

Gamma Radiation Induced In-Vitro Mutagenesis and Isolation of Mutants for Early Flowering and Phytomorphological Variations in Dendrobium ‘Emma White’
 , and
, and
Abstract
:1. Introduction
2. Results
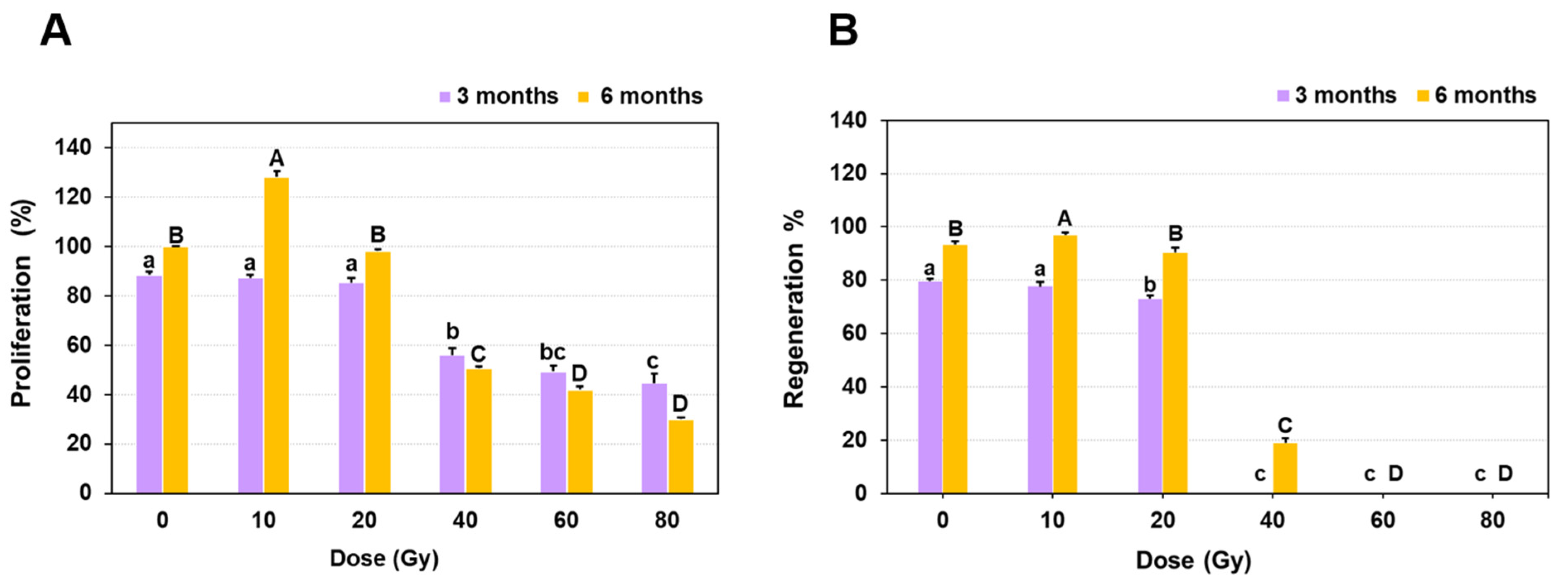
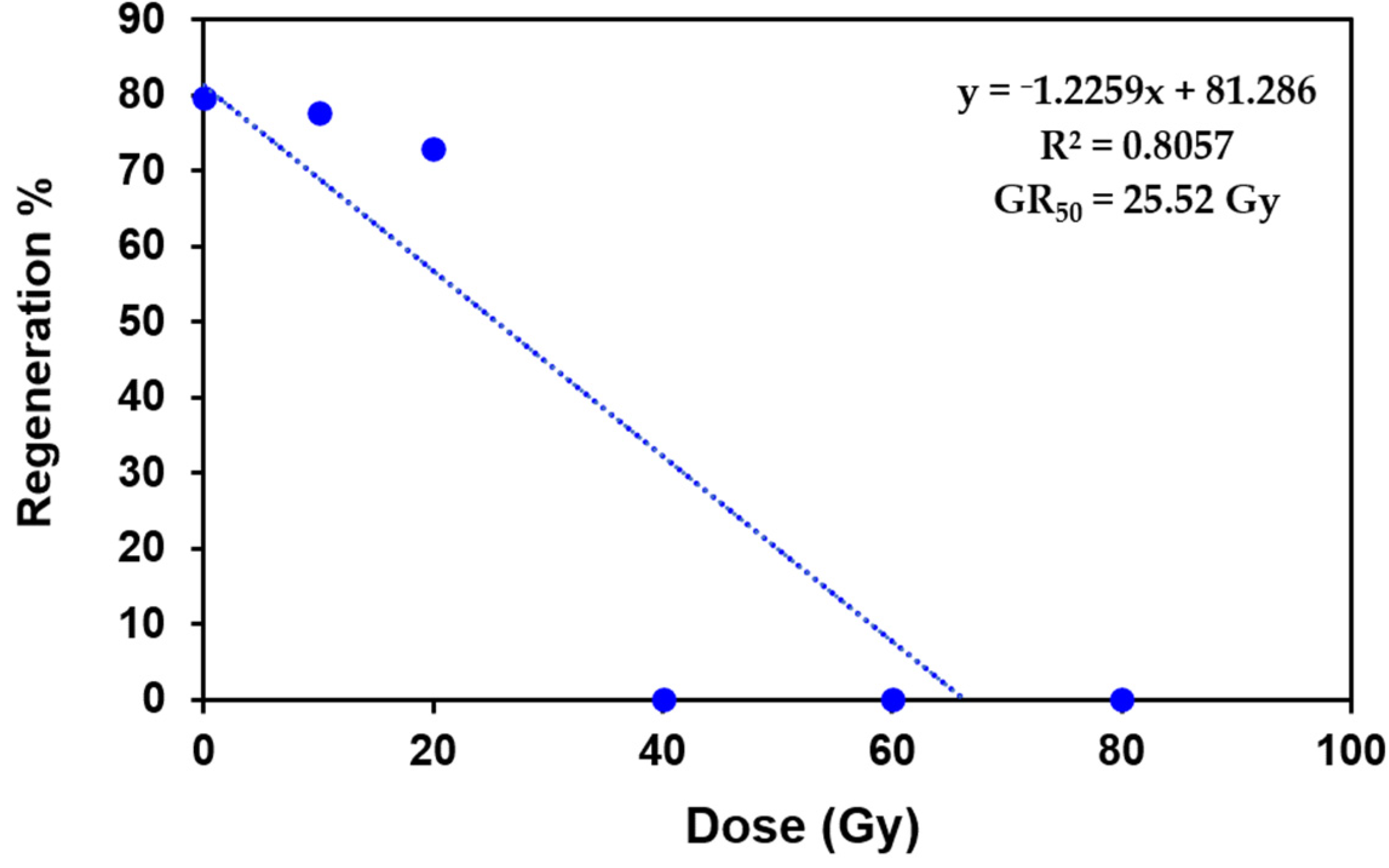
2.1. Growth Response of Protocorm Like Bodies (PLBs) to Different Doses of Gamma Radiation
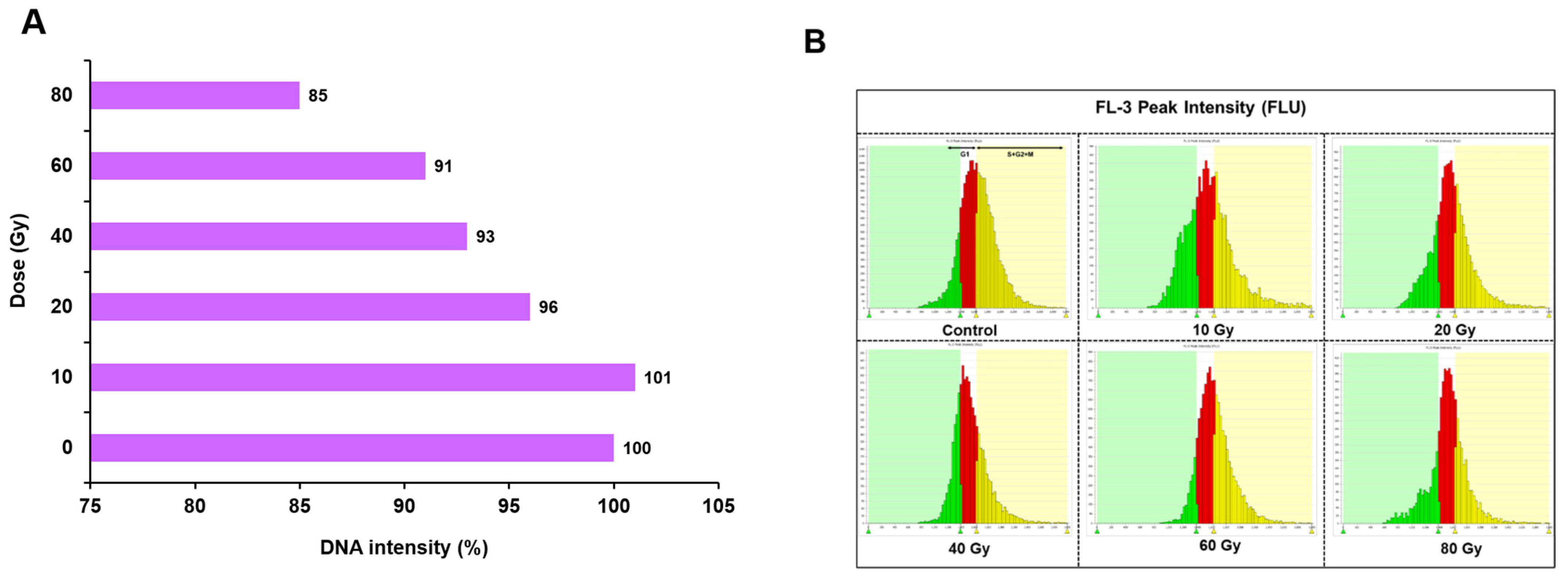
2.2. Nuclear DNA Content Estimation and Cell Cycle Analysis in Irradiated PLBs by High-Throughput Cell Analyser (HTCA)
2.3. Frequency and Spectrum of Variation among Gamma-Irradiated Population of Dendrobium ‘Emma White’
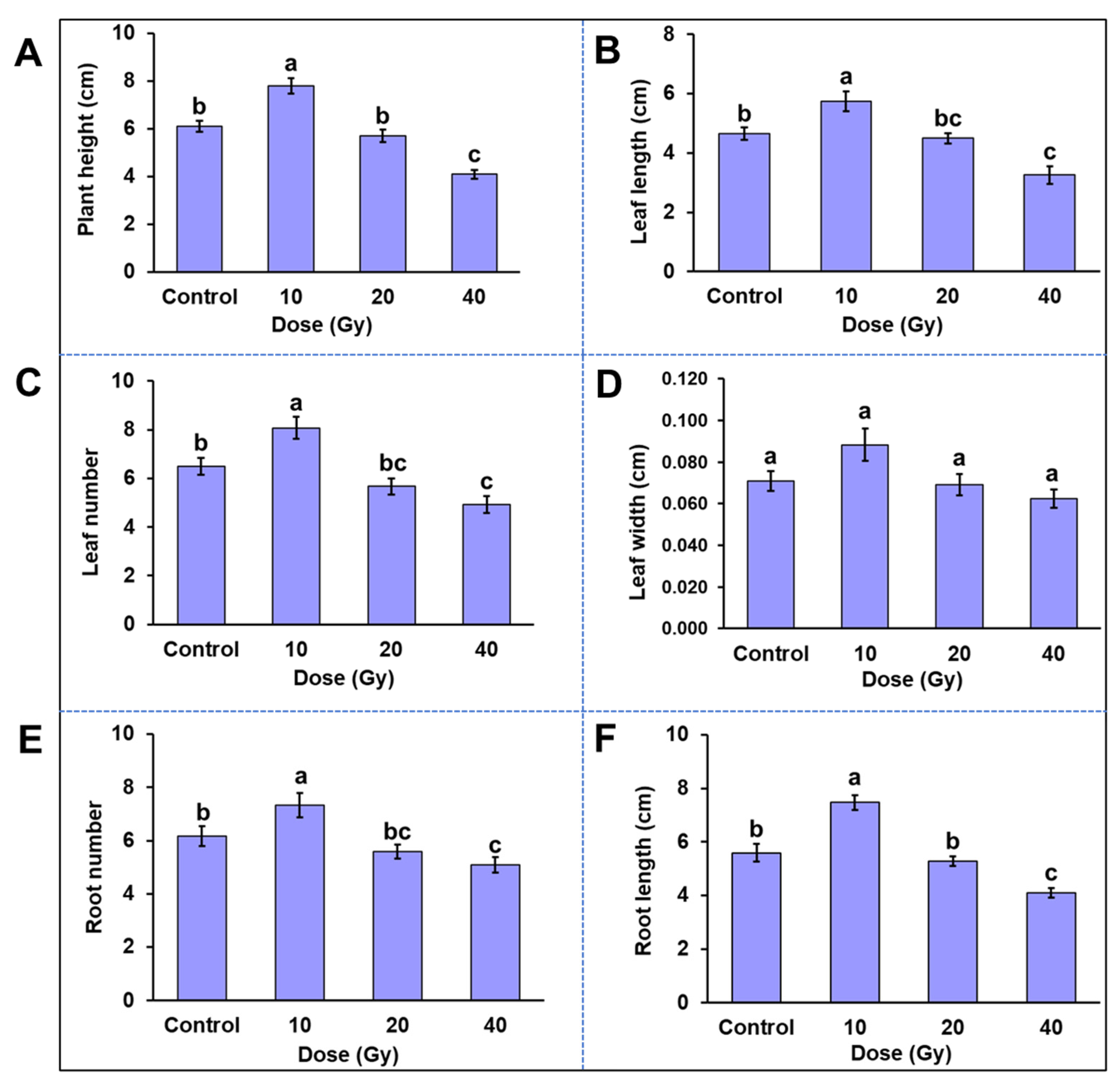
2.4. Phenotypic Variation among In Vitro Plantlets Derived from Gamma-Irradiated PLBs
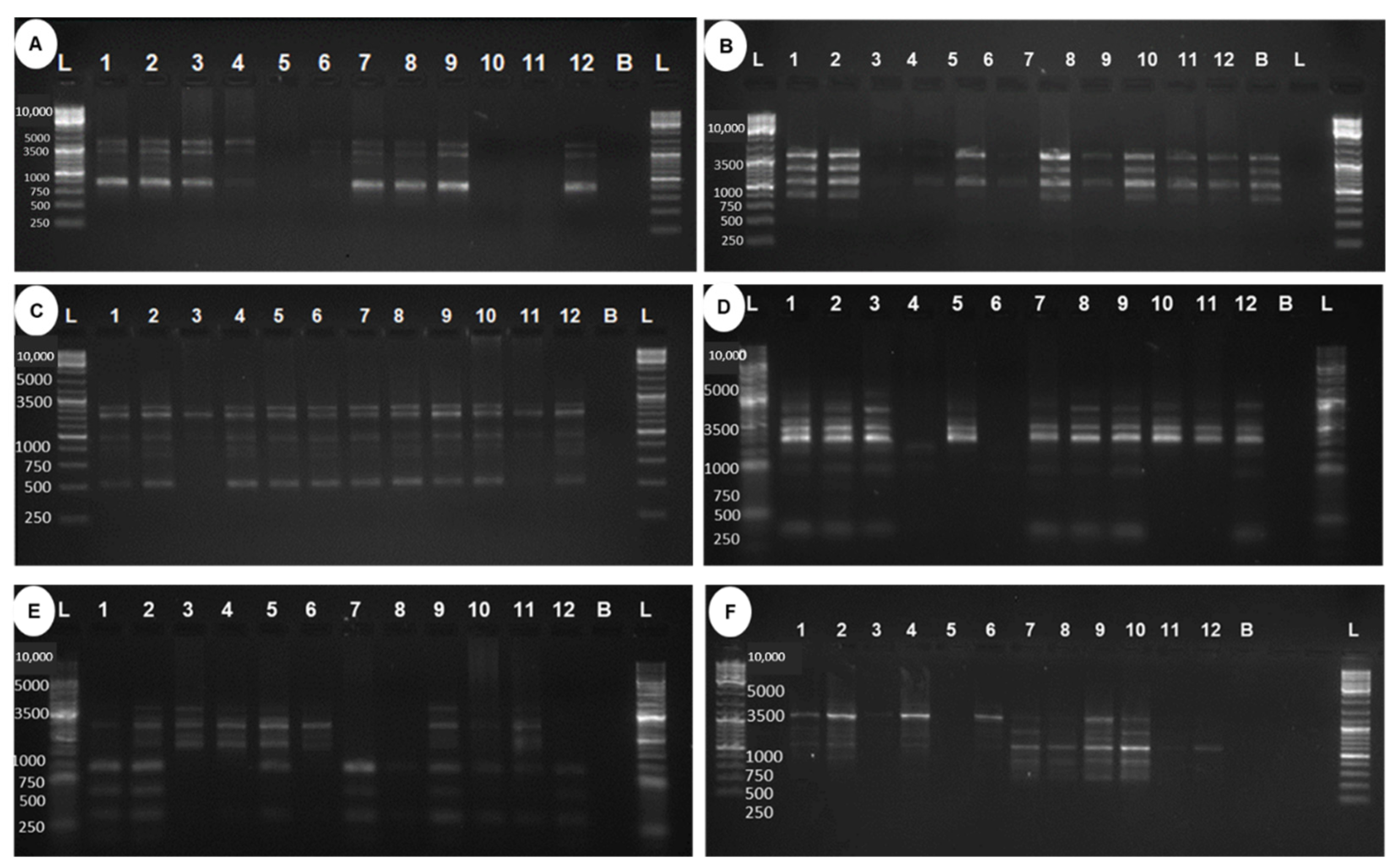
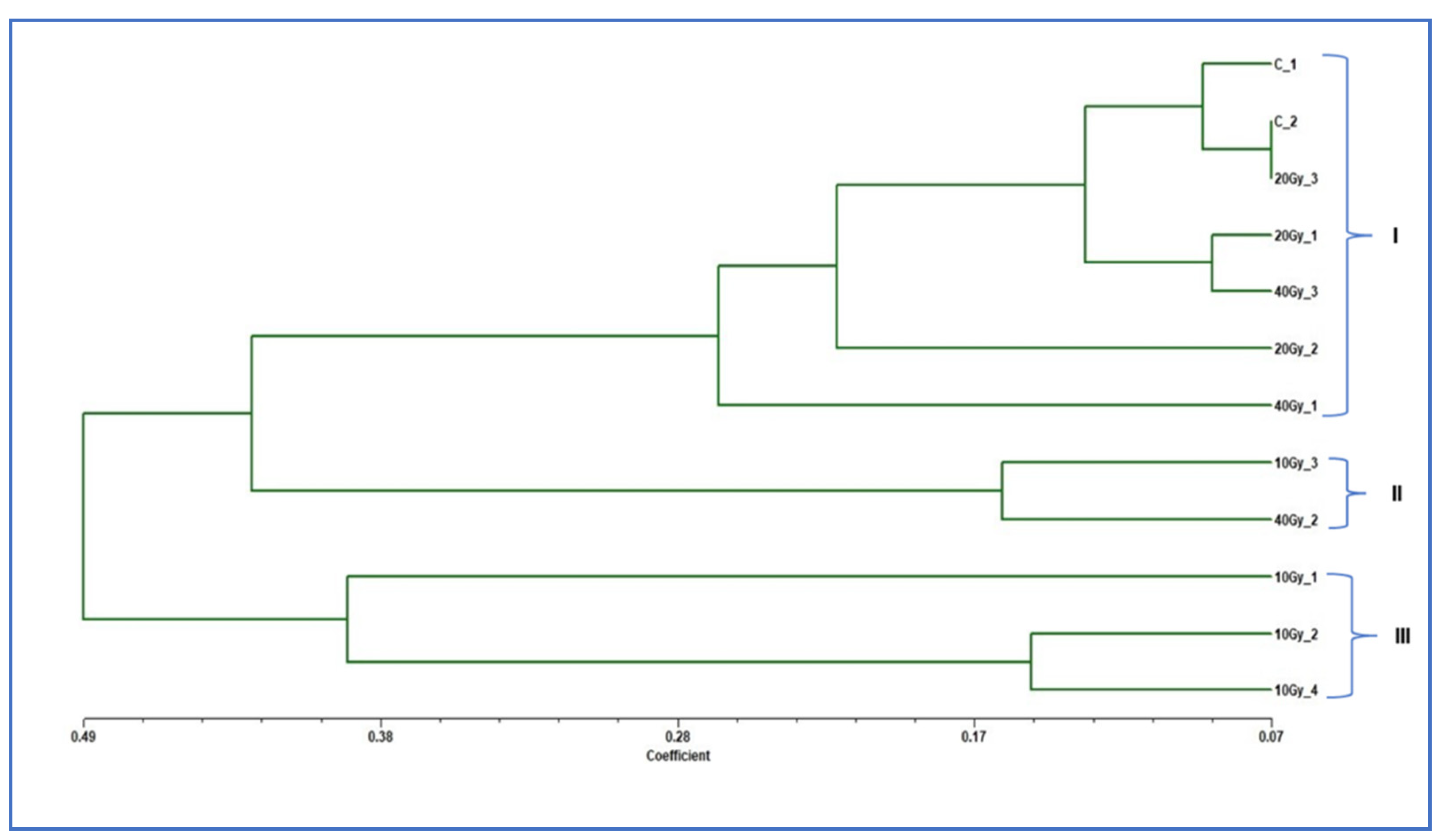
2.5. Genetic Analysis of Regenerated Mutants Based on Random Amplified Polymorphic DNA Markers
2.6. Leaf Variation among 10 Gy Mutant Population
2.7. Isolation of Early Flowering Mutant among 10 Gy Mutant Population
3. Discussion
4. Materials and Methods
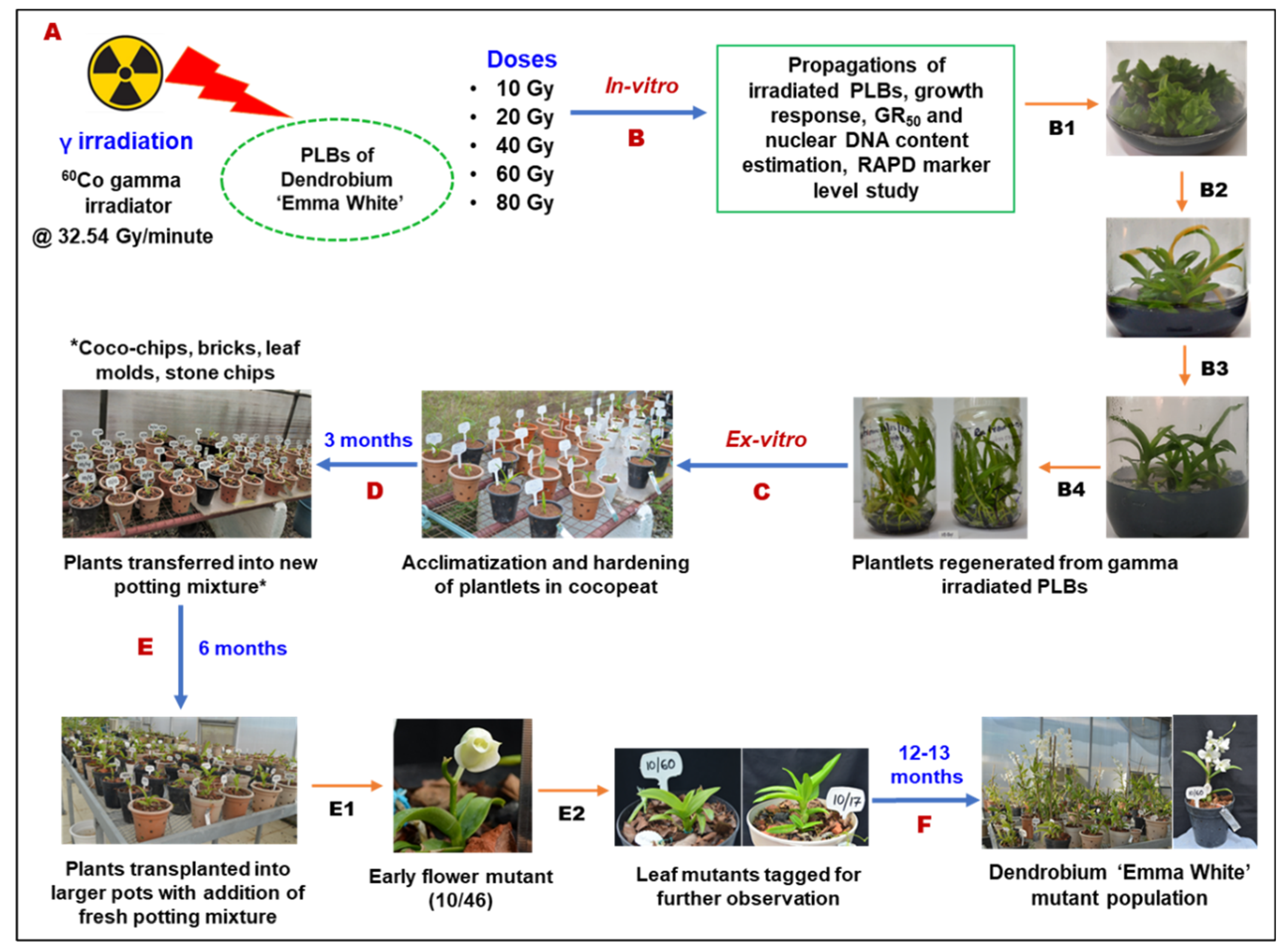
4.1. Plant Material for Gamma Irradiation Treatment
4.2. Evaluation of Growth Response of Irradiated PLBs
4.3. High-Throughput Cell Analysis and Estimation of Nuclear DNA Content of PLBs
4.4. Random Amplified Polymorphic DNA (RAPD)-Based Divergence Analysis of In Vitro Gamma Mutants
4.5. Morphological Variation Analysis of Putative Mutants at In Vitro and Ex Vitro Stage
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cakova, V.; Bonte, F.; Lobstein, A. Dendrobium: Sources of Active Ingredients to Treat Age-Related Pathologies. Aging Dis. 2017, 8, 827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moudi, M.; Go, R.; Yong, C.Y.S.; Saleh, M.N. A Review on Molecular Systematic of the Genus Dendrobium Sw. Acta Biol. Malays. 2013, 2, 71–78. [Google Scholar] [CrossRef]
- Hinsley, A.; de Boer, H.J.; Fay, M.F.; Gale, S.W.; Gardiner, L.M.; Gunasekara, R.S.; Kumar, P.; Masters, S.; Metusala, D.; Roberts, D.L.; et al. A Review of the Trade in Orchids and Its Implications for Conservation. Bot. J. Linn. Soc. 2018, 186, 435–455. [Google Scholar] [CrossRef]
- Thammasiri, K. Current Status of Orchid Production in Thailand. Acta Hortic. 2015, 1078, 25–33. [Google Scholar] [CrossRef]
- Baker, M.L.; Baker, C.O. Orchid Species Culture: Dendrobium; Timber Press: Portland, OR, USA, 1996. [Google Scholar]
- Wanitprapha, K.; Kevin, M.; Yokoyama, K.M. Dendrobium Economic Fact Sheet. Available online: https://scholarspace.manoa.hawaii.edu/bitstream/10125/54681/CtahrpsEconFact13Dendro.pdf (accessed on 30 October 2022).
- Nor Hazlina, M.S.; Wahba, L.E.; Fadelah, A.; Wickneswari, R. Genetic Relationships among 81 Dendrobium Accessions from Malaysia. Malays. Appl. Biol. 2013, 42, 35–40. [Google Scholar]
- Devadas, R.; Pattanayak, S.L.; Singh, D.R. Studies on Cross Compatibility in Dendrobium Species and Hybrids. Indian J. Genet. Plant Breed. 2016, 76, 344. [Google Scholar] [CrossRef]
- Devadas, R.; Khatiwara, P.; Barman, D.; Das, S.P. Breeding Dendrobium Phalaenopsis-Cane Type Hybrid in India: NRCO- 42 (Den. “Emma White” × Den. “Pompadour’’). Indian J. Genet. Plant Breed. 2009, 69, 237–242. [Google Scholar]
- Wilfret, G.J.; Kamemoto, H. Genome and Karyotype Relationships in the Genus Dendrobium (Orchidaceae). I. Crossability. Am. J. Bot. 1969, 56, 521–526. [Google Scholar] [CrossRef]
- Zhang, S.; Yang, Y.; Li, J.; Qin, J.; Zhang, W.; Huang, W.; Hu, H. Physiological Diversity of Orchids. Plant Divers. 2018, 40, 196–208. [Google Scholar] [CrossRef]
- Teixeira da Silva, J.A.; Jin, X.; Dobránszki, J.; Lu, J.; Wang, H.; Zotz, G.; Cardoso, J.C.; Zeng, S. Advances in Dendrobium Molecular Research: Applications in Genetic Variation, Identification and Breeding. Mol. Phylogenet. Evol. 2016, 95, 196–216. [Google Scholar] [CrossRef]
- Adams, P.B. Systematics of Dendrobiinae (Orchidaceae), with Special Reference to Australian Taxa. Bot. J. Linn. Soc. 2011, 166, 105–126. [Google Scholar] [CrossRef] [Green Version]
- Luyt, R.; Johnson, S.D. Hawkmoth Pollination of the African Epiphytic Orchid Mystacidium Venosum, with Special Reference to Flower and Pollen Longevity. Plant Syst. Evol. 2001, 228, 49–62. [Google Scholar] [CrossRef]
- Teoh, S.B. A Breeding Program for the Development of Seed Propagated Uniform Hybrid Allopolyploid Aranda Cultivars for Cut Flowers. Malay. Orchid Rev. Singap. 1986, 20, 42–45. [Google Scholar]
- Suprasanna, P.; Mirajkar, S.J.; Bhagwat, S.G. Induced Mutations and Crop Improvement. In Plant Biology and Biotechnology; Bahadur, B., Venkat Rajam, M., Sahijram, L., Krishnamurthy, K.V., Eds.; Springer: New Delhi, India, 2015; pp. 593–617. [Google Scholar] [CrossRef]
- International Atomic Energy Agency Home Page. Available online: Https://Nucleus.Iaea.Org/Sites/Mvd/SitePages/Home.Aspx (accessed on 10 May 2021).
- Thammasiri, K. Effects of gamma irradiation on protocorm-like bodies of Cattleya alliances. In Proceedings of the 6th Nuclear Science and Technology Conference, Bangkok, Thailand, 2–4 December 1996; p. 761. [Google Scholar]
- Ariffin, S.; Mohamad, A.; Hassan, A.A.; Ahmad, Z.; Basiran, M.N. Flower Morphology of Dendrobium Sonia Mutants. In Proceedings of the Research and Development Seminar, Bangi, Malaysia, 12–15 October 2010; p. 4. [Google Scholar]
- Álvarez-Holguín, A.; Morales-Nieto, C.R.; Avendaño-Arrazate, C.H.; Corrales-Lerma, R.; Villarreal-Guerrero, F.; Santellano-Estrada, E.; Gómez-Simuta, Y. Mean lethal dose (LD50) and growth reduction (GR50) due to gamma radiation in Wilman Lovegrass (Eragrostis Superba). Rev. Mex. Cienc. Pecu. 2019, 10, 227–238. [Google Scholar] [CrossRef]
- Sokal, R.R.; Rohlf, F.J. The Comparison of Dendrograms by Objective Methods. Taxon 1962, 11, 33–40. [Google Scholar] [CrossRef]
- Luan, L.Q.; Uyen, N.H.P.; Ha, V.T.T. In Vitro Mutation Breeding of Paphiopedilum by Ionization Radiation. Sci. Hortic. 2012, 144, 1–9. [Google Scholar] [CrossRef]
- Xo, D.H.; Luan, L.Q. In Vitro Mutagenesis of Cymbidium La Bell “Anna Belle” by γ-Rays Irradiation and Oligochitosan Interaction. Int. J. Environ. Agric. Biotechnol. 2017, 2, 1540–1550. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, H.; Shimizu, A.; Hase, Y.; Tanaka, A.; Shikazono, N.; Degi, K.; Morishita, T. Effects of Ion Beam Irradiation on Mutation Induction and Nuclear DNA Content in Chrysanthemum. Breed. Sci. 2010, 60, 398–404. [Google Scholar] [CrossRef] [Green Version]
- Wi, S.G.; Chung, B.Y.; Kim, J.-H.; Baek, M.-H.; Yang, D.H.; Lee, J.-W.; Kim, J.-S. Ultrastructural Changes of Cell Organelles in Arabidopsis Stems after Gamma Irradiation. J. Plant Biol. 2005, 48, 195–200. [Google Scholar] [CrossRef]
- Kim, J.-H.; Chung, B.Y.; Kim, J.-S.; Wi, S.G. Effects of Planta Gamma-Irradiation on Growth, Photosynthesis, and Antioxidative Capacity of Red Pepper (Capsicum Annuum L.) Plants. J. Plant Biol. 2005, 48, 47–56. [Google Scholar] [CrossRef]
- Minisi, F.; El-Mahrouk, M.; Rida, M.F.; Nasr, M. Effects of Gamma Radiation on Germination, Growth Characteristics and Morphological Variations of Moluccella Laevis L. Am. Eurasian J. Agric. Environ. Sci. 2013, 13, 696–704. [Google Scholar] [CrossRef]
- Fathin, T.S.; Hartati, S.; Yunus, A. Diversity Induction with Gamma Ray Irradiation on Dendrobium Odoardi Orchid. IOP Conf. Ser. Earth Environ. Sci. 2021, 637, 012035. [Google Scholar] [CrossRef]
- Pathirana, R. Plant Mutation Breeding in Agriculture. CABI Rev. 2011, 2011, 1–20. [Google Scholar] [CrossRef]
- Kozlowska-Kalisz, J. The Influence of Ionizing Radiation on Biological Activity of Endogenous Growth Regulators in Orchids/Cymbidium in Tissue Culture. Acta Hortic. 1979, 91, 261–268. [Google Scholar] [CrossRef]
- Kim, S.H.; Kim, S.W.; Ahn, J.-W.; Ryu, J.; Kwon, S.-J.; Kang, B.-C.; Kim, J.-B. Frequency, Spectrum, and Stability of Leaf Mutants Induced by Diverse γ-Ray Treatments in Two Cymbidium Hybrids. Plants 2020, 9, 546. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.M.; Lee, H.J.; Kim, Y.S.; Kang, S.Y.; Kim, D.S.; Kim, J.B.; Ahn, J.W.; Ha, B.K.; Kim, S.H. Evaluation of the Sensitivity to Ionising γ -Radiation of a Cymbidium Hybrid. J. Hortic. Sci. Biotechnol. 2016, 91, 109–116. [Google Scholar] [CrossRef]
- Bajaj, Y.P.S. Effect of Gamma-Irradiation on Growth, RNA, Protein, and Nitrogen Contents of Bean Callus Cultures. Ann. Bot. 1970, 34, 1089–1096. [Google Scholar] [CrossRef]
- Liu, N.; Lin, Z.; Guan, L.; Gaughan, G.; Lin, G. Antioxidant Enzymes Regulate Reactive Oxygen Species during Pod Elongation in Pisum Sativum and Brassica Chinensis. PLoS ONE 2014, 9, e87588. [Google Scholar] [CrossRef] [Green Version]
- Bajpay, A.; Dwivedi, D. Gamma Ray Induced Foliage Variegation and Anatomical Aberrations in Chrysanthemum (Dendranthema Grandiflora T.) Cv. Maghi. J. Pharm. Phytochem. 2019, 8, 871–874. [Google Scholar]
- Dehgahi, R.; Joniyasa, A. Gamma Irradiation-Induced Variation in Dendrobium Sonia-28 Orchid Protocorm-Like Bodies (PLBs). Fungal Genom. Biol. 2017, 7, 151. [Google Scholar] [CrossRef]
- Billore, V.; Mirajkar, S.J.; Suprasanna, P.; Jain, M. Gamma Irradiation Induced Effects on in Vitro Shoot Cultures and Influence of Monochromatic Light Regimes on Irradiated Shoot Cultures of Dendrobium Sonia Orchid. Biotechnol. Rep. 2019, 22, e00343. [Google Scholar] [CrossRef]
- Abdullah, T.L.; Endan, J.; Nazir, B.M. Changes in Flower Development, Chlorophyll Mutation and Alteration in Plant Morphology of Curcuma Alismatifolia by Gamma Irradiation. Am. J. Appl. Sci. 2009, 6, 1436–1439. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.-L.; Yuan, S.-C.; Chang, K.-W.; Chen, F.-C. Gamma Irradiation Mutagenesis in Monstera Deliciosa. Acta Hortic. 2017, 1167, 213–216. [Google Scholar] [CrossRef]
- Hosoguchi, T.; Uchiyama, Y.; Komazawa, H.; Yahata, M.; Shimokawa, T.; Tominaga, A. Effect of Three Types of Ion Beam Irradiation on Gerbera (Gerbera Hybrida) In Vitro Shoots with Mutagenesis Efficiency. Plants 2021, 10, 1480. [Google Scholar] [CrossRef]
- Wylie, P.G.; Onley, D.J.; Hammerstein, A.F.; Bowen, W.P. Advances in Laser Scanning Imaging Cytometry for High-Content Screening. ASSAY Drug Dev. Technol. 2015, 13, 66–78. [Google Scholar] [CrossRef]
- Yamaguchi, H.; Shimizu, A.; Degi, K.; Morishita, T. Effects of Dose and Dose Rate of Gamma Ray Irradiation on Mutation Induction and Nuclear DNA Content in Chrysanthemum. Breed. Sci. 2008, 58, 331–335. [Google Scholar] [CrossRef] [Green Version]
- Oražem, P.; Štajner, N.; Bohanec, B. Effect of X-Ray Irradiation on Olive Shoot Culture Evaluated by Morphological Measurements, Nuclear DNA Content and SSR and AFLP Markers. Trees 2013, 27, 1587–1595. [Google Scholar] [CrossRef]
- Preuss, S.B.; Britt, A.B. A DNA-Damage-Induced Cell Cycle Checkpoint in Arabidopsis. Genetics 2003, 164, 323–334. [Google Scholar] [CrossRef]
- Madhusudan, S.; Wilson, D.M., III. Radiation Induced DNA Damage, Repair and Therapeutics. In DNA Repair and Cancer; CRC Press: Boca Raton, FL, USA, 2013. [Google Scholar] [CrossRef]
- Abu Hassan, A.; Ariffin, S.; Ahmad, Z.; Basiran, M.N.; Oono, Y.; Hase, Y.; Shikazono, N.; Narumi, I.; Tanaka, A. Mutation Induction of Orchid Plants by Ion Beams; Japan Atomic Energy Agency–Review: Ibaraki, Japan, 2016. [Google Scholar] [CrossRef]
- Brunner, H. Radiation Induced Mutations for Plant Selection. Appl. Radiat. Isot. 1995, 46, 589–594. [Google Scholar] [CrossRef]
- Li, C.; Dong, N.; Zhao, Y.; Wu, S.; Liu, Z.; Zhai, J. A Review for the Breeding of Orchids: Current Achievements and Prospects. Hortic. Plant J. 2021, 7, 380–392. [Google Scholar] [CrossRef]
- Su, J.; Jiang, J.; Zhang, F.; Liu, Y.; Ding, L.; Chen, S.; Chen, F. Current Achievements and Future Prospects in the Genetic Breeding of Chrysanthemum: A Review. Hortic. Res. 2019, 6, 109. [Google Scholar] [CrossRef]
- Dhakshanamoorthy, D.; Selvaraj, R.; Chidambaram, A. Utility of RAPD Marker for Genetic Diversity Analysis in Gamma Rays and Ethyl Methane Sulphonate (EMS)-Treated Jatropha Curcas Plants. Comptes Rendus Biol. 2015, 338, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Ong, T. Detection of Genomic Instability in Lung Cancer Tissues by Random Amplified Polymorphic DNA Analysis. Carcinogenesis 1998, 19, 233–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khosravi, A.R.; Kadir, M.A.; Kadzemin, S.B.; Zaman, F.Q.; De Silva, A.E. RAPD Analysis of Colchicine Induced Variation of the Dendrobium Serdang Beauty. Afr. J. Biotechnol. 2009, 8, 1455–1465. [Google Scholar]
- Mallón, R.; Rodríguez-Oubiña, J.; González, M.L. In Vitro Propagation of the Endangered Plant Centaurea Ultreiae: Assessment of Genetic Stability by Cytological Studies, Flow Cytometry and RAPD Analysis. Plant Cell Tissue Organ Cult. 2010, 101, 31–39. [Google Scholar] [CrossRef]
- Srivastava, D.; Mc, G.; Sarangi, S. In Vitro Mutagenesis and Characterization of Mutants through Morphological and Genetic Analysis in Orchid Aerides Crispa Lindl. Indian J. Exp. Biol. 2018, 56, 385–394. [Google Scholar]
- Hartati, S.; Muliawati, E.S. Short Communication: Genetic Variation of Coelogyne pandurate, C. rumphii and Their Hybrids Based on RAPD Markers. Biodiversitas J. Biol. Divers. 2020, 21, 4709–4713. [Google Scholar] [CrossRef]
- Tantasawat, T.P.; Khairum, A.; Tharapreuksapong, A.; Poolsawat, O.; Tantasawat, P.A. Molecular Characterization of Dendrobium ‘Earsakul’ Mutants from in Vitro Selection for Black Rot Resistance. J. Appl. Hortic. 2017, 19, 130–134. [Google Scholar] [CrossRef]
- Feldman, A.B.; Leung, H.; Baraoidan, M.; Elmido-Mabilangan, A.; Canicosa, I.; Quick, W.P.; Sheehy, J.; Murchie, E.H. Increasing Leaf Vein Density via Mutagenesis in Rice Results in an Enhanced Rate of Photosynthesis, Smaller Cell Sizes and Can Reduce Interveinal Mesophyll Cell Number. Front. Plant Sci. 2017, 8, 1883. [Google Scholar] [CrossRef] [Green Version]
- Kull, T. Cypripedium Calceolus L. J. Ecol. 1999, 87, 913–924. [Google Scholar] [CrossRef]
- Schmidt, G.; Zotz, G. Inherently Slow Growth in Two Caribbean Epiphytic Species: A Demographic Approach. J. Veg. Sci. 2002, 13, 527–534. [Google Scholar] [CrossRef]
- Hsiao, Y.-Y.; Pan, Z.-J.; Hsu, C.-C.; Yang, Y.-P.; Hsu, Y.-C.; Chuang, Y.-C.; Shih, H.-H.; Chen, W.-H.; Tsai, W.-C.; Chen, H.-H. Research on Orchid Biology and Biotechnology. Plant Cell Physiol. 2011, 52, 1467–1486. [Google Scholar] [CrossRef] [Green Version]
- Teixeira da Silva, J.A.; Hossain, M.M.; Sharma, M.; Dobránszki, J.; Cardoso, J.C.; Zeng, S. Acclimatization of in Vitro -Derived Dendrobium. Hortic. Plant J. 2017, 3, 110–124. [Google Scholar] [CrossRef]
- Hee, K.H.; Loh, C.S.; Yeoh, H.H. Early in Vitro Flowering and Seed Production in Culture in Dendrobium Chao Praya Smile (Orchidaceae). Plant Cell Rep. 2007, 26, 2055–2062. [Google Scholar] [CrossRef]
- Alvarez-Buylla, E.R.; Pelaz, S.; Liljegren, S.J.; Gold, S.E.; Burgeff, C.; Ditta, G.S.; Ribas de Pouplana, L.; Martínez-Castilla, L.; Yanofsky, M.F. An Ancestral MADS-Box Gene Duplication Occurred before the Divergence of Plants and Animals. Proc. Natl. Acad. Sci. USA 2000, 97, 5328–5333. [Google Scholar] [CrossRef] [Green Version]
- Patil, U.; Karale, A.; Katwate, S. Mutation Breeding in Chrysanthemum (Dendranthema Grandiflora T.). J. Pharmacogn. Phytochem. 2017, 6, 230–232. [Google Scholar]
- Ariffin, S.; Basiran, M.N. Induced Mutation of Dendrobium Orchid. In Proceedings of the Research and Development Seminar, Institute for Nuclear Technology Research, MINT, Bangi, Malaysia, 17–19 October 2000; p. 8. [Google Scholar]
- Magdalita, P.M.; Pascual, A.O.S.; Villareal, R.L. Characterization and Flowering Behavior of Eleven Philippine Native Phalaenopsis Species and Gamma Irradiation Effects on Phalaenopsis Aphrodite. Philipp. J. Sci. 2019, 149, 1–10. [Google Scholar] [CrossRef]
- Gamborg, O.L.; Murashige, T.; Thorpe, T.A.; Vasil, I.K. Plant Tissue Culture Media. In Vitro 1976, 12, 473–478. [Google Scholar] [CrossRef]
- deAlmeida, C.E.; Ochoa, R.; de Lima, M.C.; David, M.G.; Pires, E.J.; Peixoto, J.G.; Salata, C.; Bernal, M.A. A Feasibility Study of Fricke Dosimetry as an Absorbed Dose to Water Standard for 192Ir HDR Sources. PLoS ONE 2014, 9, e115155. [Google Scholar] [CrossRef] [Green Version]
- Sahoo, S.A.; Jha, Z.; Verulkar, S.B.; Srivastava, A.K.; Suprasanna, P. High-Throughput Cell Analysis Based Protocol for Ploidy Determination in Anther-Derived Rice Callus. Plant Cell Tissue Organ Cult. 2019, 137, 187–192. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. Isolation of Plant DNA from Fresh Tissue. Focus 1990, 12, 13–15. [Google Scholar]
- Goh, M.W.K.; Kumar, P.P.; Lim, S.H.; Tan, H.T.W. Random Amplified Polymorphic DNA Analysis of the Moth Orchids, Phalaenopsis (Epidendroideae: Orchidaceae). Euphytica 2005, 141, 11–22. [Google Scholar] [CrossRef]
- Inthawong, S.; Bundithya, W.; Kuanprasert, N.; Apavatjrut, P. Analysis of Intersectional Hybrids of Dendrobium by RAPD Technique. Kasetsart J. Nat. Sci. 2006, 40, 456–461. [Google Scholar]
- Niknejad, A.; Kadir, M.A.; Kadzimin, S.B.; Abdullah, N.A.P.; Sorkheh, K. Molecular Characterization and Phylogenetic Relationships among and within Species of Phalaenopsis (Epidendroideae: Orchidaceae) Based on RAPD Analysis. Afr. J. Biotechnol. 2009, 8, 5225–5240. [Google Scholar] [CrossRef]
- Abbas, B.; Dailami, M.; Listyorini, F.H.; Munarti. Genetic Variations and Relationships of Papua’s Endemic Orchids Based on RAPD Markers. Nat. Sci. 2017, 9, 377–385. [Google Scholar] [CrossRef] [Green Version]
- Zalewska, M.; Jerzy, M. Mutation Spectrum in Dendranthema Grandiflora Tzvelev After In Vivo and In Vitro Regeneration of Plants from Irradiated Leaves. Acta Hortic. 1997, 16, 615–618. [Google Scholar] [CrossRef]
- Jensen, R.J. NTSYS-Pc: Numerical Taxonomy and Multivariate Analysis System; Version 2.2; Exeter Publishing Setauke: New York, NY, USA, 2000. [Google Scholar]
- Powell, W.; Morgante, M.; Andre, C.; Hanafey, M.; Vogel, J.; Tingey, S.; Rafalski, A. The Comparison of RFLP, RAPD, AFLP and SSR (Microsatellite) Markers for Germplasm Analysis. Mol. Breed. 1996, 2, 225–238. [Google Scholar] [CrossRef]
- Nagy, S.; Poczai, P.; Cernák, I.; Gorji, A.M.; Hegedűs, G.; Taller, J. PICcalc: An Online Program to Calculate Polymorphic Information Content for Molecular Genetic Studies. Biochem. Genet. 2012, 50, 670–672. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Fresh Weight (mg) |
|---|---|
| Control | 648.6 ± 27.4 a |
| 10 Gy | 448.2 ± 16.6 b |
| 20 Gy | 294.4 ± 14.3 c |
| 40 Gy | 237.1 ± 4.9 d |
| 60 Gy | 232.0 ± 4.7 d |
| 80 Gy | 209.1 ± 5.9 d |
| Gamma Dose | Leaf Margin | Leaf Vein | Leaf Texture | Leaf Apex | Leaf Shape | Leaf Distribution | Leaf Color | Plantlets Regenerated | Total No. of Mutants | Variation Frequency (%) | Variation Spectrum | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| T/S | DM | T/R | DN | Lobed | Linear | Ovate | Distorted | LC | T/R | YLWM | |||||
| Control | - | - | - | - | - | - | - | - | - | - | - | 300 | 0 | - | - |
| 10 Gy | 4 | 4 | - | 6 | - | 2 | 2 | - | 1 | 2 | - | 300 | 21 | 7.0 | 7 |
| 20 Gy | 5 | 2 | 3 | 3 | - | - | 1 | 5 | 3 | - | - | 230 | 22 | 9.5 | 7 |
| 40 Gy | 8 | 3 | 2 | 5 | 1 | - | 1 | 3 | 2 | 1 | 5 | 60 | 31 | 51.6 | 10 |
| SI. No. | Primer | Total No. of Bands | Monomorphic | Polymorphic | Polymorphism (%) | PIC |
|---|---|---|---|---|---|---|
| 1 | OPB12 | 4 | 0 | 4 | 100 | 0.48 |
| 2 | OPA4 | 4 | 0 | 4 | 100 | 0.41 |
| 3 | OPAW13 | 6 | 4 | 2 | 33.33 | 0.15 |
| 4 | OPAW17 | 6 | 0 | 6 | 100 | 0.41 |
| 5 | OPB5 | 8 | 0 | 8 | 100 | 0.5 |
| 6 | OPF1 | 5 | 0 | 5 | 100 | 0.5 |
| 7 | Total | 33 | 4 | 29 | - | - |
| 8 | Mean | 5.50 | 0.67 | 4.83 | 88.89 | 0.41 |
| C_1 | C_2 | 10 Gy_1 | 10 Gy_2 | 10 Gy_3 | 10 Gy_4 | 20 Gy_1 | 20 Gy_2 | 20 Gy_3 | 40 Gy_1 | 40 Gy_2 | 40 Gy_3 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| C_1 | 0 | |||||||||||
| C_2 | 0.09 | 0 | ||||||||||
| 10 Gy_1 | 0.37 | 0.33 | 0 | |||||||||
| 10 Gy_2 | 0.37 | 0.33 | 0.33 | 0 | ||||||||
| 10 Gy_3 | 0.48 | 0.36 | 0.38 | 0.56 | 0 | |||||||
| 10 Gy_4 | 0.48 | 0.36 | 0.46 | 0.15 | 0.44 | 0 | ||||||
| 20 Gy_1 | 0.13 | 0.13 | 0.55 | 0.55 | 0.61 | 0.61 | 0 | |||||
| 20 Gy_2 | 0.24 | 0.27 | 0.46 | 0.38 | 0.61 | 0.43 | 0.20 | 0 | ||||
| 20 Gy_3 | 0.10 | 0.07 | 0.32 | 0.39 | 0.43 | 0.50 | 0.10 | 0.26 | 0 | |||
| 40 Gy_1 | 0.26 | 0.19 | 0.66 | 0.57 | 0.37 | 0.54 | 0.28 | 0.38 | 0.17 | 0 | ||
| 40 Gy_2 | 0.45 | 0.33 | 0.63 | 0.89 | 0.16 | 0.61 | 0.41 | 0.49 | 0.33 | 0.20 | 0 | |
| 40 Gy_3 | 0.13 | 0.17 | 0.51 | 0.51 | 0.56 | 0.48 | 0.09 | 0.15 | 0.15 | 0.30 | 0.37 | 0 |
| 10 Gy Mutant and Control Lines | DFBI | DTFF | DTW |
|---|---|---|---|
| 10/46 | 224 | 294 | 50 |
| 10/27 | 405 | 457 | 107 |
| 10/39 | 463 | 512 | 79 |
| 10/60 | 595 | 645 | 86 |
| 10/118 | 574 | 669 | 53 |
| 10/7 | 613 | 678 | 81 |
| C/4 | 906 | 954 | 85 |
| C/5(1) | 886 | 923 | 126 |
| C/5(2) | 886 | 923 | 76 |
| C/8 | 910 | 969 | 87 |
| C/13 | 790 | 862 | 101 |
| C/11 | 870 | 961 | 52 |
| C/6 | 1067 | 1112 | 79 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sherpa, R.; Devadas, R.; Bolbhat, S.N.; Nikam, T.D.; Penna, S. Gamma Radiation Induced In-Vitro Mutagenesis and Isolation of Mutants for Early Flowering and Phytomorphological Variations in Dendrobium ‘Emma White’. Plants 2022, 11, 3168. https://doi.org/10.3390/plants11223168
Sherpa R, Devadas R, Bolbhat SN, Nikam TD, Penna S. Gamma Radiation Induced In-Vitro Mutagenesis and Isolation of Mutants for Early Flowering and Phytomorphological Variations in Dendrobium ‘Emma White’. Plants. 2022; 11(22):3168. https://doi.org/10.3390/plants11223168
Chicago/Turabian StyleSherpa, Rubina, Ramgopal Devadas, Sadashiv Narayan Bolbhat, Tukaram Dayaram Nikam, and Suprasanna Penna. 2022. "Gamma Radiation Induced In-Vitro Mutagenesis and Isolation of Mutants for Early Flowering and Phytomorphological Variations in Dendrobium ‘Emma White’" Plants 11, no. 22: 3168. https://doi.org/10.3390/plants11223168
{kind=link}