
Taxonomy Complexity of Some Tyrrhenian Endemic Limonium Species Belonging to L. multiforme Group (Plumbaginaceae): New Insights from Molecular and Morphometric Analyses

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
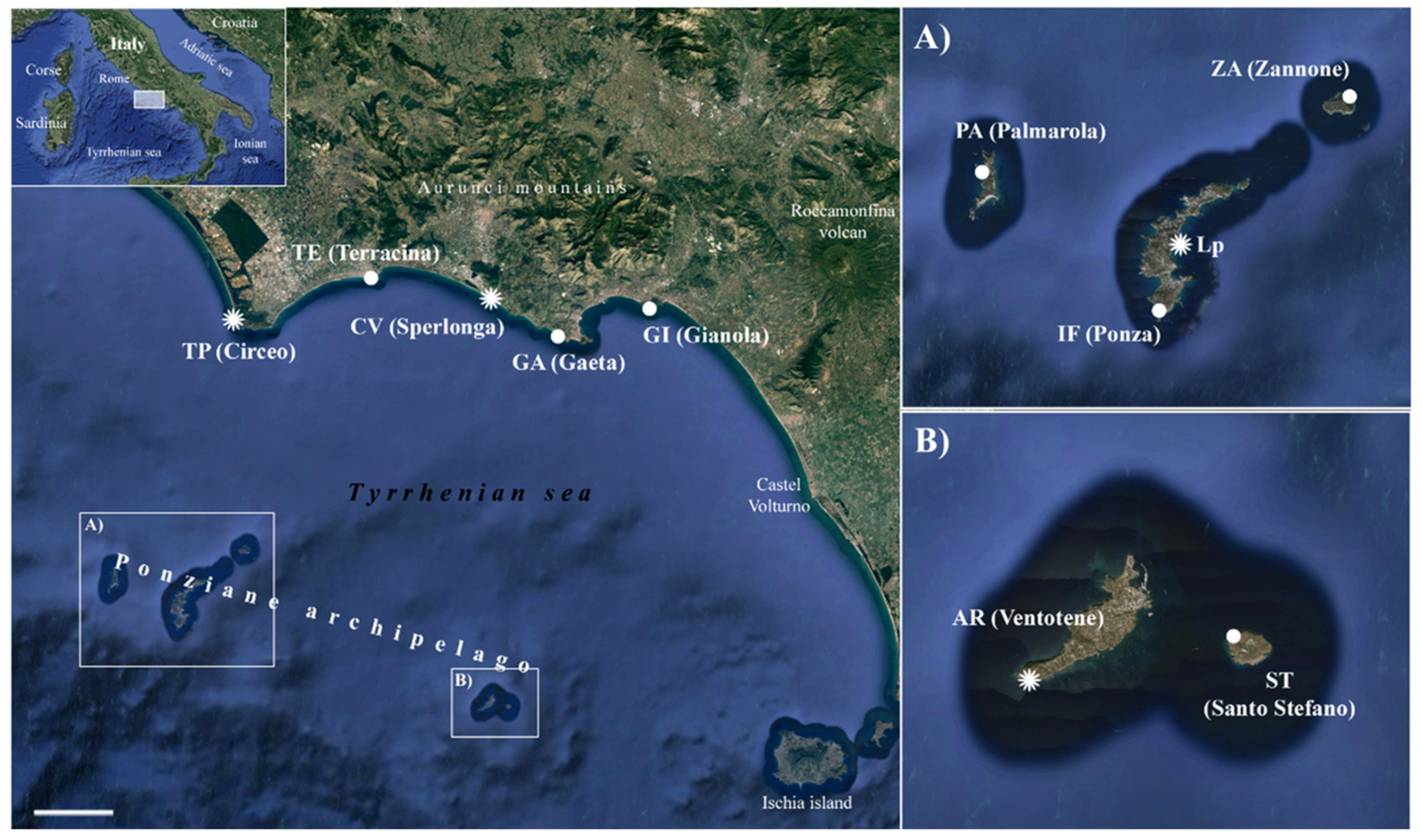
2.1. Plant Material
2.2. Molecular Analysis
2.3. Genomic DNA Isolation
2.4. PCR Amplification and Sequence Analyses
2.5. Data Analysis
2.6. Morphometric Analysis
3. Results
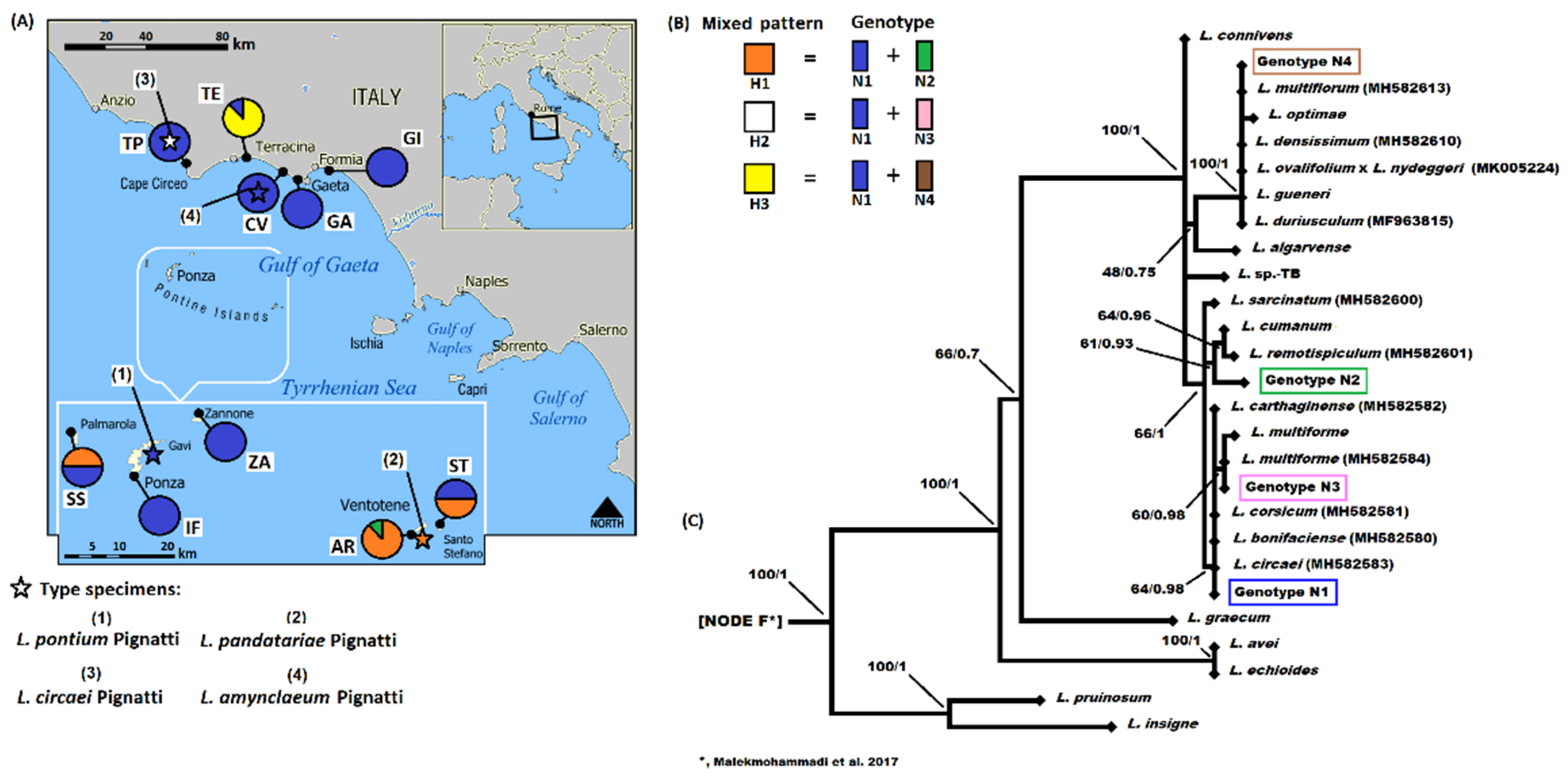
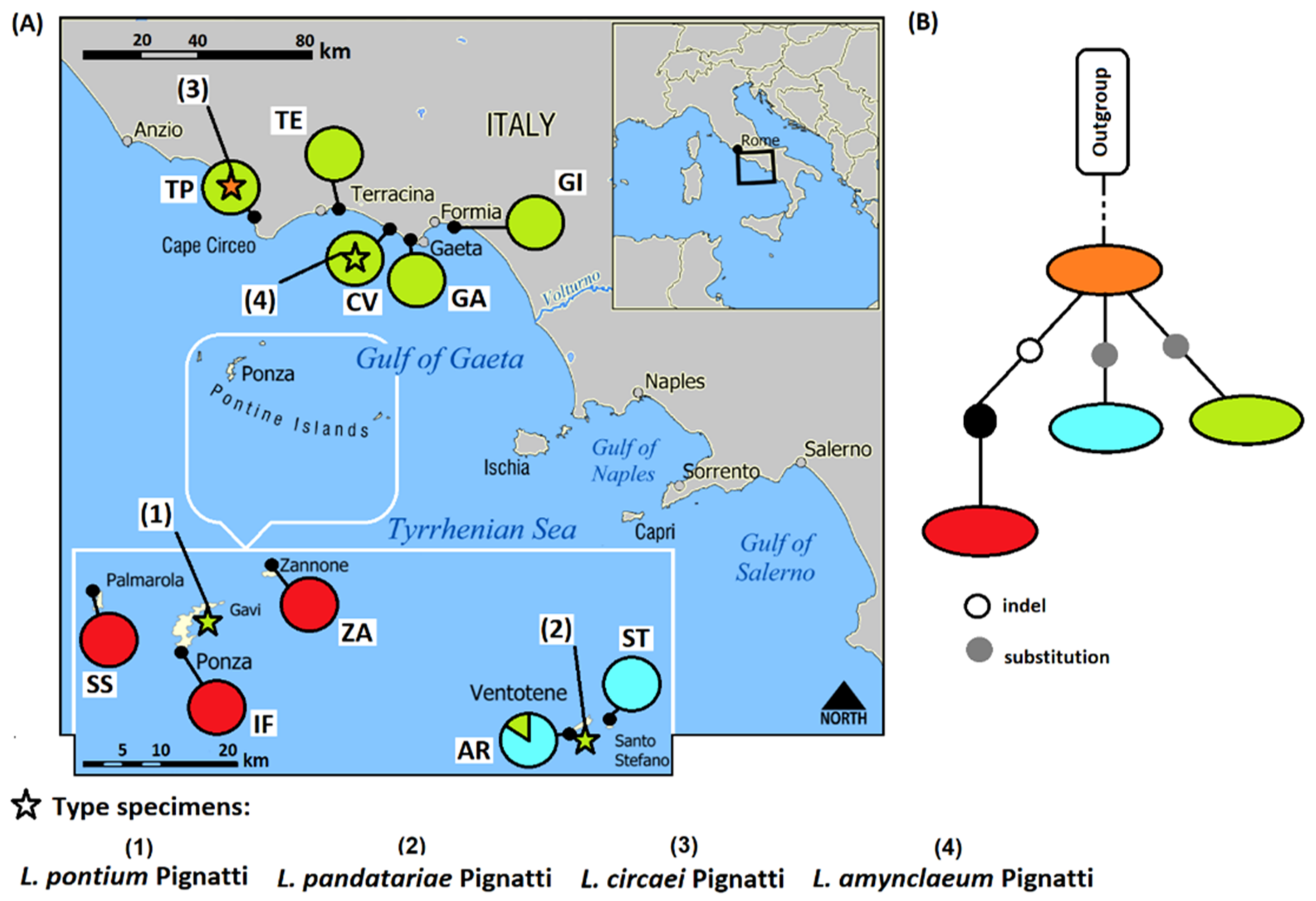
3.1. Molecular Analyses
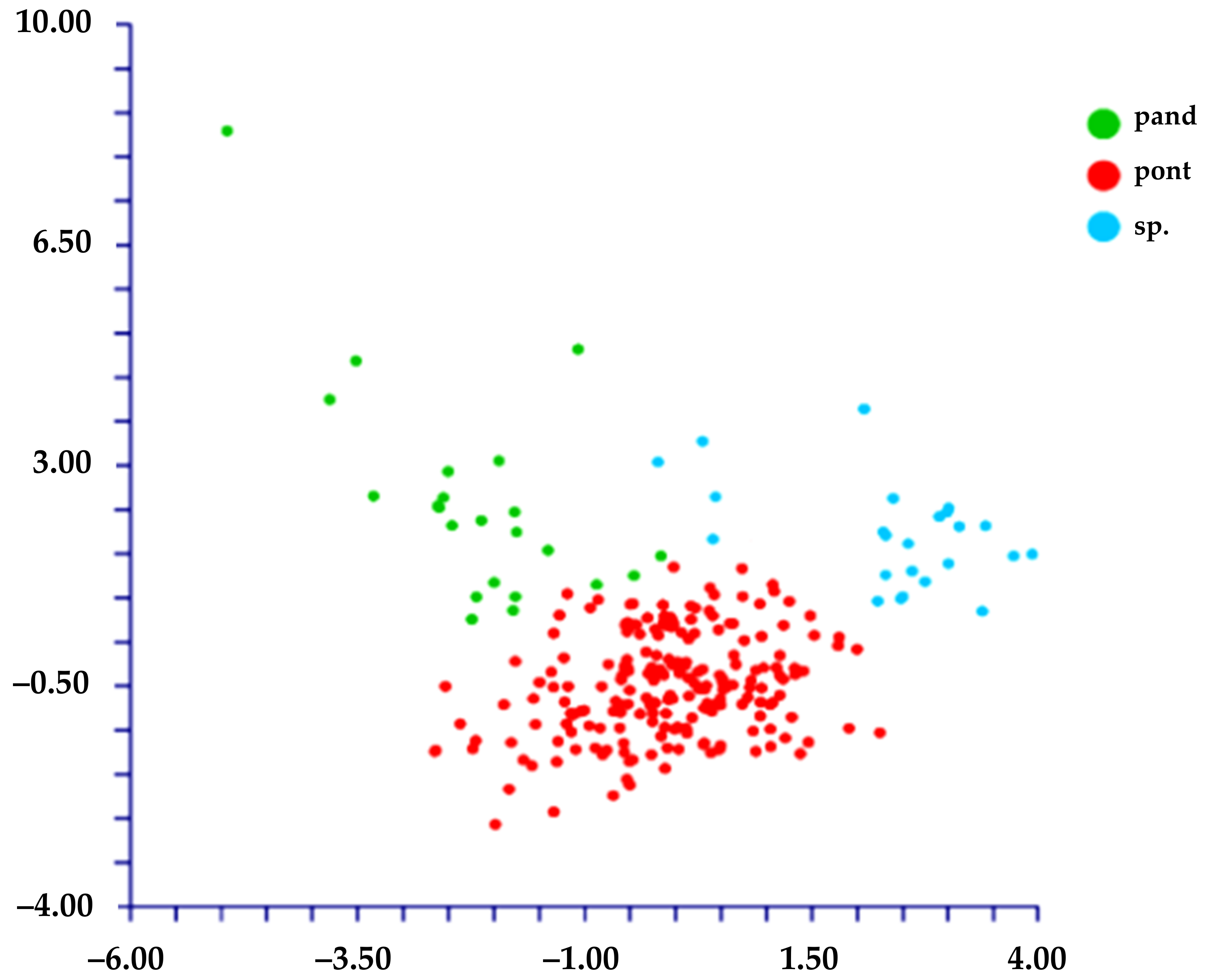
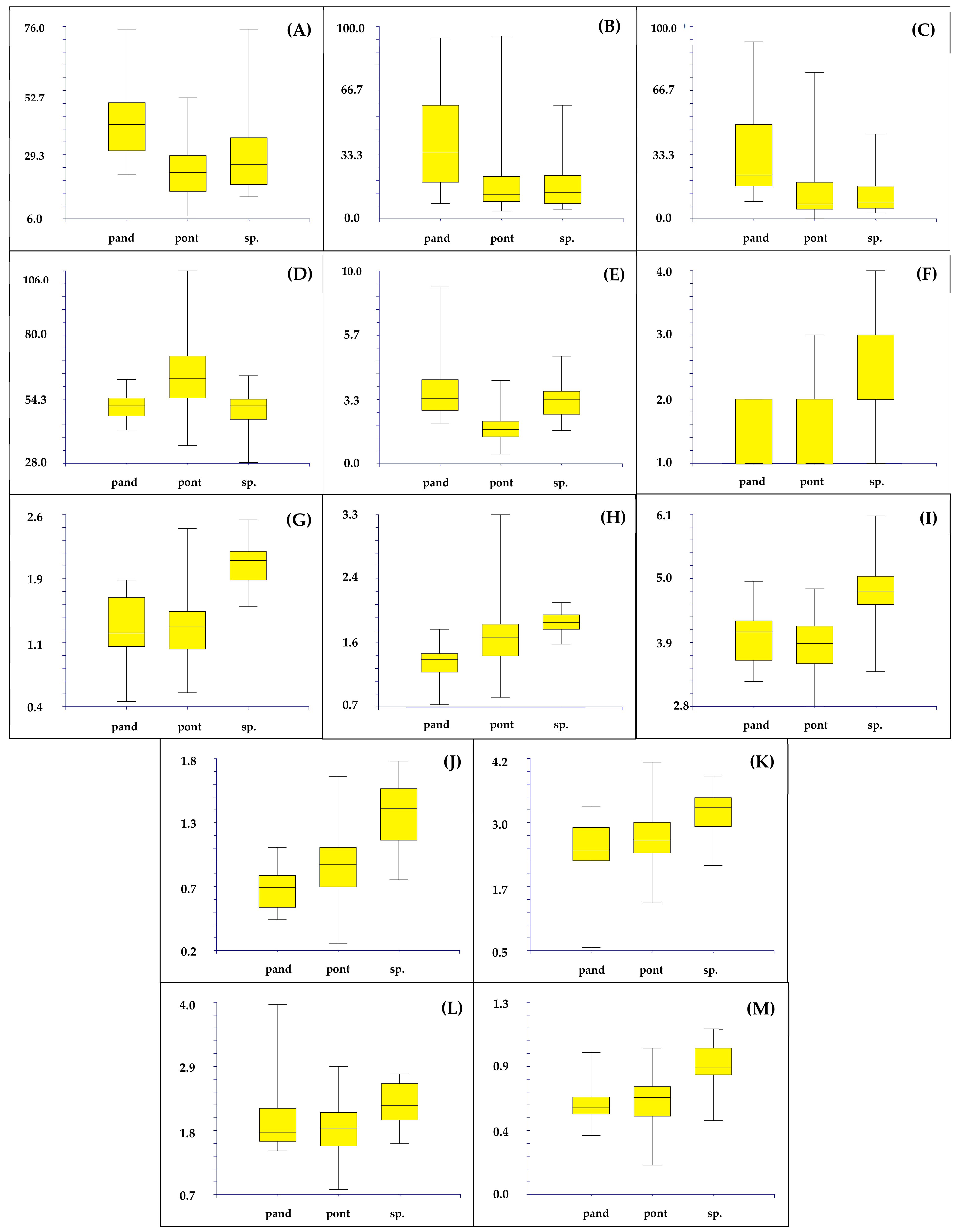
3.2. Morphometric Analysis
- (1)
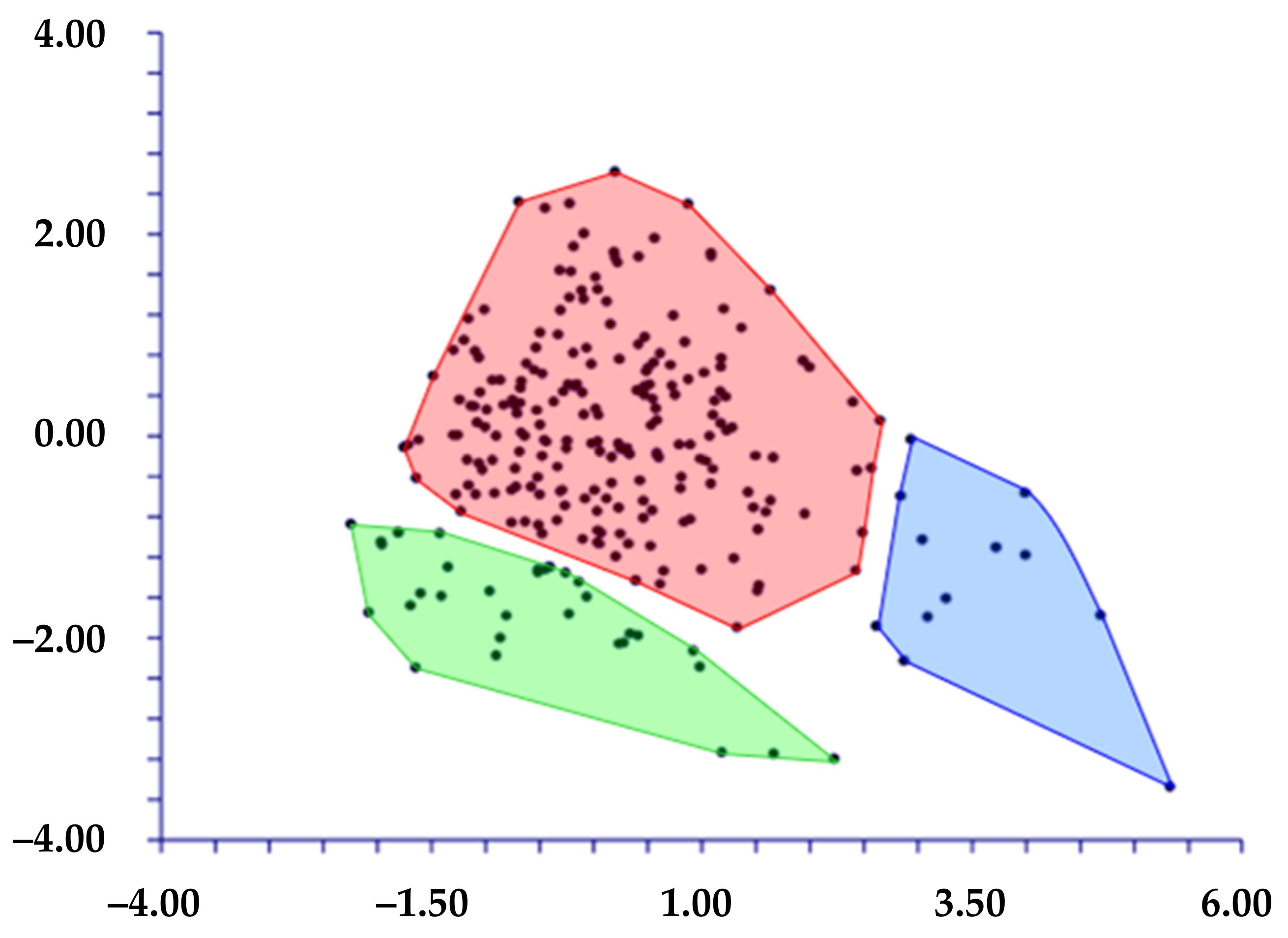
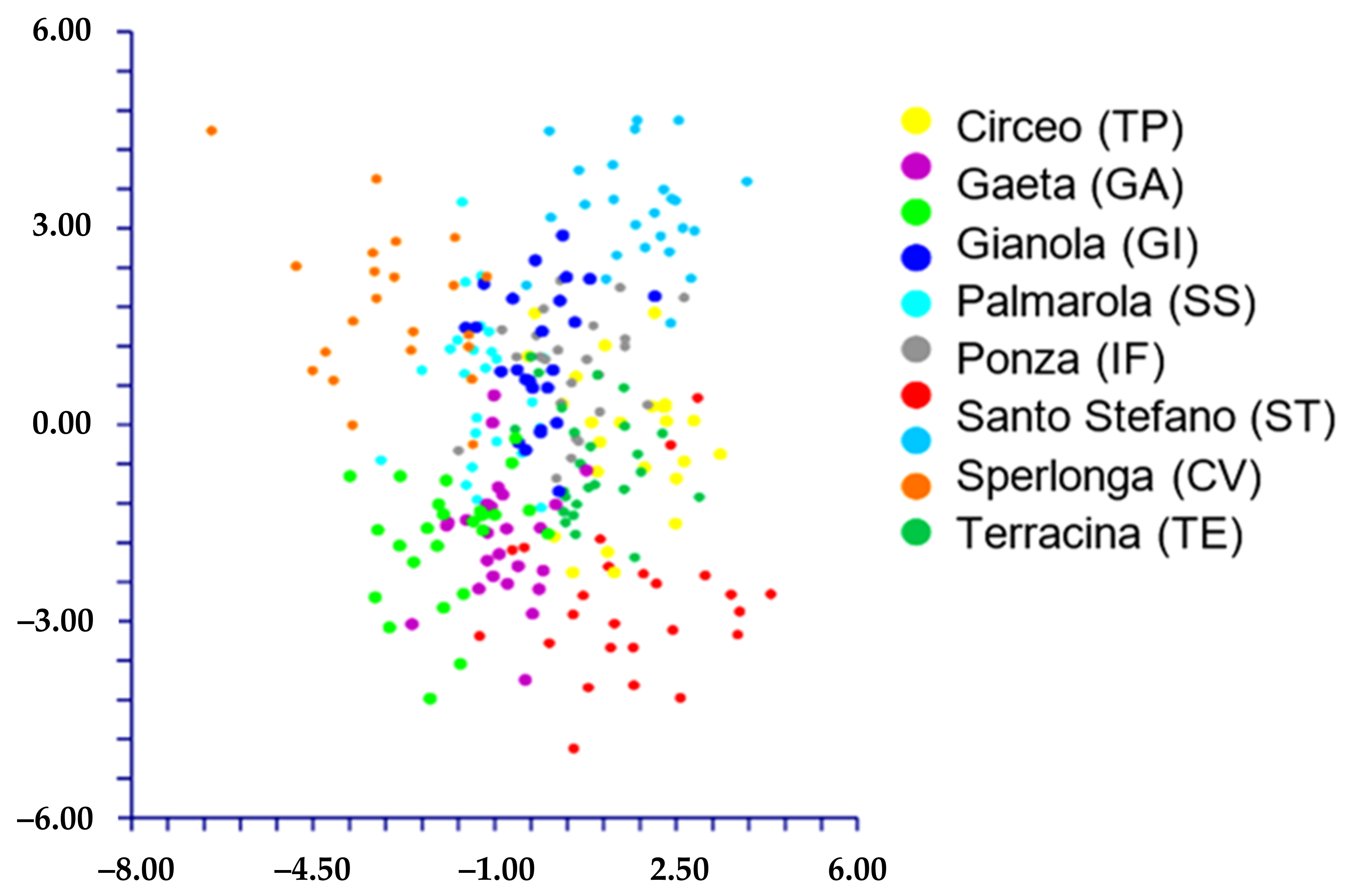
- When we classified the populations using the localities’ names (= ten groups; see Table 1), DA predicted four groups (Figure 6) based on the first two discriminant functions which explain 72.2% of the total variation (eigenvalues: 40.5% (1st function) and 31.7% (2nd function)). These four groups, partially overlapped, correspond to the following localities: A) AR (Ventotene), B) CV (Sperlonga), C) TE (Terracina), and D) a large group including the remaining localities. The matrix of actual/predicted groups displays high percentages along the diagonal (whose values reveal the matching of actual and predicted observations for each group) for CV (100%), TE (96%), and AR (82%), whereas low or very low percentages characterized the diagonal values of the other groups. The value of correct classification is low (61.8%).
- (2)
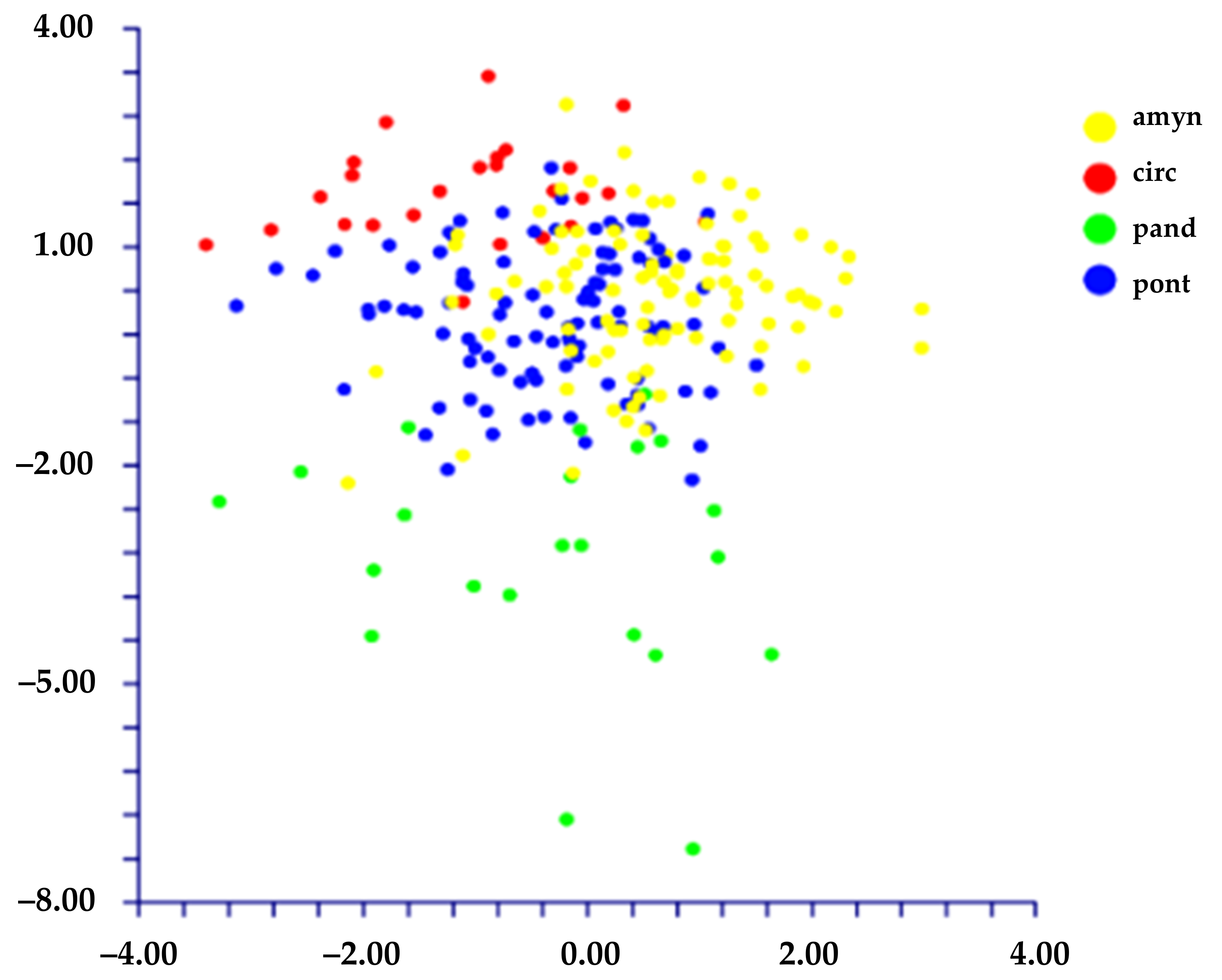
- When we classified the populations using the names of the species (= four groups, L. amynclaeum, L. circaei, L. pandatariae, and L. pontium), DA predicted two partially overlapped groups (Figure 7) based on the first two discriminant functions which explain 84.5% of the total variation (eigenvalues: 64.9% (1st function) and 19.6% (2nd function)). These two groups correspond to (A) L. pantadariae (one locality, AR (Ventotene); see Table 1) and (B) a group comprising all the individuals identified as L. amynclaeum, L. circaei, and L. pontium and collected in the remaining nine localities (coast of Lazio and the islands of the Ponziane archipelago except Ventotene; see Table 1). In fact, the matrix of actual/predicted groups displays a percentage of the diagonal value of L. pandatariae of 56%. The value of correct classification is low (57.5%).


4. Discussion
5. Taxonomic Treatment
Diagnostic Key
| 9. Leaves 3.6–5.3 mm wide……………………………………………………………………………………………… 10 |
| 10. Spikelets 1–2 per cm; inner bract 3.0–3.5 mm long…………… Limoniumremotispiculum |
| 10. Spikelets 2–6 per cm; inner bract 3.7–4.3 mm long……………………………………… 11 |
| 11. Spikes 25–80 mm long; outer bract 1.5–2.0 mm long………… Limonium brutium Brullo |
| 11. Spikes 8–18 mm long; outer bract 1.1–1.6 mm long …………… Limonium pontium s.lat. |
| 11a. Flowers per spikelet (1–)2–3(–4); outer bract 1.8–2.1 mm long; inner bract 4.5–5.1 mm long; limb 1.1–1.6 mm long ………………………………………………… subsp. terracinense |
| 11b. Flowers per spikelet 1–2(–3); outer bract 1.0–1.6 mm
long; inner bract 3.5–4.2 mm long; limb 0.7–1.1 mm long………………………………………....................................... 12 |
| 12a. Leaves 14.2–29.3 mm long; average angle between branches
49–54°; sterile branches 22–44; fertile branches 19–58; middle bracts 1.2–1.4 × 0.5– 0.8…………………………………………………………................................ subsp. pandatariae |
| 12b. Leaves 30.5–48.0 mm long; average angle between branches
54–70°; sterile branches 6–19; fertile branches 8–21; middle bracts 1.4–1.9 × 0.7– 1.1………………………………………………………………………………… subsp.pontium |
| 9. Leaves 5.0–11.0 mm wide…………………………………………………………………… 13 |
| 13. Leaves flat, densely arranged along the caudice
spikelets 2–3 per cm…………………………………………………………………… Limonium gorgonae Pignatti |
| 13. Leaves with margin revolute to convolute laxly arranged along the caudice; spikelets 3–4 per cm ………………………………………………………………… Limoniummultiforme |
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- POWO. Plants of the World Online. Facilitated by the Royal Botanic Gardens, Kew. Published on the Internet. Available online: http://www.plantsoftheworldonline.org/ (accessed on 9 March 2021).
- Erben, M.; Del Guacchio, E. Limonium divaricatum (Plumbaginaceae), the Arabian Phoenix of sea-lavenders. Phytotaxa 2021, 516, 178–186. [Google Scholar] [CrossRef]
- Bogdanović, S.; Brullo, S. Taxonomic revision of the Limonium cancellatum group (Plumbaginaceae) in Croatia. Phytotaxa 2015, 215, 1. [Google Scholar] [CrossRef]
- Malekmohammadi, M.; Akhani, H.; Borsch, T. Phylogenetic relationships of Limonium (Plumbaginaceae) inferred from multiple chloroplast and nuclear loci. Taxon 2017, 66, 1128–1146. Available online: https://www.jstor.org/stable/26824606 (accessed on 9 March 2021). [CrossRef]
- Cortinhas, A.; Erben, M.; Paes, A.P.; Santo, D.E.; Guara-Requena, M.; Caperta, A.D. Taxonomic complexity in the halophyte Limonium vulgare and related taxa (Plumbaginaceae): Insights from analysis of morphological, reproductive and karyological data. Ann. Bot. 2015, 115, 369–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubitzki, K. Plumbaginaceae Juss. In The families and Genera of Vascular Plants; Kubitzki, K., Rohwer, J.G., Bittrich, V., Eds.; Springer: Berlin, Germany, 1993; Volume 2, pp. 523–530. [Google Scholar]
- Lledó, M.D.; Erben, M.; Crespo, M.B. Myriolepis, a new genus segregated from Limonium (Plumbaginaceae). Taxon 2003, 52, 67–73. [Google Scholar] [CrossRef] [Green Version]
- Lledó, M.D.; Karis, O.P.; Crespo, M.B.; Fay, M.F.; Chase, M.W. Endemism and Evolution in Macaronesian and Mediterranean Limonium Taxa. In The Biology of Island Floras; Bramwell, D., Caujapé-Castells, J., Eds.; Cambridge University Press: New York, NY, USA, 2011; pp. 325–337. [Google Scholar]
- Akhani, H.; Malekmohammadi, M.; Mahdavi, P.; Gharibiyan, A.; Chase, M.W. Phylogenetics of the Irano-Turanian taxa of Limonium (Plumbaginaceae) based on ITS nrDNA sequences and leaf anatomy provides evidence for species delimitation and relationships of lineages. Bot. J. Linn. Soc. 2013, 171, 519–550. [Google Scholar] [CrossRef] [Green Version]
- Koutroumpa, K.; Theodoridis, S.; Warren, B.H.; Jiménez, A.; Celep, F.; Doğan, M.; Romeiras, M.M.; Santos-Guerra, A.; Fernández-Palacios, J.M.; Caujapé-Castells, J.; et al. An expanded molecular phylogeny of Plumbaginaceae, with emphasis on Limonium (sea lavenders): Taxonomic implications and biogeographic considerations. Ecol. Evol. 2018, 8, 12397–12424. [Google Scholar] [CrossRef] [Green Version]
- Baker, H.G. Dimorphism and monomorphism in the Plumbaginaceae III. Correlation of geographical distribution patterns with dimorphism and monomorphism in Limonium. Ann. Bot. 1953, 17, 615–627. [Google Scholar] [CrossRef]
- Ferrer-Gallego, P.P.; Rosselló, R.; Rosato, M.; Rosselló, J.A.; Laguna, E. Limonium albuferae (Plumbaginaceae), a new polyploid species from the Eastern Iberian Peninsula. Phytotaxa 2016, 252, 114–122. [Google Scholar] [CrossRef]
- Vallariello, R.; Iamonico, D.; Del Guacchio, E. Typification of three accepted names in Limonium (Plumbaginaceae). Phytotaxa 2016, 263, 131–138. [Google Scholar] [CrossRef]
- Iamonico, D.; Del Guacchio, E. (2528) Proposal to conserve the name Statice minuta (Limonium minutum) (Plumbaginaceae) with a conserved type. Taxon 2017, 66, 759–760. [Google Scholar] [CrossRef]
- Iamonico, D.; Vallariello, R.; Del Guacchio, E. Nomenclatural and distributional remarks on Limonium tenoreanum (Plumbaginaceae), narrow endemic to southern Italy. Nord. J. Bot. 2017, 35, 445–448. [Google Scholar] [CrossRef]
- Del Guacchio, E.; Erben, M.; Iamonico, D. (2626) Proposal to reject the name Limonium oleifolium (Plumbaginaceae). Taxon 2018, 67, 649. [Google Scholar] [CrossRef]
- Del Guacchio, E.; Brullo, S.; Iamonico, D. (2586) Proposal to reject the name Statice reticulata (Limonium reticulatum) (Plantaginaceae). Taxon 2018, 67, 213–214. [Google Scholar] [CrossRef]
- Del Guacchio, E.; Erben, M.; Iamonico, D. The Linnaean names in Statice (Plumbaginaceae). Taxon 2018, 67, 606–610. [Google Scholar] [CrossRef]
- Dolcher, T.; Pignatti, S. Un’ipotesi sull’evoluzione dei Limonium del bacino mediterraneo. G. Bot. Ital. 1971, 105, 95–107. [Google Scholar] [CrossRef]
- Erben, M. Karyotype differentiation and its consequences in Mediterranean Limonium. Webbia 1979, 34, 409–417. [Google Scholar] [CrossRef]
- Arrigoni, P.V.; Diana, S. Contribution à la connaissance du genre Limonium en Corse. Candollea 1993, 482, 631–677. [Google Scholar]
- Castro, M.; Rosselló, J.A. Karyology of Limonium (Plumbaginaceae) species from the Balearic Islands and the western Iberian Peninsula. Bot. J. Linn. Soc. 2007, 155, 257–272. [Google Scholar] [CrossRef] [Green Version]
- Lledó, M.D.; Crespo, M.B.; Fay, M.F.; Chase, M.W. Molecular phylogenetics of Limonium and related genera (Plumbaginaceae): Biogeographical and systematic implications. Am. J. Bot. 2005, 92, 1189–1198. [Google Scholar] [CrossRef] [Green Version]
- Bartolucci, F.; Peruzzi, L.; Galasso, G.; Albano, A.; Alessandrini, A.; Ardenghi, N.M.G.; Astuti, G.; Bacchetta, G.; Ballelli, S.; Banfi, E.; et al. An updated checklist of the vascular flora native to Italy. Plant Biosyst. 2018, 152, 179–303. Available online: http://hdl.handle.net/11568/1061675 (accessed on 9 March 2021). [CrossRef]
- Domina, G. Plumbaginaceae Juss. Available online: http://ww2.bgbm.org/EuroPlusMed/PTaxonDetail.asp?NameId=28346&PTRefFk=7500000 (accessed on 11 March 2021).
- Brullo, S.; Guarino, R. Limonium Mill. In Flora d’Italia, 2nd ed.; Pignatti, S., Guarino, R., La Rosa, M., Eds.; Edagricole: Milan, Italy, 2019; Volume 4, pp. 456–463. [Google Scholar]
- Brullo, S.; Guarino, R. Limonium Mill. In Flora d’Italia, 2nd ed.; Pignatti, S., Guarino, R., La Rosa, M., Eds.; Edagricole: Milan, Italy, 2017; Volume 2, pp. 18–48. [Google Scholar]
- Pignatti, S. Studi sui Limonium. VIII. Bot. J. Linn. Soc. 1971, 64, 361–370. [Google Scholar]
- Pignatti, S. New species of Limonium from Italy and Tunesia. Webbia 1982, 36, 47–56. [Google Scholar] [CrossRef]
- Bedini, G.; Peruzzi, L. Chrobase.it—Chromosome Numbers for the Italian Flora V. 2.0. Available online: http://bot.biologia.unipi.it/chrobase/ (accessed on 25 October 2021).
- Ferrer-Gallego, P.P.; Rosselló, J.A.; Del Guacchio, E.; Iamonico, D. Typification of the Linnaean name Statice limonium (Plumbaginaceae). Taxon 2018, 67, 191–195. [Google Scholar] [CrossRef]
- Thiers, B. Index Herbariorum: A Global Directory of Public Herbaria and Associated Staff; New York Botanical Garden’s Virtual Herbarium: New York, NY, USA, 2022; Available online: http://sweetgum.nybg.org/ih/ (accessed on 15 March 2022).
- Corriveau, J.L.; Coleman, A.W. Rapid screening method to detect potential biparental inheritance of plastid DNA and results for over 200 Angiosperm species. Am. J. Bot. 1988, 75, 1443–1458. [Google Scholar] [CrossRef]
- Róis, A.S.; Sádio, F.; Paulo, O.S.; Teixeira, G.; Paes, A.P.; Espírito-Santo, D.; Sharbel, T.F.; Caperta, A.D. Phylogeography and modes of reproduction in diploid and tetraploid halophytes of Limonium species (Plumbaginaceae): Evidence for a pattern of geographical parthenogenesis. Ann. Bot. 2016, 117, 27–50. [Google Scholar] [CrossRef] [Green Version]
- Koutroumpa, K.; Warren, B.H.; Theodoridis, S.; Coiro, M.; Romeiras, M.M.; Jiménez, A.; Conti, E. Geo-climatic changes and apomixis as major drivers of diversification in the Mediterranean Sea lavenders (Limonium Mill.). Front. Plant Sci. 2021, 11, 612258. [Google Scholar] [CrossRef]
- Palacios, C.; Rosselló, J.A.; González-Candelas, F. Study of the evolutionary relationships among Limonium species (Plumbaginaceae) using nuclear and cytoplasmic molecular markers. Mol. Phylogenet. Evol. 2000, 14, 232–249. [Google Scholar] [CrossRef] [Green Version]
- Róis, A.S.; Castro, S.; Loureiro, J.; Sádio, F.; Rhazi, L.; Guara-Requena, M.; Caperta, A.D. Genome sizes and phylogenetic relationships suggest recent divergence of closely related species of the Limonium vulgare complex (Plumbaginaceae). Plant Syst. Evol. 2018, 304, 955–967. [Google Scholar] [CrossRef]
- De Castro, O.; Geraci, A.; Mannino, A.M.; Mormile, N.; Santangelo, A.; Troia, A.A. Contribution to the Characterization of Ruppia drepanensis (Ruppiaceae), A Key Species of Threatened Mediterranean Wetlands. Ann. Mo. Bot. Gard. 2021, 106, 1–9. [Google Scholar] [CrossRef]
- Aceto, S.; Caputo, P.; Cozzolino, S.; Gaudio, L.; Moretti, A. Phylogeny and evolution of Orchis and allied genera based on ITS DNA variation: Morphological gaps and molecular continuity. Mol. Phylogenet. Evol. 1999, 13, 67–76. [Google Scholar] [CrossRef]
- De Castro, O.; Di Maio, A.; Lozada García, J.A.; Piacenti, D.; Vázquez-Torres, M.; De Luca, P. Plastid DNA sequencing and nuclear SNP genotyping help resolve the puzzle of central American Platanus. Ann. Bot. 2013, 112, 589–602. [Google Scholar] [CrossRef]
- Taberlet, P.; Gielly, L.; Pautou, G.; Bouvet, J. Universal primers for amplification of three non-coding regions of chloroplast DNA. Plant Mol. Biol. 1991, 17, 1105–1109. [Google Scholar] [CrossRef]
- Prince, L.M. Plastid primers for Angiosperm phylogenetics and phylogeography. Appl. Plant Sci. 2015, 3, 1400085. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. Bioedit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Thornhill, A.H.; Baldwin, B.G.; Freyman, W.A.; Nosratinia, S.; Kling, M.M.; Morueta-Holme, N.; Madsen, T.P.; Ackerly, D.D.; Mishler, B.D. Spatial phylogenetics of the native California flora. BMC Biol. 2017, 15, 96. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarisation in Bayesian phylogenetics using Tracer 1.7. Syst Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [Green Version]
- Kozlov, A.M.; Darriba, D.; Flouri, T.; Morel, B.; Stamatakis, A. RAxML-NG: A fast, scalable, and user-friendly tool for maximum likelihood phylogenetic inference. Bioinformatics 2019, 35, 4453–4455. [Google Scholar] [CrossRef] [Green Version]
- Clement, M.; Posada, D.; Crandall, K. TCS: A computer program to estimate gene genealogies. Mol. Ecol. 2000, 9, 1657–1660. [Google Scholar] [CrossRef] [PubMed]
- Templeton, A.R.; Crandall, K.A.; Sing, C.F. A cladistic analysis of phenotypic associations with haplotypes inferred from restriction endonuclease mapping and DNA sequence data. III. Cladogram estimation. Genetics 1992, 132, 619–633. [Google Scholar] [CrossRef] [PubMed]
- Topliss, J.G.; Costello, R.J. Chance correlations in structure-activity studies using multiple regression analysis. J. Med. Chem. 1972, 15, 1066–1068. [Google Scholar] [CrossRef] [PubMed]
- Graham, M.H. Confronting multicollinearity in ecological multiple regression. ESA 2003, 84, 2809–2815. [Google Scholar] [CrossRef] [Green Version]
- Stewart, S.; Ivy, M.I.; Anslyn, E.V. The use of principal component analysis and discriminant analysis in differential sensing routines. Chem. Soc. Rev. 2013, 43, 70–84. [Google Scholar] [CrossRef]
- Koutstaal, B.P.; Markusse, M.M.; de Munck, W. Aspects of Seed Dispersal by Tidal Movements. In Vegetation between Land and Sea; Huiskes, A.H.L., Blom, C.W.P.M., Rozema, J., Eds.; Springer: Dordrecht, Geobotany, 1987; Volume 11, pp. 226–235. [Google Scholar]
- Archbald, G.; Boyer, K.E. Potential for spread of Algerian sea lavender (Limonium ramosissimum subsp. provinciale) in tidal marshes. Invasive Plant Sci. Manag. 2017, 7, 454–463. [Google Scholar] [CrossRef]
- Brullo, S. Limonium brutium, a new species from S. Italy. Flora Medit. 1992, 2, 109–112. [Google Scholar]
- Moreno, J.; Terrones, A.; Alonso, M.A.; Ana, J.; Crespo, M.B. Limonium tobarrense (Plumbaginaceae), a new species from the southeastern Iberian Peninsula. Phytotaxa 2016, 257, 61–70. [Google Scholar] [CrossRef]
- Turland, N.J.; Wiersema, J.H.; Barrie, F.R.; Greuter, W.; Hawksworth, D.L.; Herendeen, P.S.; Knapp, S.; Kusber, W.-H.; Li, D.-Z.; Marhold, K.; et al. International Code of Nomenclature for Algae, Fungi, and Plants (Shenzhen Code) Adopted by the Nineteenth International Botanical Congress, Shenzhen, China, July 2017; Regnum Vegetabile; Koeltz Botanical Books: Glashütten, Germany, 2018; Volume 159, pp. 1–254. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Name | Population Code | Locality |
|---|---|---|
| Limonium amynclaeum Pignatti | CV | Lazio region, Sperlonga, Torre Capovento *, 13°28′27.54″ E, 41°14′19.65″ N, 30 June 2011, Iberite et Cacciarini |
| Limonium amynclaeum Pignatti | GA | Lazio region, Gaeta, Trecento scalini, 13°31′50.67″ E, 41°13′9.79″ N, 12 August 2015, Iamonico |
| Limonium amynclaeum Pignatti | GI | Lazio region, Gianola, 13°40′29.95″ E, 41°14′47.23″ N, 12 August 2015, Iamonico |
| Limonium amynclaeum Pignatti | TE | Lazio region, Terracina, 13°15′47.30″ E, 41°17′20.17″ N, 13 July 2015, Iberite et Nicolella |
| Limonium circaei Pignatti | TP | Lazio region, Circeo, Torre Paola *, 13° 2′4.50″ E, 41°14′47.06″ N, 13 July 2015, Iberite et Nicolella |
| Limonium pontium Pignatti | IF | Lazio region, Ponza, Il Faro, 12°57′15.90″ E, 40°52′48.09″ N, 29 July 2014, Iberite et Nicolella |
| Limonium pontium Pignatti | SS | Lazio region, Palmarola, San Silverio, 12°51′18.56″ E, 40°56′23.08″ N, 27 June 2016, Iberite et Nicolella |
| Limonium pontium Pignatti | ST | Lazio region, Santo Stefano, 13°27′4.73″ E, 40°47′29.12″ N, 26 July 2015, Iberite et Nicolella |
| Limonium pontium Pignatti | ZA | Lazio region, Zannone, 13°3′38.31″ E, 40°58′17.75″ N, 25 July 2016, Iberite et Nicolella |
| Limonium pandatariae Pignatti | AR | Lazio region, Ventotene, Arco, 13°27′4.73″ E, 40°47′29.12″ N, 27 July 2015, Iberite et Nicolella |
| ITS1 * | ITS2 * | GenBank | Genotype | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 80 | 208 | 384 | 390 | 396 | 420 | 435 | 446 | 449 | 513 | 551 | 583 | ||
| TCCCA | - | C | C | C | C | T | C | G | T | C | - | OP452889 | N1 |
| GGAT- | - | C | C | C | C | C | C | G | T | C | GA | OP452890 | N2 |
| TCCCA | - | C | C | C | C | T | C | A | T | C | - | OP452891 | N3 |
| TCCCT | TA | T | T | T | T | T | A | G | C | T | - | OP452892 | N4 |
| petB-petD * | GenBank | trnL(UAA)-trnF(GAA) * | GenBank | Haplotype | ||
|---|---|---|---|---|---|---|
| petB-petD IGS | petD intron | trnL(UAA) intron | ||||
| 126 | 302 | 742 | 87 | |||
| T | T | C | OP485326 | A6 | OP485330 | A |
| T | C | C | OP485327 | A6 | OP485331 | B |
| C | T | C | OP485328 | A6 | OP485332 | G |
| T | T | T | OP485329 | A5 | OP48533 | R |
| Height |
| Number of fertile branches * |
| Number of sterile branches * |
| Average angle between branches |
| Length of the leaf |
| Width of the leaf |
| Length of ludicule |
| Maximum distance among the spikelets |
| Length of the spike |
| Number of spikelets * |
| Length of the spikelets |
| Length of the outer bract |
| Width of the outer bract |
| Length of the median bract |
| Width of the median bract |
| Length of the inner bract |
| Width of the inner bract |
| Number of flowers per spikelet |
| Length of the tube |
| Length of the limb |
| Length of the corolla lobes |
| Width of the corolla lobes |
| Test Statistic | Test Value | F-Ratio | p (0.05) |
|---|---|---|---|
| Wilks’ Lambda | 0.142926 | 16.38 | 0.00000001 |
| Hotelling–Lawley Trace | 3.2985 | 16.35 | 0.00000001 |
| Pillai’s Trace | 1.242707 | 16.41 | 0.00000001 |
| Roy’s Largest Root | 1.797216 | 17.97 | 0.00000001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iamonico, D.; De Castro, O.; Di Iorio, E.; Nicolella, G.; Iberite, M. Taxonomy Complexity of Some Tyrrhenian Endemic Limonium Species Belonging to L. multiforme Group (Plumbaginaceae): New Insights from Molecular and Morphometric Analyses. Plants 2022, 11, 3163. https://doi.org/10.3390/plants11223163
Iamonico D, De Castro O, Di Iorio E, Nicolella G, Iberite M. Taxonomy Complexity of Some Tyrrhenian Endemic Limonium Species Belonging to L. multiforme Group (Plumbaginaceae): New Insights from Molecular and Morphometric Analyses. Plants. 2022; 11(22):3163. https://doi.org/10.3390/plants11223163
Chicago/Turabian StyleIamonico, Duilio, Olga De Castro, Emanuela Di Iorio, Gianluca Nicolella, and Mauro Iberite. 2022. "Taxonomy Complexity of Some Tyrrhenian Endemic Limonium Species Belonging to L. multiforme Group (Plumbaginaceae): New Insights from Molecular and Morphometric Analyses" Plants 11, no. 22: 3163. https://doi.org/10.3390/plants11223163