
Varietal Resistance and Chemical Ecology of the Rice Stink Bug, Oebalus pugnax, on Rice, Oryza sativa

Abstract
:1. Introduction
2. Results
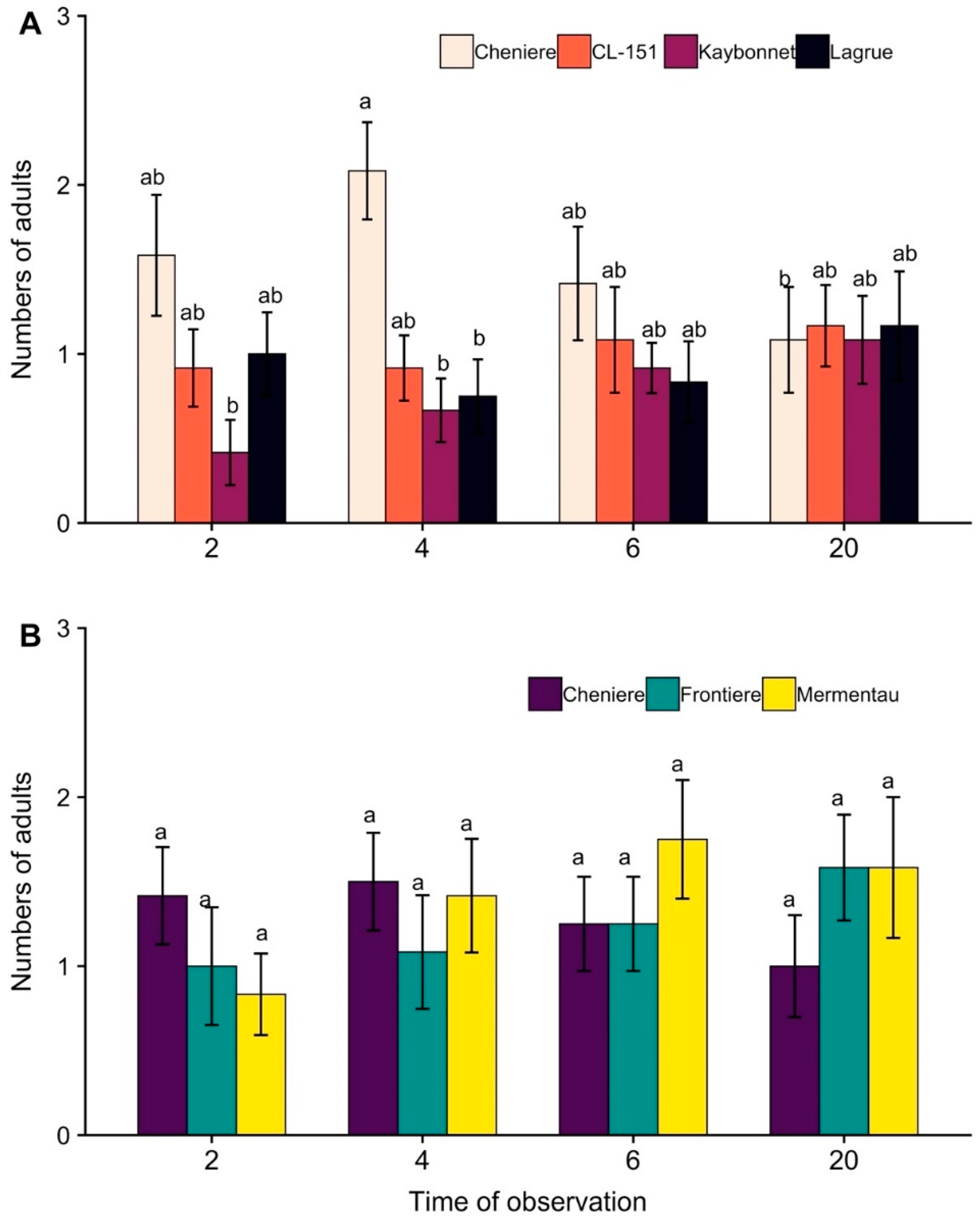
2.1. Varietal Preference Study
2.2. Varietal Performance Study
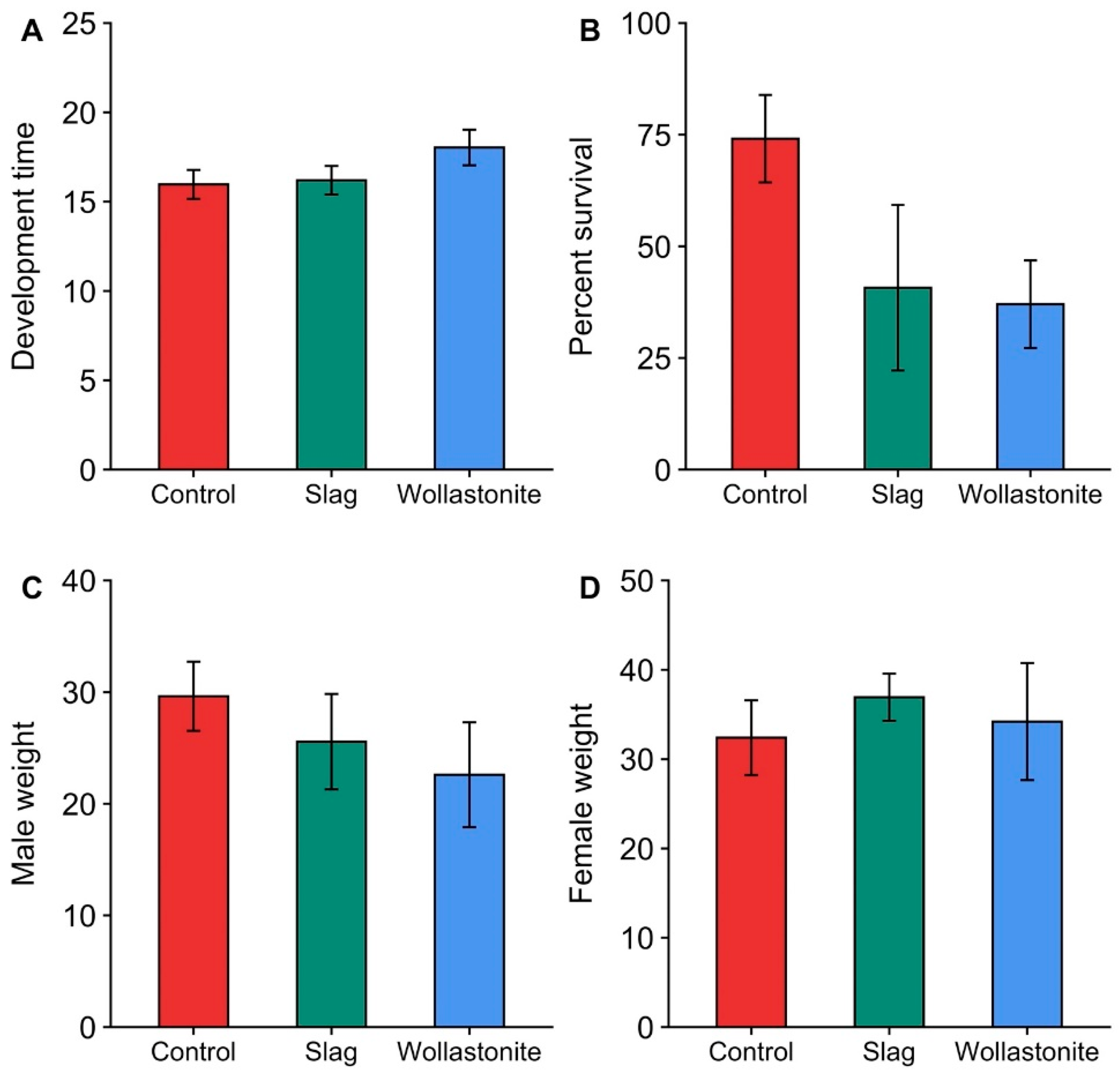
2.3. Silicon Study
2.4. Chemical Ecology of Dorsal Abdominal Gland and Metathoracic Gland Extracts
3. Discussion
3.1. Varietal Preference Study
3.2. Varietal Performance Study
3.3. Silicon Study
3.4. Chemical Ecology of Dorsal Abdominal Glands and Metathoracic Glands
4. Materials and Methods
4.1. Insect Rearing
4.2. Rice Varieties and Plants
4.3. Varietal Preference Study
4.4. Varietal Performance Study
4.5. Silicon Study
4.6. Extraction of Dorsal Abdominal Glands and Metathoracic Glands
4.7. Data Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mammadov, J.; Buyyarapu, R.; Guttikonda, S.K.; Parliament, K.; Abdurakhmonov, I.Y.; Kumpatla, S.P. Wild relatives of maize, rice, cotton, and soybean: Treasure troves for tolerance to biotic and abiotic stresses. Front. Plant Sci. 2018, 9, 886. [Google Scholar] [CrossRef] [PubMed]
- Way, M.O. Rice arthropod pests and their management in the United States. In Rice: Origin, History, Technology, and Production; Smith, C.W., Dilday, R.H., Eds.; Wiley: Hoboken, NJ, USA, 2003; pp. 437–456. [Google Scholar]
- Patel, D.T.; Stout, M.J.; Fuxa, J.R. Effects of rice panicle age on quantitative and qualitative injury by the rice stink bug (Hemiptera: Pentatomidae). Fla. Entomol. 2006, 89, 321–327. [Google Scholar] [CrossRef]
- Espino, L.; Way, M.O. Relative susceptibility of stages of rice panicle development to male and female Oebalus pugnax. Southwest. Entomol. 2007, 32, 203–211. [Google Scholar] [CrossRef]
- Awuni, G.A.; Gore, J.; Cook, D.; Musser, F.; Catchot, A.; Dobbins, C. Impact of Oebalus pugnax (Hemiptera: Pentatomidae) infestation timing on rice yields and quality. J. Econ. Entomol. 2015, 108, 1739–1747. [Google Scholar] [CrossRef] [Green Version]
- Blackman, B.; Lanka, S.; Hummel, N.; Way, M.; Stout, M. Comparison of the effects of neonicotinoids and pyrethroids against Oebalus pugnax (Hemiptera: Pentatomidae) in rice. Fla. Entomol. 2015, 98, 18–26. [Google Scholar] [CrossRef]
- Moldenhauer, K.A.K.; Scott, B.; Hardke, J. BR Wells Arkansas Rice Research Studies 2019; Arkansas Agricultural Experiment Station Research Series; University of Arkansas: Fayetteville, AR, USA, 2020; Available online: https://scholarworks.uark.edu/aaesser/163 (accessed on 17 May 2022).
- Lidova, J.; Buric, M.; Kouba, A.; Velisek, J. Acute toxicity of two pyrethroid insecticides for five non-indigenous crayfish species in Europe. Vet. Med. 2019, 64, 125–133. [Google Scholar] [CrossRef] [Green Version]
- Sudarsono, H.; Bernhardt, J.L.; Tugwell, N.P. Survival of immature Telenomus podisi (Hymenoptera: Scelionidae) and rice stink bug (Hemiptera: Pentatomidae) embryos after field applications of methyl parathion and carbaryl. J. Econ. Entomol. 1992, 85, 375–378. [Google Scholar] [CrossRef]
- Smith, C.M. Plant Resistance to Arthropods: Molecular and Conventional Approaches; Springer Press: Dordrecht, The Netherlands, 2005. [Google Scholar]
- Nilakhe, S.S. Rice lines screened for resistance to the rice stink bug. J. Econ. Entomol. 1976, 69, 703–705. [Google Scholar] [CrossRef]
- Bernhardt, J.L.; Moldenhauer, K.A.K.; Gibbons, J.W. Screening rice lines for susceptibility to rice stink bug: Results from the Arkansas rice performance tests. In Pest Managemet: Insects B R Wells Rice Research Studies; Arkansas Agricultural Experimental Station Research Series; University of Arkansas: Fayetteville, AR, USA, 2004; Volume 529, pp. 159–249. [Google Scholar]
- Reynolds, O.L.; Keeping, M.G.; Meyer, J.H. Silicon augmented resistance of plants to herbivorous insects: A review. Ann. Appl. Biol. 2009, 155, 171–186. [Google Scholar] [CrossRef]
- Frew, A.; Weston, L.A.; Reynolds, O.L.; Gurr, G.M. The role of silicon in plant biology: A paradigm shift in research approach. Ann. Bot. 2018, 121, 1265–1273. [Google Scholar] [CrossRef]
- Ma, J.F.; Yamaji, N. Silicon uptake and accumulation in higher plants. Trends Plant Sci. 2006, 11, 392–397. [Google Scholar] [CrossRef] [PubMed]
- Sidhu, J.K.; Stout, M.J.; Blouin, D.C.; Datnoff, L.E. Effect of silicon soil amendment on performance of sugarcane borer, Diatraea saccharalis (Lepidoptera: Crambidae) on rice. Bull. Entomol. Res. 2013, 103, 656–664. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Han, Y.Q.; Li, P.; Wen, L.Z.; Hou, M.L. Silicon amendment to rice plants impairs sucking behaviors and population growth in the phloem feeder Nilaparvata lugens (Hemiptera: Delphacidae). Sci. Rep. 2017, 7, 1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhavanam, S.; Stout, M.J. Assessment of silicon and mycorrhizae mediated constitutive and induced systemic resistance in rice, Oryza sativa L., against the Fall armyworm, Spodoptera frugiperda Smith. Plants 2021, 10, 2126. [Google Scholar] [CrossRef]
- Lavinsky, A.O.; Detmann, K.C.; Reis, J.V.; Avila, R.T.; Sanglard, M.L.; Pereira, L.F.; Sanglard, L.M.V.P.; Rodrigues, F.A.; Araujo, W.L.; DaMatta, F.M. Silicon improves rice grain yield and photosynthesis specifically when supplied during the reproductive growth stage. J. Plant Physiol. 2016, 206, 125–132. [Google Scholar] [CrossRef]
- Yamaji, N.; Ma, J.F. A transporter at the node responsible for intervascular transfer of silicon in rice. Plant Cell 2009, 21, 2878–2883. [Google Scholar] [CrossRef] [Green Version]
- Awuni, G.A.; Gore, J.; Cook, D.; Musser, F.; Bond, J. Seasonal abundance and phenology of Oebalus pugnax (Hemiptera: Pentatomidae) on graminaceous hosts in the delta region of Mississippi. Environ. Entomol. 2015, 44, 931–938. [Google Scholar] [CrossRef] [Green Version]
- Hokkanen, H.M.T. Trap cropping in pest management. Annu. Rev. Entomol. 1991, 36, 119–138. [Google Scholar] [CrossRef]
- Borges, M.; Aldrich, J.R. Instar-specific defensive secretions of stink bugs (Heteroptera: Pentatomidae). Experientia 1992, 48, 893–896. [Google Scholar] [CrossRef]
- Aldrich, J.R. Chemical ecology of the Heteroptera. Annu. Rev. Entomol. 1988, 33, 211–238. [Google Scholar] [CrossRef]
- Blum, M.S.; Traynham, J.G.; Chidester, J.B.; Boggus, J.D. N-tridecane and trans-2-heptenal in scent gland of the rice stink bug Oebalus pugnax (F). Science 1960, 132, 1480–1481. [Google Scholar] [CrossRef]
- Moraes, M.C.B.; Pareja, M.; Laumann, R.A.; Borges, M. The chemical volatiles (semiochemicals) produced by neotropical stink bugs (Hemiptera: Pentatomidae). Neotrop Entomol 2008, 37, 489–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borges, M.; Blassioli-Moraes, M.C. The semiochemistry of Pentatomidae. In Stink Bugs: Biorational Control Based on Communication Processes; Cokl, A., Borges, M., Eds.; CRC Press: Boca Raton, FL, USA, 2017; pp. 95–124. [Google Scholar]
- Durak, D.; Kalender, Y. Fine structure and chemical analysis of the metathoracic scent gland secretion in Graphosoma lineatum (Linnaeus, 1758) (Heteroptera, Pentatomidae). Comptes Rendus. Biol. 2009, 332, 34–42. [Google Scholar] [CrossRef]
- Lima, B.S.A.; Martínez, L.C.; Plata-Rueda, A.; dos Santos, M.H.; de Oliveira, E.E.; Zanuncio, J.C.; Serrão, J.E. Interaction between predatory and phytophagous stink bugs (Heteroptera: Pentatomidae) promoted by secretion of scent glands. Chemoecology 2021, 31, 209–219. [Google Scholar] [CrossRef]
- Onnink, C.M.; Williams, A.A.; Williams, J.S. Electrophysiological and behavioral evidence for (E)-2-hexenal as a female attracting pheromone produced by disturbed Megacopta cribraria (Hemiptera: Plataspidae). J. Entomol. Sci. 2017, 52, 39–51. [Google Scholar] [CrossRef]
- Vieira, C.R.; Blassioli-Moraes, M.C.; Borges, M.; Pires, C.S.S.; Sujii, E.R.; Laumann, R.A. Field evaluation of (E)-2-hexenal efficacy for behavioral manipulation of egg parasitoids in soybean. Biocontrol 2014, 59, 525–537. [Google Scholar] [CrossRef] [Green Version]
- Fraga, D.F.; Parker, J.; Busoli, A.C.; Hamilton, G.C.; Nielsen, A.L.; Rodriguez-Saona, C. Behavioral responses of predaceous minute pirate bugs to tridecane, a volatile emitted by the brown marmorated stink bug. J. Pest Sci. 2017, 90, 1107–1118. [Google Scholar] [CrossRef] [Green Version]
- Bhavanam, S.; Wilson, B.; Blackman, B.; Stout, M. Biology and management of the rice stink bug (Hemiptera: Pentatomidae) in rice, Oryza sativa (Poales: Poaceae). J. Integr. Pest Manag. 2021, 12, 20. [Google Scholar] [CrossRef]
- Pickett, J.A.; Wadhams, L.J.; Woodcock, C.M. Developing sustainable pest control from chemical ecology. Agric. Ecosyst. Environ. 1997, 64, 149–156. [Google Scholar] [CrossRef]
- Blassioli-Moraes, M.C.; Borges, M.; Michereff, M.F.F.; Magalhães, D.M.; Laumann, R.A. Semiochemicals from plants and insects on the foraging behavior of Platygastridae egg parasitoids. Pesqui. Agropecu. Bras. 2016, 51, 454–464. [Google Scholar] [CrossRef]
- Stout, M.J. Reevaluating the conceptual framework for applied research on host-plant resistance. Insect Sci. 2013, 20, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Johnson, D.T.; Bryant, R.J.; Bernhardt, J.L. Volatiles induction in rice stink bug host grasses and rice plants. In Pest Managemet: Insects B R Wells Rice Research Studies; Arkansas Agriculture and Extension Service Research Series; University of Arkansas: Fayetteville, AR, USA, 2006; Volume 550, pp. 178–183. [Google Scholar]
- Chiluwal, K.; Kim, J.; Do Bae, S.; Park, C.G. Essential oils from selected wooden species and their major components as repellents and oviposition deterrents of Callosobruchus chinensis (L.). J. Asia Pac. Entomol. 2017, 20, 1447–1453. [Google Scholar] [CrossRef]
- Conboy, N.J.A.; McDaniel, T.; George, D.; Ormerod, A.; Edwards, M.; Donohoe, P.; Gatehouse, A.M.R.; Tosh, C.R. Volatile organic compounds as insect repellents and plant elicitors: An integrated pest management (IPM) strategy for glasshouse whitefly (Trialeurodes vaporariorum). J. Chem. Ecol. 2020, 46, 1090–1104. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, A.K.; Prajapati, V.; Khanuja, S.P.S.; Kumar, S. Effect of d-limonene on three stored product beetles. J. Econ. Entomol. 2003, 96, 990–995. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, M.; Suzuki, T.; Yoshida, T.; Kato, T.; Oi, T.; Mitsuya, S.; Inukai, Y. The role of Oryza sativa L. ‘Milyang 44’ husks on the resistance to two rice stink bugs. Breed Sci. 2020, 70, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Showemimo, F.A. Head bugs: Components of host-plant resistance in sorghum (Sorghum bicolor (L.) Moench). J. Food Agric. Environ. 2003, 1, 270–272. [Google Scholar]
- Sugiura, K.; Oi, T.; Tanaka, T.; Hamagashira, A.; Ouk, R.; Nakamura, M.; Ide, Y.; Tsuda, K.; Ito, A.; Yamauchi, A. Resistance factors of pecky rice incidence caused by the rice stink bugs (Leptocorisa chinensis, Nezara viridula) in rice line CRR-99-95W. Plant Prod. Sci. 2022, 25, 172–182. [Google Scholar] [CrossRef]
- Antunes, C.; Mendes, R.; Lima, A.; Barros, G.; Fields, P.; Da Costa, L.B.; Rodrigues, J.C.; Silva, M.J.; Correia, A.M.; Carvalho, M.O. Resistance of rice varieties to the stored product insect, Sitophilus zeamais (Coleoptera: Curculionidae). J. Econ. Entomol. 2016, 109, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Costa, D.C.D.; Almeida, A.C.D.; Araujo, M.D.; Heinrichs, E.A.; Lacerda, M.C.; Barrigossi, J.A.F.; de Jesus, F.G. Resistance of rice varieties to Sitophilus oryzae (Coleoptera: Curculionidae). Fla. Entomol. 2016, 99, 769–773. [Google Scholar] [CrossRef] [Green Version]
- Naseri, B.; Majd-Marani, S. Assessment of eight rice cultivars flour for feeding resistance to Tribolium castaneum (Herbst) (Coleoptera: Tenebrionidae). J. Stored Prod. Res. 2020, 88, 101650. [Google Scholar] [CrossRef]
- Hori, M.; Naito, S. Feeding behaviors of rice-ear bugs, Trigonotylus caelestialium and Stenotus rubrovittatus (Hemiptera: Miridae), in response to starch and its related substances. Appl. Entomol. Zool. 2018, 53, 143–150. [Google Scholar] [CrossRef]
- Chippendale, G.M. Dietary carbohydrates: Role in survival of the adult rice weevil, Sitophilus oryzae. J. Insect Physiol. 1972, 18, 949–957. [Google Scholar] [CrossRef]
- Wenefrida, I.; Utomo, H.S.; Linscombe, S.D. Development and registration of ‘Frontière’, a high protein rice cultivar. J. Plant Regist. 2017, 11, 240–244. [Google Scholar] [CrossRef]
- Linscombe, S.D.; Sha, X.; Bearb, K.; Chu, Q.R. Registration of ‘Cheniere’ rice. Crop. Sci. 2006, 46, 1814. [Google Scholar] [CrossRef]
- Oard, J.H.; Harrell, D.L.; Groth, D.E.; Bearb, K.F.; White III, L.M.; Linscombe, S.D. Registration of ‘Mermentau’ rice. J. Plant Regist. 2014, 8, 135–138. [Google Scholar] [CrossRef]
- Silva, J.P.G.F.; Baldin, E.L.L.; Canassa, V.F.; Souza, E.S.; Lourencao, A.L. Assessing antixenosis of soybean entries against Piezodorus guildinii (Hemiptera: Pentatomidae). Arthropod Plant Interact. 2014, 8, 349–359. [Google Scholar] [CrossRef]
- Panizzi, A.R.; Silva, F.A.C. Insetos sugadores de sementes (Heteroptera). In Bioecologia e Nutricao de Insetos: Base Para o Manejo de Praga; Panizzi, A.R., Parra, J.R.P., Eds.; Embrapa Informacao Tecnologica: Brasilia, Brazil, 2009; pp. 465–522. [Google Scholar]
- Harrison, S.J.; Raubenheimer, D.; Simpson, S.J.; Godin, J.-G.J.; Bertram, S.M. Towards a synthesis of frameworks in nutritional ecology: Interacting effects of protein, carbohydrate and phosphorus on field cricket fitness. Proc. R. Soc. B Biol. Sci. 2014, 281, 20140539. [Google Scholar] [CrossRef] [Green Version]
- Kökdener, M.; Kiper, F. The impact of diet protein and carbohydrate on select life-history traits of the housefly Musca domestica Linnaeus, 1758 (Diptera: Muscidae). Mun. Ent. Zool. 2020, 15, 171–179. [Google Scholar]
- Cammack, J.A.; Tomberlin, J.K. The impact of diet protein and carbohydrate on select life-history traits of the black soldier fly Hermetia illucens (L.)(Diptera: Stratiomyidae). Insects 2017, 8, 56. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Chen, M.; Lin, G.; Yang, Y.; Yu, X.; Wu, Y.; Xiong, F. Structural development and physicochemical properties of starch in caryopsis of super rice with different types of panicle. BMC Plant Biol. 2019, 19, 482. [Google Scholar] [CrossRef] [PubMed]
- Raubenheimer, D.; Simpson, S.J. Organismal stoichiometry: Quantifying non-independence among food components. Ecology 2004, 85, 1203–1216. [Google Scholar] [CrossRef]
- Chambers, P.G.; Simpson, S.J.; Raubenheimer, D. Behavioural mechanisms of nutrient balancing in Locusta migratoria nymphs. Anim. Behav. 1995, 50, 1513–1523. [Google Scholar] [CrossRef]
- de Souza, P.V.; Machado, B.R.; Zanuncio, J.C.; Araújo, M.S.; Alves, G.C.S.; de Jesus, F.G. Cultivation of resistant soybean varieties and application of silicon (Si) on biology of Euschistus heros (Hemiptera: Pentatomidae). Aust. J. Crop. Sci. 2016, 10, 1404–1409. [Google Scholar] [CrossRef]
- War, A.R.; Taggar, G.K.; Hussain, B.; Taggar, M.S.; Nair, R.M.; Sharma, H.C. Plant defence against herbivory and insect adaptations. AoB Plants 2018, 10, ply037. [Google Scholar] [CrossRef]
- Bowling, C.C. Stylet sheath as an indicator of feeding-activity of the rice stink bug (Heteroptera, Pentatomidae). J. Econ. Entomol. 1979, 72, 259–260. [Google Scholar] [CrossRef]
- Blackman, B.D.; Stout, M. Development of rice stink bug, Oebalus pugnax F., nymphs on rice kernels and effects of nymphal feeding on rice yields. Southwest. Entomol. 2017, 42, 641–650. [Google Scholar] [CrossRef]
- Espino, L.; Way, M.O.; Olson, J.K. Most susceptible stage of rice panicle development to Oebalus pugnax (Hemiptera: Pentatomidae). J. Econ. Entomol. 2007, 100, 1282–1290. [Google Scholar] [CrossRef]
- Moldenhauer, K.; Counce, P.; Hardke, J. Rice growth and development.; University of Arkansas Division of Agriculture, Cooperative Extension Service: Fayetteville, AR, USA, 2018; pp. 9–20. [Google Scholar]
- Cato, A.J.; Lorenz, G.M.; Bateman, N.R.; Hardke, J.T.; Black, J.L.; Thrash, B.C.; Johnson, D.L.; Gore, J.; Studebaker, G.; Fan, S.X. Susceptibility of rice to Oebalus pugnax (F.)(Hemiptera: Pentatomidae) feeding at different levels of grain maturity and impacts on insecticide termination. J. Econ. Entomol. 2020, 113, 249–254. [Google Scholar] [CrossRef]
- Ho, H.-Y.; Millar, J.G. Compounds in metathoracic glands of adults and dorsal abdominal glands of nymphs of the stink bugs, Chlorochroa uhleri, C. sayi, and C. ligata (Hemiptera: Pentatomidae). Zool. Stud. 2001, 40, 193–198. [Google Scholar]
- Fávaro, C.F.; Santos, T.B.; Zarbin, P.H.G. Defensive compounds and male-produced sex pheromone of the stink bug, Agroecus griseus. J. Chem. Ecol. 2012, 38, 1124–1132. [Google Scholar] [CrossRef]
- Noge, K.; Prudic, K.L.; Becerra, J.X. Defensive roles of (E)-2-Alkenals and related compounds in Heteroptera. J. Chem. Ecol. 2012, 38, 1050–1056. [Google Scholar] [CrossRef] [PubMed]
- Eliyahu, D.; Ceballos, R.A.; Saeidi, V.; Becerra, J.X. Synergy versus potency in the defensive secretions from nymphs of two Pentatomomorphan families (Hemiptera: Coreidae and Pentatomidae). J. Chem. Ecol. 2012, 38, 1358–1365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noge, K.; Becerra, J.X. 4-Oxo-(E)-2-hexenal produced by Heteroptera induces permanent locomotive impairment in crickets that correlates with free thiol depletion. FEBS Open Bio 2015, 5, 319–324. [Google Scholar] [CrossRef]
- Prudic, K.L.; Noge, K.; Becerra, J.X. Adults and nymphs do not smell the same: The different defensive compounds of the giant mesquite bug (Thasus neocalifornicus: Coreidae). J. Chem. Ecol. 2008, 34, 734–741. [Google Scholar] [CrossRef]
- Nilakhe, S.S. Host Plant Resistance and Ecological Studies on the Rice Stink Bug Oebalus pugnax (Fabricius); Louisiana State University: Baton Rouge, LA, USA, 1974. [Google Scholar]
- Zhong, Y.Z.; Zhang, J.P.; Ren, L.L.; Tang, R.; Zhan, H.X.; Chen, G.H.; Zhang, F. Behavioral responses of the egg parasitoid Trissolcus japonicus to volatiles from adults of its stink bug host, Halyomorpha halys. J. Pest Sci. 2017, 90, 1097–1105. [Google Scholar] [CrossRef]
- Laumann, R.A.; Aquino, M.F.S.; Moraes, M.C.B.; Pareja, M.; Borges, M. Response of the egg parasitoids Trissolcus basalis and Telenomus podisi to compounds from defensive secretions of stink bugs. J. Chem. Ecol. 2009, 35, 8–19. [Google Scholar] [CrossRef]
- McPherson, J.E. Invasive Stink Bugs and Related Species (Pentatomoidea): Biology, Higher Systematics, Semiochemistry, and Management; CRC Press: Boca Raton, FL, USA, 2018. [Google Scholar]
- Lockwood, J.A.; Story, R.N. Adaptive functions of nymphal aggregation in the southern green stink bug, Nezara viridula (L.)(Hemiptera: Pentatomidae). Environ. Entomol. 1986, 15, 739–749. [Google Scholar] [CrossRef]
- Lockwood, J.A.; Story, R.N. Bifunctional pheromone in the first instar of the southern green stink bug, Nezara viridula (L.)(Hemiptera: Pentatornidae): Its characterization and interaction with other stimuli. Ann. Entomol. Soc. Am. 1985, 78, 474–479. [Google Scholar] [CrossRef]
- Hamm, J.C. Ecology and Chemical Ecology of Plant-Insect Interactions in Rice—Implications for Pests Control; Louisiana State University: Baton Rouge, LA, USA, 2011. [Google Scholar]
- Gunawardena, N.E.; Herath, H.M.W.K.B. Significance of medium chain n-alkanes as accompanying compounds in hemipteran defensive secretions: An investigation based on the defensive secretion of Coridius janus. J. Chem. Ecol. 1991, 17, 2449–2458. [Google Scholar] [CrossRef]
- Shin, E.; Kim, J.; Park, C.G. Identification and laboratory behavioral test of the alarm pheromone of Acanthocoris sordidus Thunberg (Hemiptera: Coreidae). Entomol. Res. 2017, 47, 101–106. [Google Scholar] [CrossRef]
- Fucarino, A.; Millar, J.G.; McElfresh, J.S.; Colazza, S. Chemical and physical signals mediating conspecific and heterospecific aggregation behavior of first instar stink bugs. J. Chem. Ecol. 2004, 30, 1257–1269. [Google Scholar] [CrossRef] [PubMed]
- Noge, K. Hexanal, a major volatile found in fresh peanut seed, elicits foraging behavior in the laboratory-reared brown marmorated stink bug, Halyomorpha halys (Heteroptera: Pentatomidae). J. Pestic. Sci. 2019, 44, 15–19. [Google Scholar] [CrossRef] [Green Version]
- Gregorovicova, M.; Cernikova, A. Reactions of leopard geckos (Eublepharis macularius) to defensive secretion of Graphosoma lineatum (Heteroptera Pentatomidae): An experimental approach. Ethol. Ecol. Evol. 2016, 28, 367–384. [Google Scholar] [CrossRef]
- Sosa-Gomez, D.R.; Boucias, D.G.; Nation, J.L. Attachment of Metarhizium anisopliaeto the Southern green stink bug Nezara viridula cuticle and fungistatic effect of cuticular lipids and aldehydes. J. Invertebr. Pathol. 1997, 69, 31–39. [Google Scholar] [CrossRef] [PubMed]
- da Silva, R.A.; Quintela, E.D.; Mascarin, G.M.; Pedrini, N.; Liao, L.M.; Ferri, P.H. Unveiling chemical defense in the rice stalk stink bug against the entomopathogenic fungus Metarhizium anisopliae. J. Invertebr. Pathol. 2015, 127, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Lopes, R.B.; Laumann, R.A.; Blassioli-Moraes, M.C.; Borges, M.; Faria, M. The fungistatic and fungicidal effects of volatiles from metathoracic glands of soybean-attacking stink bugs (Heteroptera: Pentatomidae) on the entomopathogen Beauveria bassiana. J. Invertebr. Pathol. 2015, 132, 77–85. [Google Scholar] [CrossRef]
- Tillman, P. Parasitism and predation of stink bug (Heteroptera: Pentatomidae) eggs in Georgia corn fields. Environ. Entomol. 2010, 39, 1184–1194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Counce, P.A.; Keisling, T.C.; Mitchell, A.J. A uniform, objective, and adaptive system for expressing rice development. Crop. Sci. 2000, 40, 436–443. [Google Scholar] [CrossRef] [Green Version]
- Van Den Dool, H.; Kratz, P.D. A generalization of the retention index system including linear temperature programmed gas-liquid partition chromatography. J. Chromatogr. A 1963, 11, 463–471. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]



{kind=link}
{kind=link}
| Rice Variety | Development Time | Survival | Male Weight | Female Weight |
|---|---|---|---|---|
| Cheniere | 16.25 ± 0.35 | 88.88 ± 5.55 | 33.17 ± 0.79 | 44.33 ± 2.04 |
| CL151 | 16.27 ± 0.18 | 91.67 ± 5.69 | 34.17 ± 0.60 | 43.60 ± 1.03 |
| Frontiere | 16.38 ± 0.26 | 91.67 ± 5.69 | 32.33 ± 0.56 | 43.33 ± 1.23 |
| Jazzman | 16.28 ± 0.32 | 77.77 ± 10.25 | 31.80 ± 1.07 | 44.67 ± 1.74 |
| Jupiter | 16.03 ± 0.22 | 83.33 ± 11.39 | 34.50 ± 1.75 | 44.33 ± 2.25 |
| Kaybonnet | 16.63 ± 0.21 | 75.02 ± 8.33 | 32.83 ± 1.05 | 43.67 ± 1.58 |
| LaGrue | 16.42 ± 0.40 | 88.88 ± 5.55 | 33.33 ± 0.76 | 44.00 ± 1.00 |
| Mermentau | 16.47 ± 0.40 | 88.87 ± 3.52 | 30.83 ± 1.25 | 39.83 ± 1.58 |
| Group | Compound | 1st Instar | 2nd Instar | 3rd Instar | 4th Instar | 5th Instar |
|---|---|---|---|---|---|---|
| Alkane | Decane * | ND | ND | 0.43 ± 0.22 | ND | ND |
| Undecane * | ND | ND | ND | 0.31 ± 0.07 | 0.19 ± 0.03 | |
| Dodecane * | 0.28 ± 0.05 | 0.28 ± 0.05 | 0.48 ± 0.19 | 0.73 ± 0.36 | 0.71 ± 0.30 | |
| Tridecane * | 26.17 ± 2.07 | 30.45 ± 1.95 | 30.83 ± 5.68 | 26.74 ± 5.41 | 22.41 ± 5.51 | |
| Tetradecane * | ND | ND | ND | ND | 0.09 ± 0.04 | |
| Pentadecane * | ND | ND | ND | ND | 0.13 ± 0.06 | |
| Alkene | 1-Tridecene † | ND | 0.11 ± 0.01 | 0.10 ± 0.06 | 0.30 ± 0.15 | 0.42 ± 0.18 |
| Aldehyde | (E)-2-Hexenal † | ND | ND | 0.24 ± 0.13 | 0.26 ± 0.12 | 0.50 ± 0.30 |
| Cinnamaldehyde † | 0.20 ± 0.08 | ND | ND | ND | ND | |
| 2-Undecenal † | 26.67 ± 1.57 | 0.65 ± 0.01 | 0.06 ± 0.03 | 0.08 ± 0.03 | ND | |
| Tridecanal † | ND | 0.09 ± 0.01 | 0.11 ± 0.07 | 0.31 ± 0.18 | ND | |
| Tetradecanal † | 8.43 ± 1.48 | 24.28 ± 1.93 | 20.99 ± 6.04 | 17.86 ± 4.72 | 22.85 ± 5.35 | |
| Oxo-alkenal | (E)-4-Oxo-2-hexenal † | 35.57 ± 0.66 | 43.30 ± 4.08 | 46.74 ± 5.87 | 53.37 ± 10.17 | 52.69 ± 11.01 |
| Terpenoids | Linalool † | 2.33 ± 0.90 | 0.77 ± 0.57 | ND | ND | ND |
| α-terpineol † | 0.42 ± 0.11 | ND | ND | ND | ND |
| Group | Compound | Retention Time (min) | Retention Index | Male | Female |
|---|---|---|---|---|---|
| Alkane | Octane * | 6.7 | 801 | 0.04 ± 0.02 | 0.08 ± 0.05 |
| Undecane * | 13.9 | 1099 | 1.41 ± 0.39 | 1.16 ± 0.57 | |
| Dodecane * | 16.1 | 1201 | 4.80 ± 0.38 | 5.16 ± 1.86 | |
| Tridecane * | 18.2 | 1304 | 37.46 ± 3.53 | 27.57 ± 4.87 | |
| Tetradecane * | 20.0 | 1399 | 0.72 ± 0.17 | 0.76 ± 0.17 | |
| Pentadecane * | 21.7 | 1499 | 1.33 ± 0.22 | 1.12 ± 0.37 | |
| Hexadecane * | 23.5 | 1601 | 0.02 ± 0.01 | 0.03 ± 0.01 | |
| Heptadecane * | 25.0 | 1699 | 0.02 ± 0.01 | 0.03 ± 0.01 | |
| Octadecane * | 26.5 | 1798 | 0.01 ± 0.00 | 0.02 ± 0.01 | |
| Alkene | 1-Tridecene † | 17.9 | 1292 | 1.32 ± 0.14 | 1.43 ± 0.32 |
| 1-Tetradecene † | 18.7 | 1330 | 0.03 ± 0.01 | 0.05 ± 0.02 | |
| 1-Pentadecene † | 21.6 | 1492 | 0.07 ± 0.03 | 0.04 ± 0.01 | |
| Alcohol | 3-Hexanol † | 6.6 | 795 | 0.04 ± 0.01 | 0.07 ± 0.02 |
| (E)-2-Octenol † | 13.2 | 1070 | 0.70 ± 0.18 | 2.51 ± 0.87 | |
| (E)-2-Decenol † | 17.5 | 1271 | 0.29 ± 0.07 | 2.29 ± 1.89 | |
| Aldehyde | (E)-2-Hexenal † | 8.1 | 853 | 4.62 ± 0.89 | 7.89 ± 1.37 |
| (Z)-2-Heptenal † | 10.6 | 959 | 0.02 ± 0.01 | 0.04 ± 0.01 | |
| (E)-2-Octenal † | 13.1 | 1062 | 13.54 ± 1.15 | 9.46 ± 2.22 | |
| (E, E)- 2,4-Octadienal † | 14.2 | 1113 | ND | 0.02 ± 0.02 | |
| (E)-2-Nonenal † | 15.3 | 1164 | 0.04 ± 0.01 | 0.05 ± 0.04 | |
| (Z)-4-Decenal † | 16.0 | 1199 | ND | 0.60 ± 0.58 | |
| (Z)-2-Decenal † | 17.4 | 1266 | 5.33 ± 0.79 | 3.84 ± 1.71 | |
| Tetradecanal † | 23.7 | 1615 | 0.04 ± 0.02 | 0.06 ± 0.02 | |
| Oxo-alkenal | (E)-4-Oxo-2-hexenal † | 10.9 | 970 | 12.07 ± 1.22 | 12.26 ± 2.57 |
| Ketone | 3-Hexanone † | 6.4 | 784 | 0.02 ± 0.00 | 0.02 ± 0.01 |
| 2-Hexanone † | 6.5 | 789 | 0.16 ± 0.04 | 0.23 ± 0.06 | |
| 6-Tridecanone | 21.4 | 0.05 ± 0.01 | 0.05 ± 0.02 | ||
| 4-Tridecanone | 21.3 | 0.11 ± 0.02 | 0.10 ± 0.04 | ||
| Esters | (Z)-2-Hexenyl acetate † | 12.0 | 1014 | 0.04 ± 0.02 | 1.90 ± 0.89 |
| (E)-2-Heptenyl acetate † | 14.3 | 1116 | ND | 0.01 ± 0.01 | |
| 1-Octenyl-3-acetate | 16.4 | 8.73 ± 1.20 | 14.60 ± 2.66 | ||
| (E)-2-Decenyl acetate † | 20.2 | 1410 | 6.97 ± 0.43 | 6.53 ± 0.78 | |
| Isopropyl palmitate † | 29.7 | 2014 | 0.01 ± 0.00 | 0.01 ± 0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhavanam, S.; Stout, M.J. Varietal Resistance and Chemical Ecology of the Rice Stink Bug, Oebalus pugnax, on Rice, Oryza sativa. Plants 2022, 11, 3169. https://doi.org/10.3390/plants11223169
Bhavanam S, Stout MJ. Varietal Resistance and Chemical Ecology of the Rice Stink Bug, Oebalus pugnax, on Rice, Oryza sativa. Plants. 2022; 11(22):3169. https://doi.org/10.3390/plants11223169
Chicago/Turabian StyleBhavanam, Santhi, and Michael J. Stout. 2022. "Varietal Resistance and Chemical Ecology of the Rice Stink Bug, Oebalus pugnax, on Rice, Oryza sativa" Plants 11, no. 22: 3169. https://doi.org/10.3390/plants11223169