.58 fgh | 0.537 ± 0.34 a | 10.7 ± 3.71 d | |||
| 5 cm | 9.83 ± 6.94 cde | 4.633 ± 0.88 cd | 0.357 ± 0.32 c | 8.8 ± 0.56 fg | ||||
| 10 cm | 8.86 ± 0.33 ef | 3.7 ± 0.58 hi | 0.370 ± 0. 21 bc | 9.43 ± 0.89 ef | ||||
| 15 cm | 7.5 ± 2.52 gh | 2.8 ± 0.00 k | 0.467 ± 0.41 ab | 10.13 ± 0.89 de | ||||
| Two-way ANOVA F-Statistic | ||||||||
| K2SO4 | 2017.8 * | 2364.29 * | 96.497 * | 10,710.7 * | ||||
| Pruning | 1456.4 * | 597. 59 * | 10.313 * | 4358.4 * | ||||
| K2SO4 × pruning | 5253.8 * | 1365.23 * | 14.407 * | 7284.4 * | ||||
| K2SO4 | Pruning Levels | % Ash | % Crude Fat | % Crude Protein | % NDF | % NFC | % Moisture | Energy Value (KJ/100 g) |
|---|---|---|---|---|---|---|---|---|
| 0 M | Unpruned | 15.7 ± 0.02 bcd | 2.2 ± 0.05 abc | 24.2 ± 0.04 cd | 33.2 ± 0.2 cd | 24.6 ± 0.2 def | 10.1 ± 0.04 g | 1476.4 ± 0.3 ab |
| 5 cm | 14.9 ± 0.20 cd | 2.3 ± 0.08 abc | 23.6 ± 0.04 cde | 31.8 ± 0.4 ef | 27.2 ± 0.4 bcd | 10.3 ± 0.1 efg | 1491.1 ± 2.2 ab | |
| 10 cm | 14.5 ± 0.20 d | 2.0 ± 0.05 bc | 24.1 ± 0.01 cd | 34.6 ± 0.1 bc | 24.7 ± 01 de | 10.5 ± 0.07 defg | 1493.5 ± 6 a | |
| 15 cm | 14.0 ± 2.30 d | 2.0 ± 0.02 bc | 23.46 ± 0.1 de | 35.3 ± 0.2 b | 25.1 ± 2.5 de | 10.23 ± 0.08 fg | 1503.2 ± 3.2 a | |
| 0.0072 M | Unpruned | 23.6 ± 0.30 a | 1.9 ± 0.02 bc | 22.3 ± 0.2 ef | 37.8 ± 0.2 a | 14.2 ± 0.4 g | 10.9 ± 0.03 cde | 1335.7 ± 0.5 d |
| 5 cm | 16.0 ± 2.10 bcd | 2.0 ± 0.01 bc | 23.7 ± 0.4 cde | 32.3 ± 0.3 de | 25.8 ± 2.1 cde | 10.8 ± 0.1 cdef | 1469 ± 0.3 abc | |
| 10 cm | 19.1 ± 0.05 bc | 2.1 ± 0.06 abc | 23.7 ± 0.1 cde | 31.6 ± 0.3 ef | 23.3 ± 0.2 def | 12.2 ± 0.1 a | 1417.4 ± 0.2 bc | |
| 15 cm | 20.0 ± 0.04 ab | 1.8 ± 0.01 bc | 23.19 ± 0.2 def | 35.05 ± 0.08 b | 19.9 ± 0.3 f | 11.8 ± 0.08 ab | 1395 ± 0.4 cd | |
| 0.0144 M | Unpruned | 16.5 ± 0.30 bcd | 2.3 ± 0.08 ab | 25.1 ± 0.3 bc | 31.94 ± 0.08 def | 24.0 ± 0.6 def | 11.4 ± 0.2 bc | 1464.9 ± 4 abc |
| 5 cm | 14.8 ± 0.10 cd | 1.9 ± 0.02 bc | 26.6 ± 0.3 a | 30. ± 0.1 gh | 26.5 ± 0.2 cde | 11.8 ± 0.01 ab | 1486.8 ± 1.8 ab | |
| 10 cm | 16.8 ± 0.01 bcd | 2.3 ± 0.1 ab | 26.0 ± 0.1 ab | 32.6 ± 0.3 de | 22.1 ± 0.4 ef | 11.5 ± 0.1 bc | 1459.9 ± 2.1 abc | |
| 15 cm | 17.1 ± 0.07 bcd | 2.6 ± 0.29 a | 23.5 ± 0.3 de | 31 ± 0.01 fg | 25.7 ± 0.02 cde | 11.0 ± 0.06 cd | 1461.2 ± 2.1 abc | |
| 0.0216 M | Unpruned | 15.7 ± 0.20 bcd | 2.1 ± 0.08 abc | 22.5 ± 0.3 ef | 26.9 ± 0.1 j | 59.5 ± 0.5 abc | 10.9 ± 0.03 cde | 1475.5 ± 2.1 ab |
| 5 cm | 13.3 ± 0.10 d | 1.7 ± 0.06 c | 24.3 ± 0.2 cd | 30.1 ± 0.04 gh | 60.5 ± 0.2 a | 10.2 ± 0.1 fg | 1507.3 ± 3.1 a | |
| 10 cm | 15.6 ± 0.20 cd | 1.9 ± 0.01 bc | 23.2 ± 0.2 de | 28.9 ± 0.2 hi | 59.0 ± 0.1 abc | 9.9 ± 0.2 g | 1473.2 ± 3.6 ab | |
| 15 cm | 16.2 ± 0.30 bcd | 1.9 ± 0.1 bc | 21.7 ± 0.5 f | 28.2 ± 0.3 ij | 60.1 ± 0.4 ab | 10.1 ± 0.1 fg | 1462.2 ± 3.3 abc | |
| Two-way ANOVA F-Statistic | ||||||||
| K2SO4 | 28.05 * | 10.56 * | 56.51 * | 394 * | 85.82 * | 103.53 * | 29.46 * | |
| Pruning | 9.59 * | 1.32 ns | 26.97 * | 24.2 * | 10.59 * | 3.04 * | 8.56 * | |
| K2SO4 × pruning | 3.02 * | 3.80 * | 6.31 * | 58.38 * | 8.85 * | 14.15 * | 3.18 * | |
| K2SO4 | Pruning Levels | Polyphenols (mg GAE/g) | Flavonols (mg QE/g) | FRAP (µmol AAE/g) | DPPH (µmol TE/g) | TEAC (µmol TE/g) |
|---|---|---|---|---|---|---|
| 0 M | Unpruned | 3.55 ± 0.15 de | 3.59 ± 0.09 def | 14.98 ± 0. 14 gh | 5.04 ± 0.02 fg | 4.45 ± 0.09 gh |
| 5 cm | 3.04 ± 0.07 ef | 0.69 ± 0.03 j | 15.33 ± 0.48 gh | 5.37 ± 0.18 efg | 2.69 ± 0.06 j | |
| 10 cm | 4.63 ± 0.14 bc | 1.83 ± 0.12 hi | 23.02 ± 0.80 cd | 7.36 ± 0.23 cd | 4.32 ± 0.19 gh | |
| 15 cm | 4.29 ± 0.25 bcd | 1.99 ± 0.08 hi | 20.09 ± 0.86 de | 8.63 ± 0.16 bc | 19.04 ± 0.49 a | |
| 0.0072 M | Unpruned | 6.55 ± 0.05 a | 3.33 ± 0.08 ef | 19.27 ± 0.71 ef | 6.44 ± 0.18 de | 5.52 ± 0.06 def |
| 5 cm | 5.84 ± 0.08 a | 3.88 ± 0.21 cde | 33.69 ± 1.06 a | 9.34 ± 0.18 bc | 5.12 ± 0.03 efgh | |
| 10 cm | 6.13 ± 0.29 a | 5.22 ± 0.27 a | 29.48 ± 0.85 b | 11.47 ± 0.42 a | 5.87 ± 0.31 b | |
| 15 cm | 6.33 ± 0.15 a | 4.56 ± 0.29 abc | 28.34 ± 0.63 b | 9.25 ± 0.36 b | 5.87 ± 0.31 bc | |
| 0.0144 M | Unpruned | 6.57 ± 0.19 a | 2.14 ± 0.04 hi | 10.09 ± 0.55 i | 3.54 ± 0.09 h | 5.23 ± 0.16 efg |
| 5 | 2.69 ± 0.13 f | 4.29 ± 0.21 cd | 16.31 ± 0.38 fg | 6.37 ± 0.21 de | 6.74 ± 0.07 c | |
| 10 | 4.26 ± 0.07 cd | 4.33 ± 0.08 bc | 24.14 ± 0.92 c | 9.01 ± 0.32 b | 6.35 ± 0.27 cd | |
| 15 | 5.02 ± 0.13 b | 5.05 ± 0.09 ab | 19.46 ± 0.23 ef | 6.18 ± 0.22 def | 4.17 ± 0.07 hi | |
| 0.0216 M | Unpruned | 4.13 ± 0.15 cd | 2.47 ± 0.09 gh | 16.52 ± 0.71 fg | 6.39 ± 0.17 de | 3.27 ± 0.14 ij |
| 5 cm | 2.93 ± 0.06 ef | 3.04 ± 0.05 fg | 10.99 ± 0.29 i | 5.59 ± 0.47 ef | 6.82 ± 0.09 c | |
| 10 cm | 3.15 ± 0.02 ef | 1.53 ± 0.08 hi | 12.48 ± 0.55 hi | 4.31 ± 0.07 gh | 4.57 ± 0.06 fgh | |
| 15 cm | 5.23 ± 0.16 ef | 1.82 ± 0.06 hi | 12.52 ± 0.44 hi | 4.10 ± 0.09 gh | 3.29 ± 0.08 ij | |
| Two-way ANOVA F-Statistics | ||||||
| K2SO4 | 36.97 * | 47.58 * | 726.08 * | 68.19 * | 74.58 * | |
| Pruning | 26.77 * | 1.20 * | 704.83 * | 57.81 * | 187.34 * | |
| K2SO4 × Pruning | 26.56 * | 36.38 * | 751.74 * | 97.04 * | 476.14 * | |
| NFT/Table | Gutter 1 | Gutter 2 | Gutter 3 | Gutter 4 |
|---|---|---|---|---|
| 1 | 5 cm + 0 M | 10 cm + 0 M | 15 cm + 0 M | Unpruned + 0 M |
| 2 | 5 cm + 0.0072 M | 10 cm + 0.0072 M | 15 cm + 0.0072 M | Unpruned + 0.0072 M |
| 3 | 5 cm + 0.0144 M | 10 cm + 0.0144 M | 15 cm + 0.0144 M | Unpruned + 0.0144 M |
| 4 | 5 cm + 0.0216 M | 10 cm + 0.0216 M | 15 cm + 0.0216 M | Unpruned + 0.0216 M |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bulawa, B.; Sogoni, A.; Jimoh, M.O.; Laubscher, C.P. Potassium Application Enhanced Plant Growth, Mineral Composition, Proximate and Phytochemical Content in Trachyandra divaricata Kunth (Sandkool). Plants 2022, 11, 3183. https://doi.org/10.3390/plants11223183
Bulawa B, Sogoni A, Jimoh MO, Laubscher CP. Potassium Application Enhanced Plant Growth, Mineral Composition, Proximate and Phytochemical Content in Trachyandra divaricata Kunth (Sandkool). Plants. 2022; 11(22):3183. https://doi.org/10.3390/plants11223183
Chicago/Turabian StyleBulawa, Bakholise, Avela Sogoni, Muhali Olaide Jimoh, and Charles Petrus Laubscher. 2022. "Potassium Application Enhanced Plant Growth, Mineral Composition, Proximate and Phytochemical Content in Trachyandra divaricata Kunth (Sandkool)" Plants 11, no. 22: 3183. https://doi.org/10.3390/plants11223183