
Correlation between Vegetation Structure and Species Diversity in Traditional Villages in Karst Topographic Regions of the Zunyi City, China



Abstract
:1. Introduction
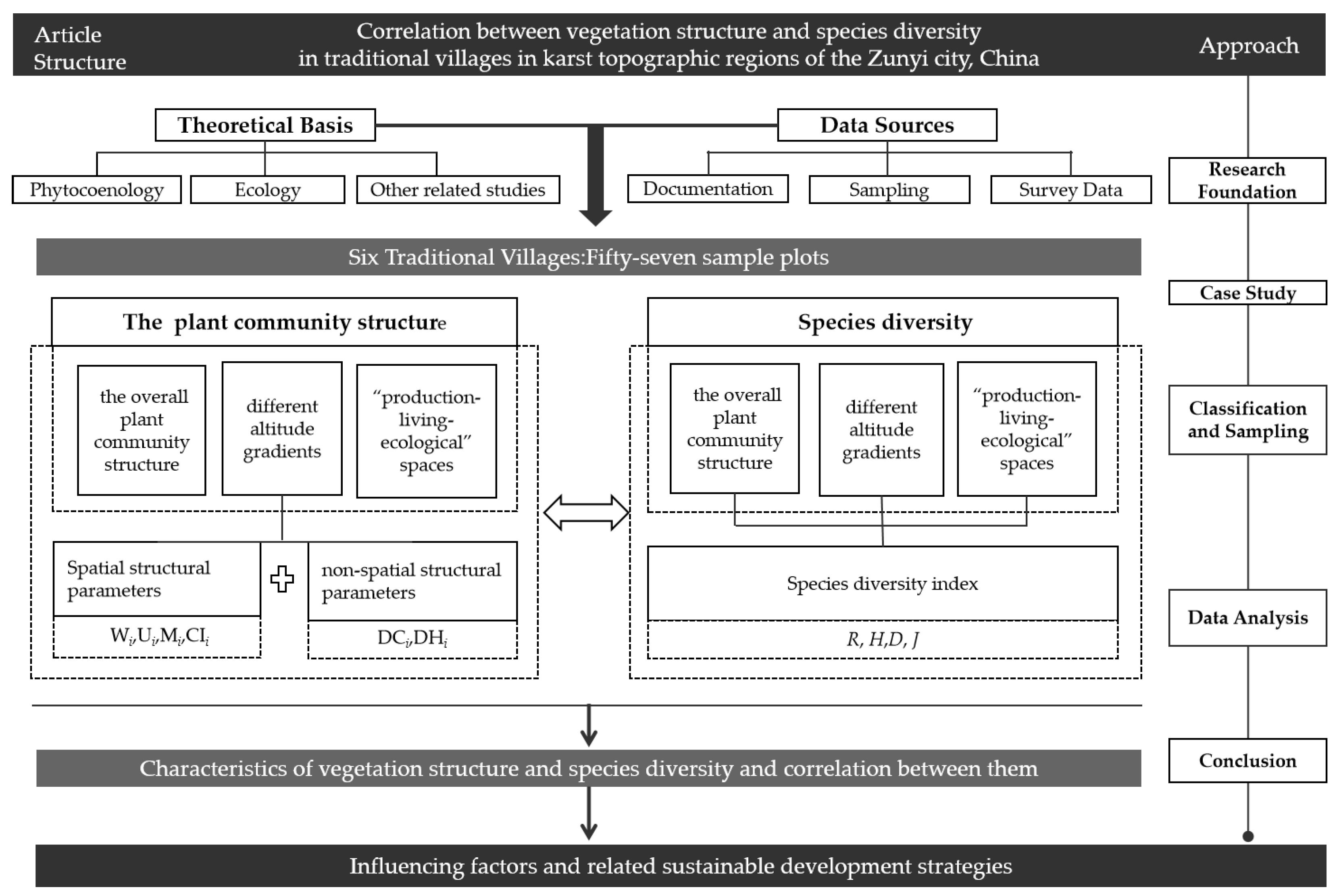
2. Materials and Methods
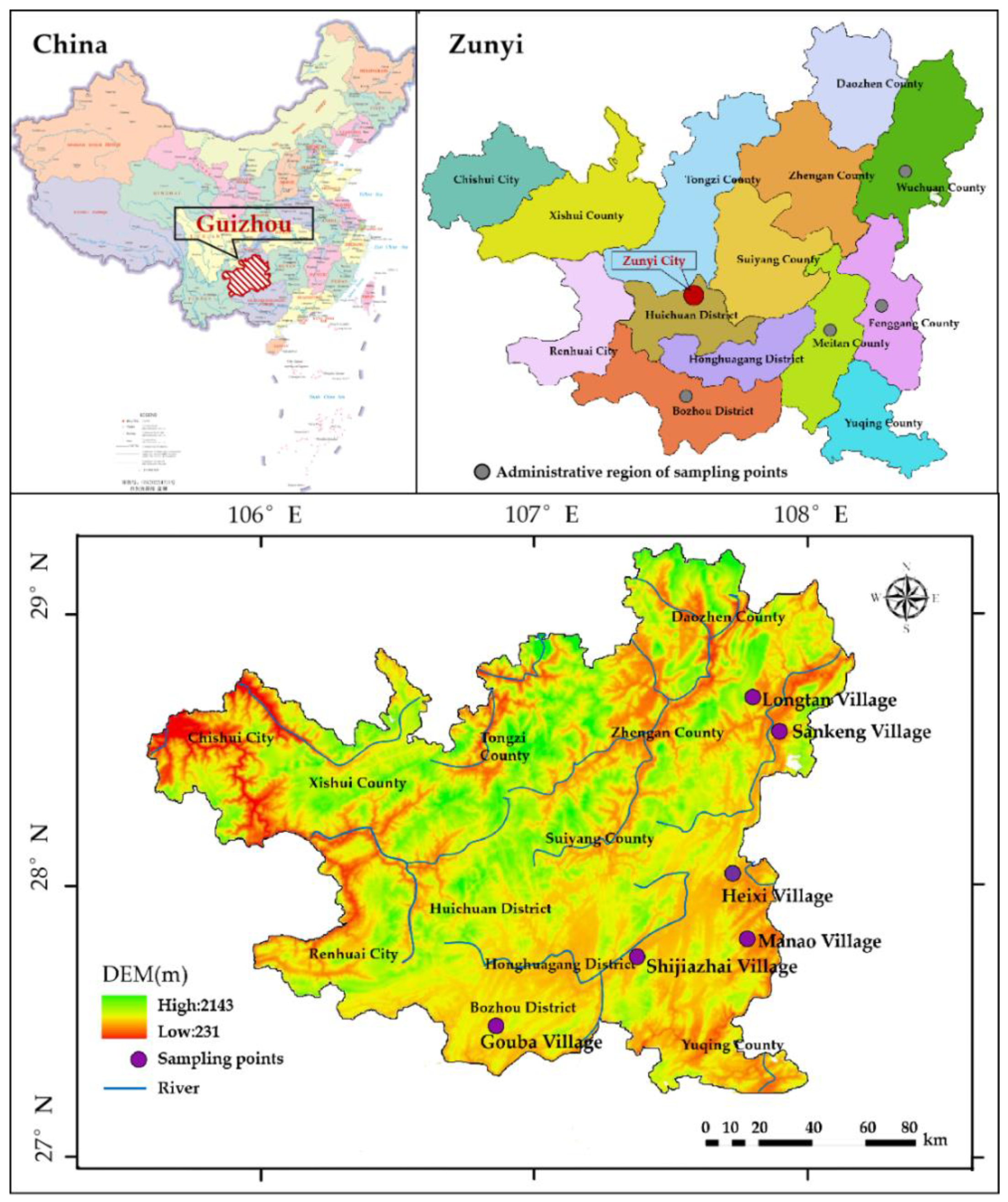
2.1. Study Area
2.2. Research Methodology
2.2.1. Village Selection and Overview
2.2.2. Determination of Parameter Values for Each Analytical Perspective
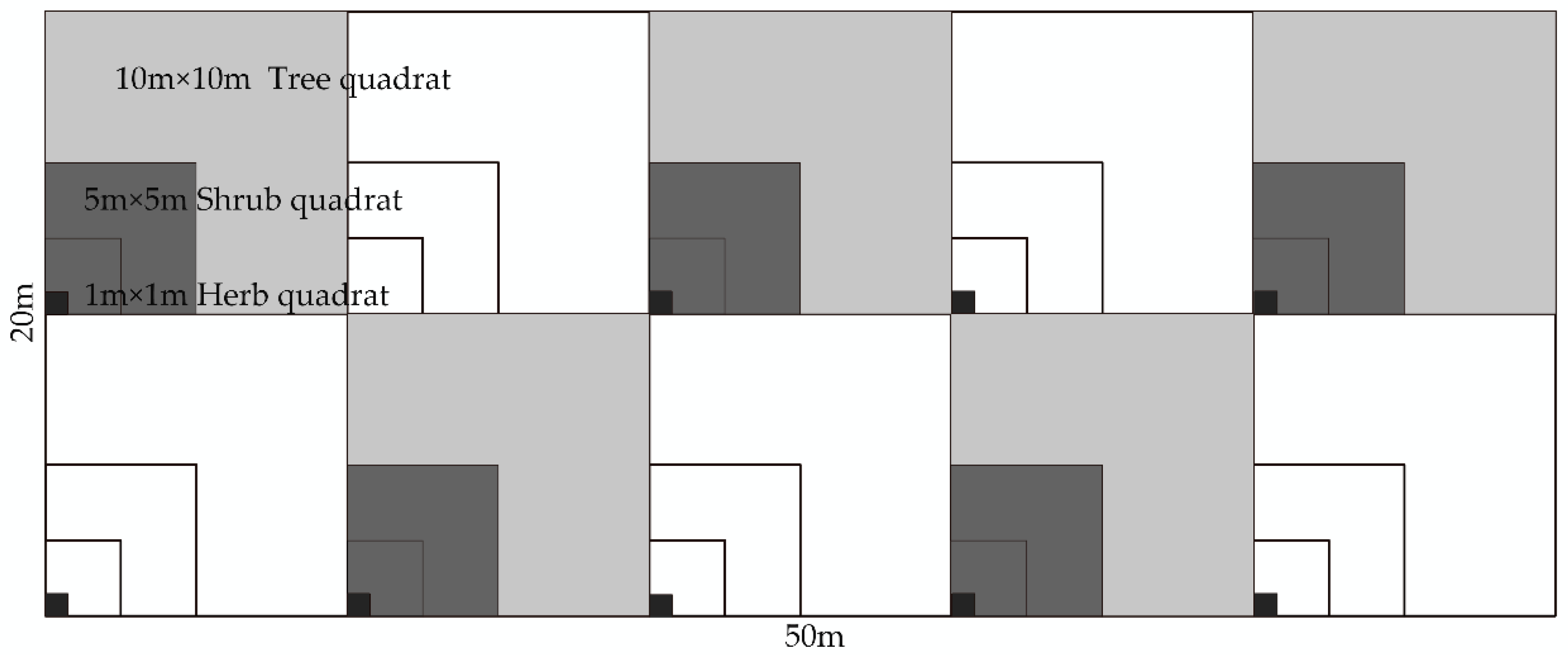
2.2.3. Sample Plot Setting and Survey
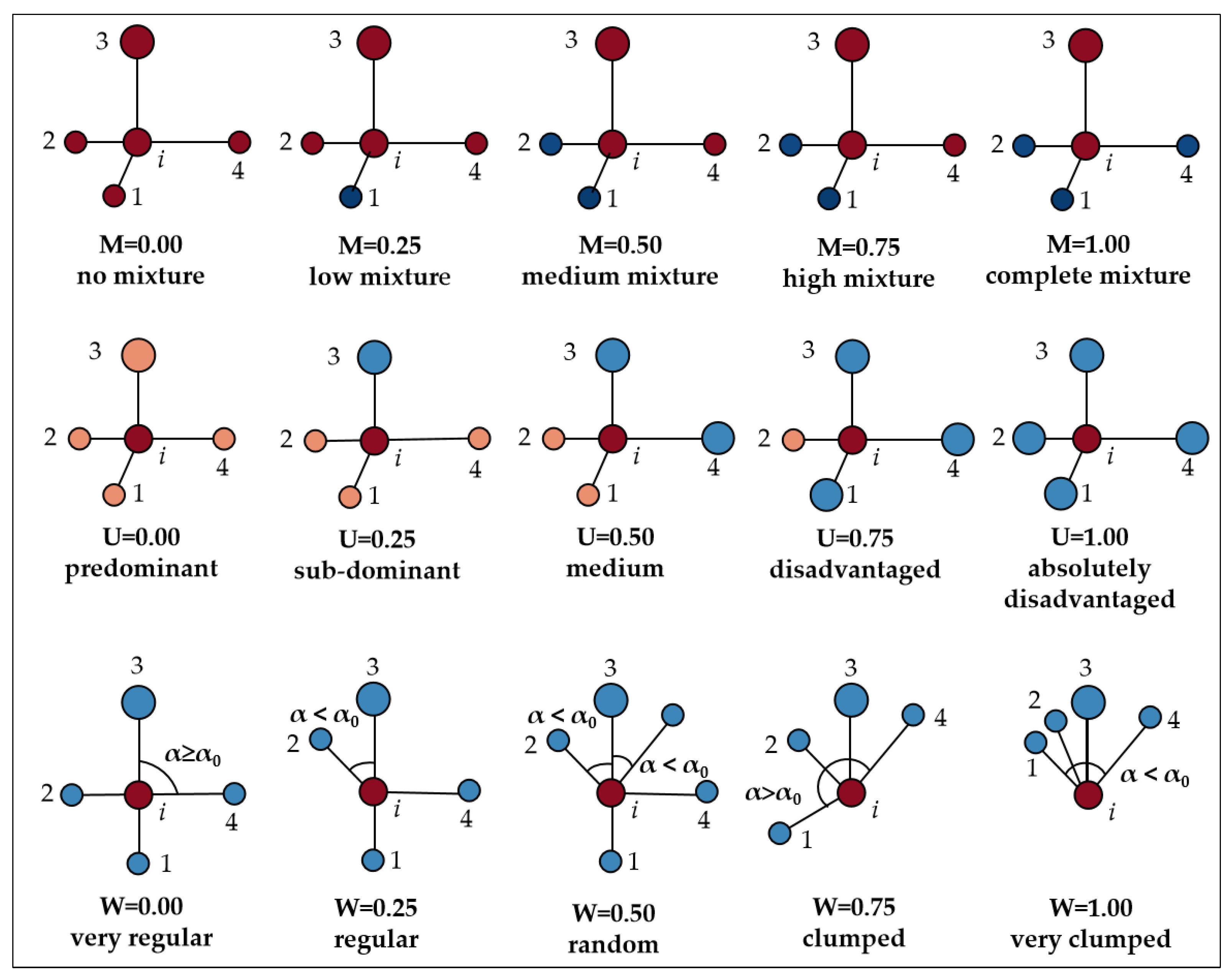
2.2.4. Determination of Plant Community Spatial Structure and Non-Spatial Structure Parameters
2.2.5. Determination of Plant Community Species Diversity Index
2.3. Data Processing
3. Results
3.1. Analysis of Plant Community Structure in Traditional Villages in Karst Topographic Regions of the Zunyi City
3.1.1. Analysis of the Overall Plant Community Structure of Traditional Villages in Karst Topographic Regions of the Zunyi
3.1.2. Analysis of Plant Community Structure at Different Altitude Gradients
3.1.3. Analysis of the Plant Community Structure under the “Production-Living-Ecological” Spaces

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter Name | Uniform Angle Index (W) | Dominance (U) | Mingling (M) | Competition Index (CI) | Growth Vitality (DC) | Stability (DH) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| W > 0.517 | 0.475 < W < 0.517 | W < 0.475 | 0 | (0, 0.25] | (0.25, 0.5] | (0.5, 0.75] | (0.75, 1] | 0 | (0, 0.25] | (0.25, 0.5] | (0.5, 0.75] | (0.75, 1] | —— | —— | —— | |
| Specific meaning | Aggregate distribution | Random distribution | Evenly distributed | dominance | sub-dominant | mean state | disadvantages | Absolute | Zero Mixing | Weakly mixed | Moderately mixed | Strength mixing | Very strong | —— | —— | —— |
| Percentage | 33.33% | 28.07% | 38.60% | -- | 3.51% | 50.88% | 45.61% | -- | 5.26% | 8.77% | 10.53% | 52.63% | 22.81% | 1.715 ± 0.515 | 0.333 ± 0.132 | 0.645 ± 0.209 |
| Altitude (m) | Uniform Angle Index (W) | Dominance (U) | Mingling (M) | Competition Index (CI) | Growth Vitality (DC) | Stability (DH) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| W > 0.517 | 0.475 < W < 0.517 | W < 0.475 | 0 | (0, 0.25] | (0.25, 0.5] | (0.5, 0.75] | (0.75, 1] | 0 | (0, 0.25] | (0.25, 0.5] | (0.5, 0.75] | (0.75, 1] | —— | —— | —— | |
| Specific meaning | aggregated distribution | random distribution | uniform distribution | dominance | sub-dominant | mean state | disadvantages | absolute disadvantage | zero degree mixed | weak mixed | moderate mixed | intensity mixing | very strong mixed | —— | —— | —— |
| 600–700 | 60.00% | 10.00% | 30.00% | —— | —— | 20.00% | 80.00% | —— | —— | 20.00% | 10.00% | 50.00% | 20.00% | 1.913 ± 0.303a | 0.334 ± 0.135a | 0.706 ± 0.265a |
| 700–800 | 27.27% | 18.18% | 54.55% | —— | 9.09% | 72.73% | 18.18% | —— | 9.09% | —— | —— | 63.64% | 27.27% | 1.721 ± 0.600ab | 0.293 ± 0.091a | 0.539 ± 0.162b |
| 800–900 | 11.11% | 38.89% | 50.00% | —— | —— | 50.00% | 44.44% | 5.56% | —— | —— | 16.67% | 55.56% | 27.78% | 1.605 ± 0.421b | 0.351 ± 0.172a | 0.676 ± 0.185a |
| 1000–1100 | 25.00% | 33.33% | 41.67% | —— | 8.33% | 58.33% | 33.33% | —— | 16.67% | 25.00% | 16.67% | 33.33% | 8. 33% | 1.569 ± 0.675b | 0.344 ± 0.104a | 0.632 ± 0.242ab |
| 1100–1160 | 57.14% | 28.57% | 14.29% | —— | —— | 57.14% | 42.86% | —— | —— | —— | 14.29% | 42.86% | 42.86% | 1.917 ± 0.471ab | 0.333 ± 0.126a | 0.659 ± 0.155ab |
| “PLE Spaces” | Uniform Angle Index (W) | Dominance (U) | Mingling (M) | Competition Index (CI) | Growth Vitality (DC) | Stability (DH) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| W > 0.517 | 0.475 < W < 0.517 | W < 0.475 | 0 | (0, 0.25] | (0.25, 0.5] | (0.5, 0.75] | (0.75, 1] | 0 | (0, 0.25] | (0.25, 0.5] | (0.5, 0.75] | (0.75, 1] | —— | —— | —— | |
| Specific meaning | Aggregate distribution | Random distribution | uniform distribution | dominance | sub-dominant | mean state | disadvantages | Absolute | Zero Mixing | Weak mixed | Moderately mixed | intensity mixing | Very strong | —— | —— | —— |
| produce | 22.22% | 44.44% | 33.33% | —— | 22.22% | 44.44% | 33.33% | —— | 11.11% | 11.11% | —— | 66.67% | 11.11% | 1.382 ± 0.472a | 0.297 ± 0.100ab | 0.500 ± 0.130a |
| Living | 37.93% | 27.59% | 34.48% | —— | —— | 48.28% | 51.72% | —— | —— | 3.45% | 10.34% | 48.28% | 37.93% | 1.707 ± 0.425ab | 0.375 ± 0.152a | 0.682 ± 0.210b |
| Ecology | 26.32% | 15.79% | 57.89% | —— | —— | 57.89% | 36.84% | 5.26% | 10.53% | 15.79% | 21.05% | 42.11% | 10.53% | 1.884 ± 0.611b | 0.285 ± 0.092b | 0.657 ± 0.216ab |
3.2. Analysis of Plant Species Diversity in Traditional Villages in Karst Topographic Regions of the Zunyi
3.2.1. Overall Plant Community Species Diversity in Traditional Villages in Karst Topographic Regions of the Zunyi
3.2.2. Species Diversity of Plant Communities at Different Altitude Gradients
3.2.3. Species Diversity Analysis of Plant Communities under the “Production-Living-Ecological” Spaces
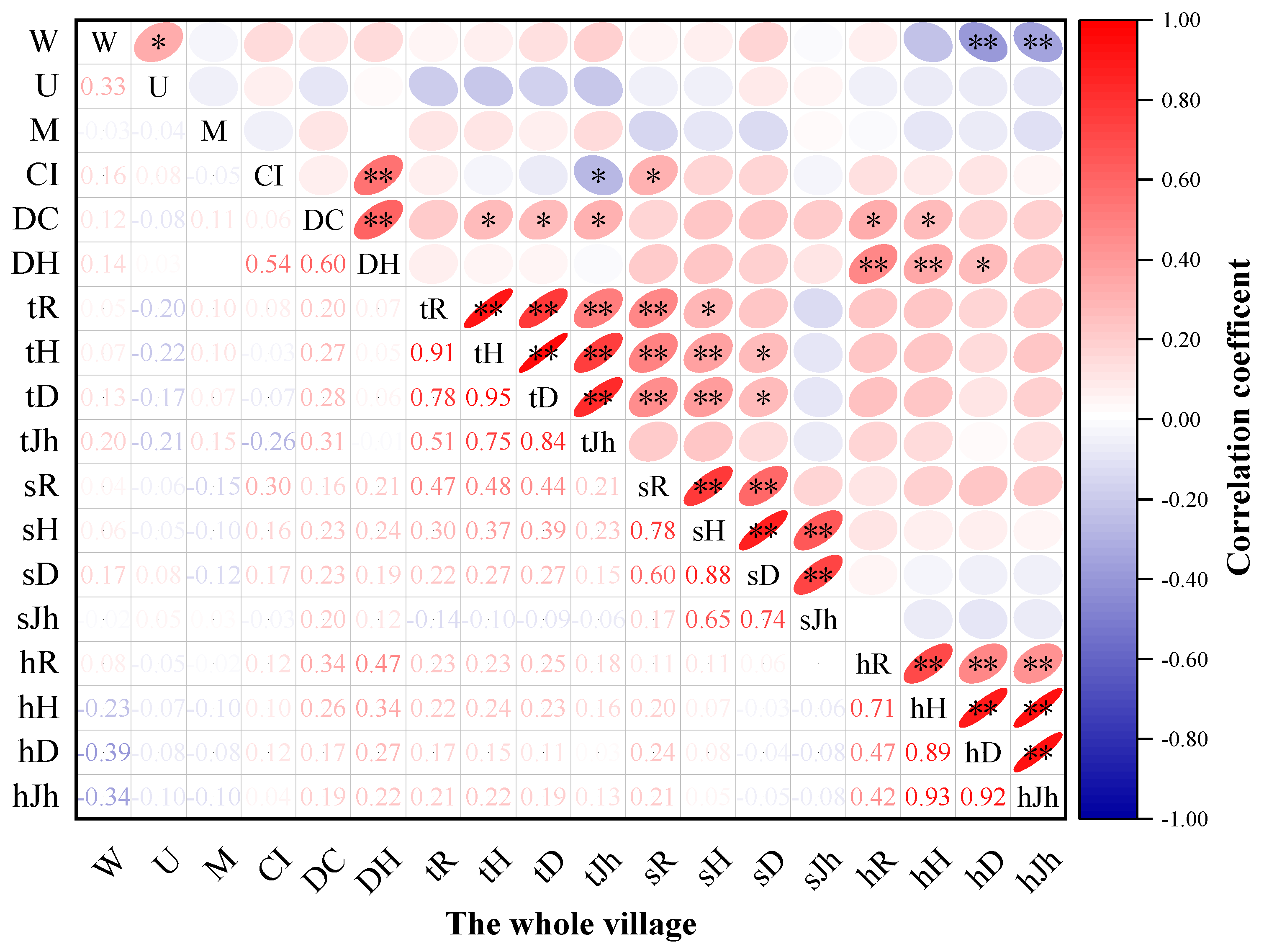
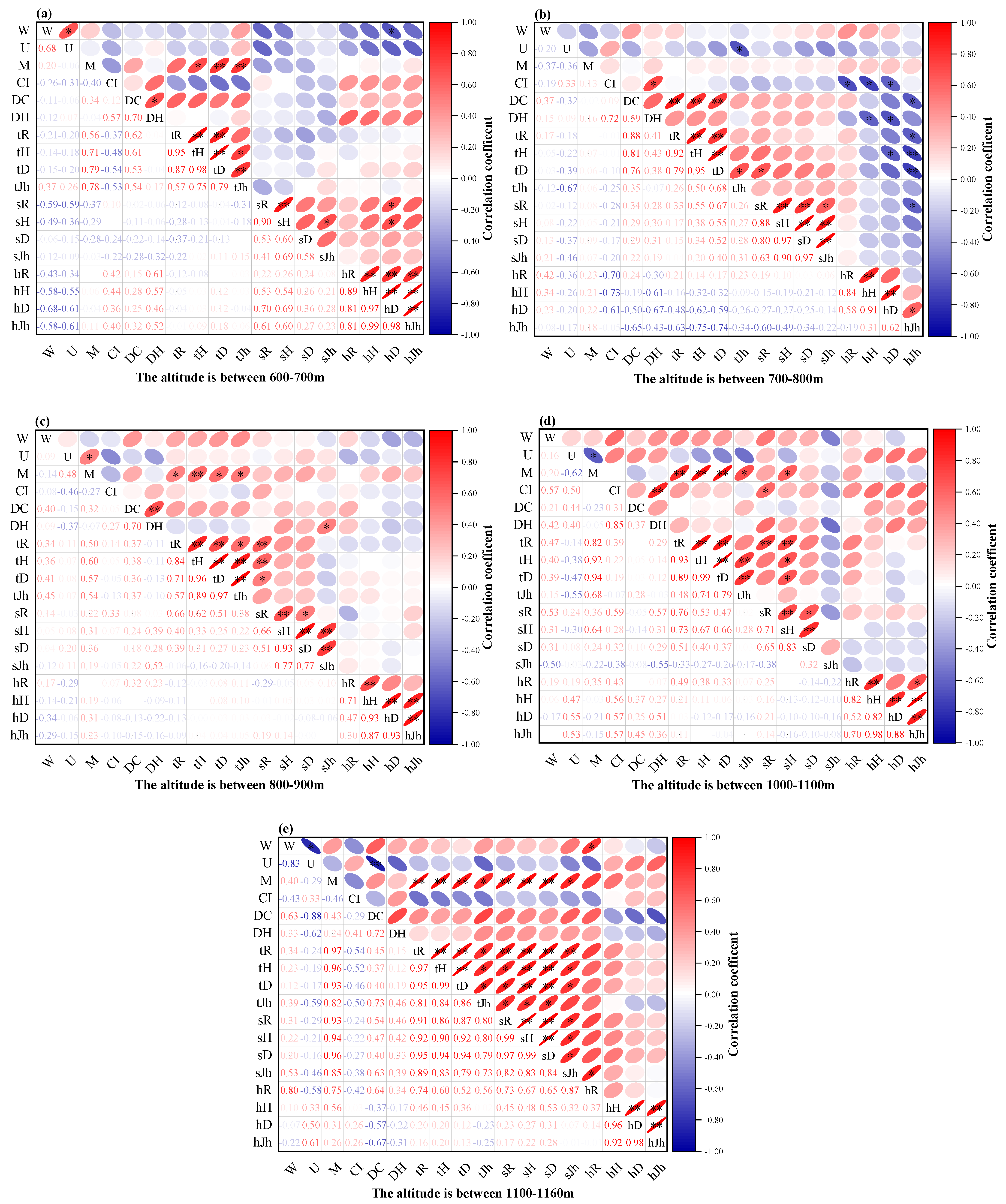
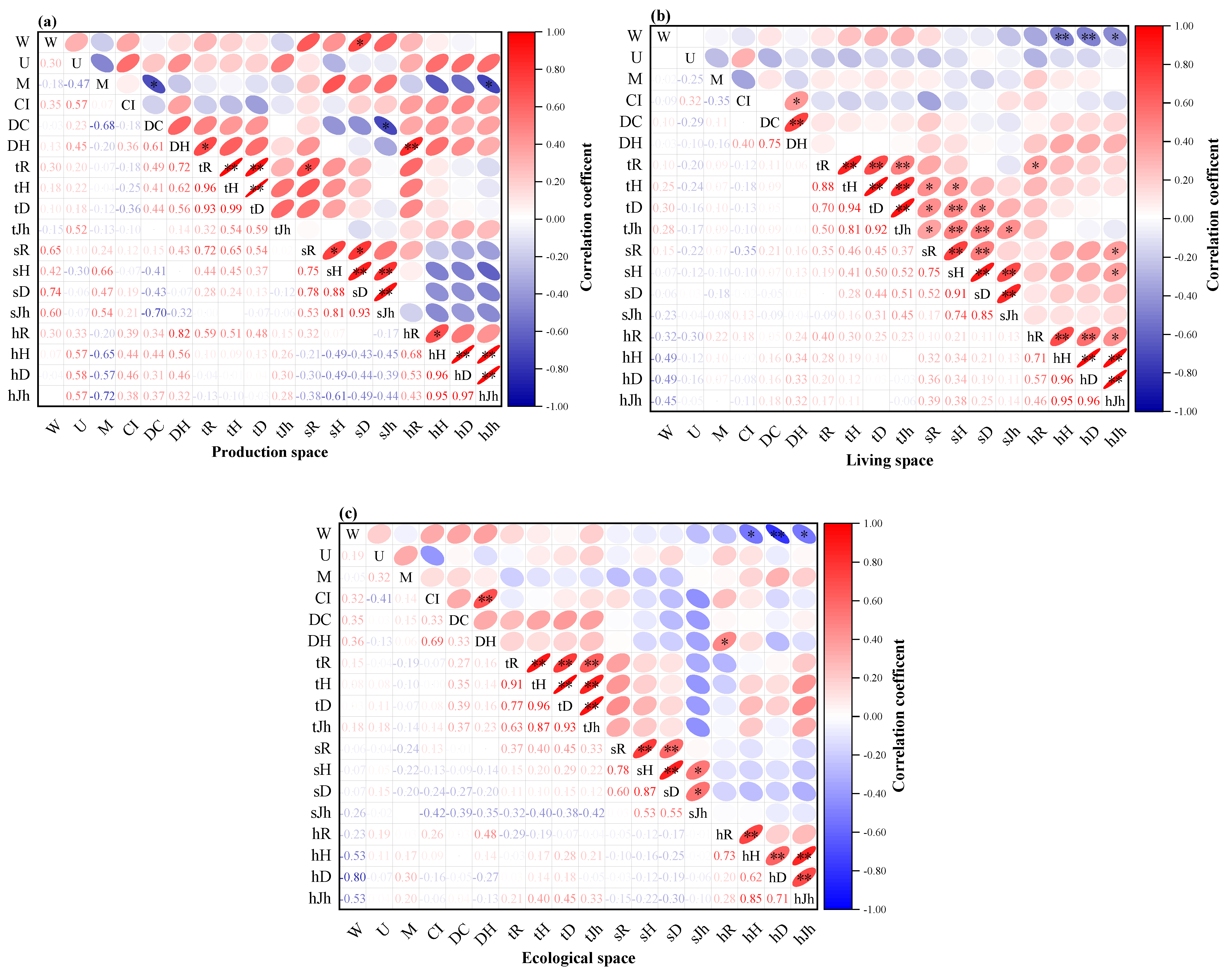
3.3. Correlation between Plant Community Structure and Species Diversity in Traditional Villages in Karst Topographic Regions of the ZUNYI City
3.3.1. Relationship between Plant Community Structure and Species Diversity in Traditional Villages in Karst Topographic Regions of the Zunyi City as a Whole
3.3.2. Relationship between Plant Community Structure and Species Diversity in Traditional Villages at Different Altitude Gradients
3.3.3. Relationship between Plant Community Structure and Species Diversity under the “Production-Living-Ecological” Spaces of Traditional Villages
4. Discussion
4.1. Plant Community Structure and Species Diversity Characteristics of Traditional Villages in Karst Topographic Regions of the Zunyi
4.2. Characteristics of the Relationship between Plant Community Structure and Species Diversity in Traditional Villages in Karst Topographic Regions of the Zunyi City and Development Strategies
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Feng, J. The Dilemma and Development Path of Traditional Villages: Traditional Villages are Another Type of Cultural Heritage. Folk Cult. Forum 2013, 6, 7–12. [Google Scholar] [CrossRef]
- Gao, J.; Wu, B. Revitalizing traditional villages through rural tourism: A case study of Yuanjia Village, Shaanxi Province, China. Tour. Manag. 2017, 63, 223–233. [Google Scholar] [CrossRef]
- Chen, B.; Yao, Z.; Zhang, H.; Norv, C. Research on landscape guided rural revitalization planning and design. Chin. Gard. 2020, 36, 58–62. [Google Scholar] [CrossRef]
- Mok, D.; Guan, D.S.; Huang, K.Y.; Liu, S.W. Typical plant community structure and species diversity in an ecological safety island in urban Guangzhou. J. Ecol. 2011, 31, 1515–1524. [Google Scholar]
- Li, Y.; Ye, S.; Hui, G.; Hu, Y.; Zhao, Z. Spatial structure of timber harvested according to structure-based forest management. For. Ecol. Manag. 2014, 322, 106–116. [Google Scholar] [CrossRef]
- Zhao, Z.; Hui, G.; Hu, Y.; Li, Y.; Li, Y. Application of structure-based forest management in broadleaved Korean pine mixed forest. For. Res. 2013, 26, 467–472. [Google Scholar]
- Qiu, J.; Tang, M.; Shen, L. Dynamic analysis of spatial structure in a close to nature Phyllostachys edulis stands. Acta Ecol. Sin. 2014, 34, 1444–1450. [Google Scholar]
- Delang, C.O.; Li, W.M. Species Richness and Diversity. In Ecological Succession on Fallowed Shifting Cultivation Fields; Springer: Dordrecht, The Netherlands, 2013; Volume 978, pp. 39–66. [Google Scholar]
- Xie, Z.; Chang, L.; Scheu, S.; Wu, D.; Sun, X. Taxonomic and functional diversity of Collembola in litter and soil along an altitudinal gradient at Changbai Mountain, China. Acta Ecol. Sin. 2022, 42, 3471–3481. [Google Scholar] [CrossRef]
- Zhang, W.; Xue, W.; Yang, B.; Zhao, Z. Differences in community characteristics, species diversity, and their coupling associations among three Quercus acutissima forests in Qiaoshan. Acta Ecol. Sin. 2019, 39, 3991–4001. [Google Scholar] [CrossRef]
- Tao, W.; Chen, H.; Lin, J. Spatial Form and Spatial Cognition of Traditional Village in Syntactical View: A Case Study of Xiaozhou Village, Guangzhou. Acta Geogr. Sin. 2013, 68, 209–218. [Google Scholar] [CrossRef]
- Bański, J. Transformations in Housing Construction in Rural Areas of Poland’s Lublin Region—Influence on the Spatial Set-tlement Structure and Landscape Aesthetics. Landsc. Urban Plan. 2010, 94, 116–126. [Google Scholar] [CrossRef]
- Li, K.; Zhang, G. Species Diversity and Distribution Pattern of Heritage Trees in the Rapidly-Urbanizing Province of Jiangsu, China. Forests 2021, 12, 1543. [Google Scholar] [CrossRef]
- Saiz, H.; Gómez-Gardeñes, J.; Borda, J.P.; Maestre, F.T. The structure of plant spatial association networks is linked to plant diversity in global drylands. J. Ecol. 2018, 106, 1443–1453. [Google Scholar] [CrossRef] [PubMed]
- Lurgi, M.; Montoya, D.; Montoya, J.M. The effects of space and diversity of interaction types on the stability of complex ecological networks. Theor. Ecol. 2016, 9, 3–13. [Google Scholar] [CrossRef]
- Zhang, R.; Shen, H.; Dong, S.; Li, S.; Xiao, J.; Zhi, Y.; Zhang, J.; Zuo, H.; Wu, S.; Mu, Z.; et al. Effects of 5-Year Nitrogen Addition on Species Composition and Diversity of an Alpine Steppe Plant Community on Qinghai-Tibetan Plateau. Plants 2022, 11, 966. [Google Scholar] [CrossRef]
- Yang, Y.; Fujihara, M.; Li, B.; Yuan, X.; Hara, K.; Da, L.; Tomita, M.; Zhao, Y. Structure and diversity of remnant natural evergreen broad-leaved forests at three sites affected by urbanization in Chongqing metropolis, Southwest China. Landsc. Ecol. Eng. 2011, 10, 137–149. [Google Scholar] [CrossRef]
- Gadow, K.V.; Zhang, C.Y.; Wehenkel, C.; Pommerening, A.; Corral-Rivas, J.; Korol, M.; Myklush, S.; Hui, G.Y.; Kiviste, A.; Zhao, X.H. Forest Structure and Diversity. In Continuous Cover Forestry. Managing Forest Ecosystems; Pukkala, T., von Gadow, K., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; Volume 23, pp. 29–83. [Google Scholar] [CrossRef]
- Ge, B.; Dou, Q.; Zhang, R. Study on the characteristics of seed flora of Zunyi City. Seeds 2014, 33, 53–55+59. [Google Scholar] [CrossRef]
- Liu, J.; Liu, Y.; Li, Y. Classification evaluation and spatial-temporal analysis of “production-living-ecological” spaces in China. J. Geogr. 2017, 72, 1290–1304. [Google Scholar] [CrossRef]
- Wang, G.-H.; Fang, J.-Y.; Guo, K.; Xie, Z.-Q.; Tang, Z.-Y.; Shen, Z.-H.; Wang, R.-Q.; Wang, X.-P.; Wang, D.-L.; Qiang, S.; et al. Contents and protocols for the classification and description of Vegetation Formations, Alliances and Associations of vegetation of China. J. Plant Ecol. 2020, 44, 128–178. [Google Scholar] [CrossRef]
- Chytrý, M.; Schaminée, J.H.J.; Schwabe, A. Vegetation survey: A new focus for Applied Vegetation Science. Appl. Veg. Sci. 2011, 14, 435–439. [Google Scholar] [CrossRef]
- Kouba, Y.; Martínez-García, F.; De Frutos, Á.; Alados, C.L. Plant β-diversity in human-altered forest ecosystems: The importance of the structural, spatial, and topographical characteristics of stands in patterning plant species assemblages. Eur. J. For. Res. 2014, 133, 1057–1072. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Hui, G.; Yu, S.; Luo, Y.; Yao, X.; Ye, S. Nearest neighbour relationships in Pinus yunnanensis var. tenuifolia forests along the Nanpan River, China. iFor.—Biogeosci. For. 2017, 10, 746–753. [Google Scholar] [CrossRef]
- Gadow, K.V. Zur bestandesbeschreibung in der forsteinrichtung. Forst und Holz 1993, 48, 602–606. [Google Scholar]
- Wei, H.; Dong, L.; Liu, Z. Simulation of optimal spatial structure of forest stands in major forest types in the Daxinganling Mountains. J. Appl. Ecol. 2019, 30, 3824–3832. [Google Scholar] [CrossRef]
- Wang, W.; Xing, Z.; Li, W.; Yang, X. Study on Diversity of Undergrowth Plant Community in Cibagou Nature Reserve. Am. J. Plant Sci. 2017, 08, 2149–2158. [Google Scholar] [CrossRef] [Green Version]
- Kurttila, M. The spatial structure of forests in the optimization calculations of forest planning—A landscape ecological perspective. For. Ecol. Manag. 2001, 142, 129–142. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, L.; Guo, Q.; Nie, D.; Bai, X.; Jiang, Y. Trends in the distribution of dominant tree species in forest communities at the top of succession. J. Plant Ecol. 1999, 65–77. [Google Scholar] [CrossRef]
- Chen, T.; Xu, F.; Yang, Q.; Chen, S.; Ge, X.; Wu, J.; Cui, P.; Fang, Y.; Ding, H. Spatial distribution characteristics of an evergreen broad-leaved forest in the Wuyi Mountains, Fujian Province, southeastern China. J. Ecol. 2018, 38, 1817–1825. [Google Scholar] [CrossRef]
- Zhang, G.; Wang, D.; Chai, Z.; Zhang, C.; Liu, W. Distribution characteristics of spatial structure parameters of 2 typical natural forests in Xiaolong Mountains. For. Sci. Res. 2015, 28, 531–537. [Google Scholar] [CrossRef]
- Homeier, J.; Breckle, S.-W.; Günter, S.; Rollenbeck, R.T.; Leuschner, C. Tree Diversity, Forest Structure and Productivity along Altitudinal and Topographical Gradients in a Species-Rich Ecuadorian Montane Rain Forest. Biotropica 2010, 42, 140–148. [Google Scholar] [CrossRef]
- Andreas, M.; Markus, B.; Wolfgang, S. Herb-layer diversity in deciduous forests: Raised by tree richness or beaten by beech? For. Ecol. Manag. 2008, 256, 272–281. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, W.; Du, H.; Zhong, Z.; Xiao, L.; Zhou, W.; Wang, H. Differences in community characteristics, species diversity, and their coupling associations among three forest types in the Huzhong area, Daxinganling mountains. Acta Ecol. Sin. 2018, 38, 4684–4693. [Google Scholar] [CrossRef]
- Cao, N.; Xue, D. On biodiversity conservation by Zhuang traditional culture: A case study in Jingxi City of Guangxi Zhuang Autonomous Region. Biodiver. Sci. 2019, 27, 728–734. (In Chinese) [Google Scholar]
- Latt, M.M.; Park, B.B. Tree Species Composition and Forest Community Types along Environmental Gradients in Htamanthi Wildlife Sanctuary, Myanmar: Implications for Action Prioritization in Conservation. Plants 2022, 11, 2180. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, L.; Chen, J.; Feng, L.; Li, F.; Yu, L. Functional Diversity of Plant Communities in Relationship to Leaf and Soil Stoichiometry in Karst Areas of Southwest China. Forests 2022, 13, 864. [Google Scholar] [CrossRef]
- Rahbek, C. The role of spatial scale and the perception of large-scale species-richness patterns. Ecol. Lett. 2005, 8, 224–239. [Google Scholar] [CrossRef]
- Worthy, S.J.; Paz, R.A.J.; Pérez, J.; Reynolds, A.; Cruse-Sanders, J.; Valencia, R.; Barone, J.A.; Burgess, K.S. Distribution and Community Assembly of Trees Along an Andean Elevational Gradient. Plants 2019, 8, 326. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Hu, C.; Wang, Z. Ecological Response of Urban Forest Carbon Density to Site Conditions: A Case Study of a Typical Karst Mountainous Regions in Southwest China. Forests 2022, 13, 1484. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, H.Y.H.; Taylor, A. Multiple drivers of plant diversity in forest ecosystems. Glob. Ecol. Biogeogr. 2014, 23, 885–893. [Google Scholar] [CrossRef]
- Alexander, J.M.; Kueffer, C.; Daehler, C.; Edwards, P.J.; Pauchard, A.; Seipel, T.; Arévalo, R.J.; Cavieres, L.A.; Dietz, H.; Jakobs, G.; et al. Assembly of nonnative floras along elevational gradients explained by directional ecological filtering. Proc. Natl. Acad. Sci. USA 2011, 108, 656–661. [Google Scholar] [CrossRef] [Green Version]
- Coomes, D.A.; Allen, R.B. Effects of size, competition and altitude on tree growth. J. Ecol. 2007, 95, 1084–1097. [Google Scholar] [CrossRef]
- Zhao, W.; Liu, X.; Jing, W.; Xu, L.; Niu, Y.; Qi, P.; Zhao, Y. Spatial heterogeneity in the community structure of Qinghai spruce forests in Qilian Mountains. J. Appl. Ecol. 2015, 26, 2591–2599. [Google Scholar] [CrossRef]
- MacFadyen, E.D.A. The Self-Thinning Rule. In Advances in Ecological Research; Academic Press: Cambridge, MA, USA, 1984; Volume 14, pp. 167–225. [Google Scholar] [CrossRef]
- Ye, C.; An, M.; Zhang, N.; Chen, L. Effects of Dodonaea viscosa afforestation on native plant species diversity in the Beipanjiang Karst region of Guizhou. NW. J. Bot. 2019, 39, 310–318. [Google Scholar]
- Xu, Z. Conservation of biodiversity and cultural diversity are two sides of a coin: Xishuangbanna Dai’s ecological culture as an example. Biodivers. Sci. 2015, 23, 126–130. [Google Scholar] [CrossRef]
- Ye, C.; An, M.; Zhang, N.; Chen, L. Quantifying stand structural complexity and its relationship with forest management, tree species diversity and microclimate. Agric. For. Meteorol. 2017, 242, 1–9. [Google Scholar] [CrossRef]
- Bartels, S.F.; Chen, H.Y. Interactions between overstorey and understorey vegetation along an overstorey compositional gradient. J. Veg. Sci. 2013, 24, 543–552. [Google Scholar] [CrossRef]
- Bian, G.; Zhu, G. Effects of Spatial Structure of Quercus on Species Diversity of Undergrowth Shrubs in Badagong Mountain. Chin. Agron. Bull. 2020, 36, 39–43. [Google Scholar] [CrossRef]
- Peet, R.K. The Measurement of Species Diversity. Annu. Rev. Ecol. Syst. 1974, 5, 285–307. [Google Scholar] [CrossRef]
- Zhu, J.; Zhan, B.; Chen, X.; Guan, Q. Spatial structure characteristics and plant diversity analysis of mixed coniferous and broad forests in Zijinshan. J. Cent. South Univ. For. Sci. Technol. 2018, 38, 75–80+93. [Google Scholar] [CrossRef]
- Li, F.; Pan, P.; Ning, J.K. Influence of spatial structure of fly-sown horsetail pine forest stands on the diversity of understory vegetation. J. Northeast. For. Univ. 2016, 40, 31–35. [Google Scholar] [CrossRef]
- Austrheim, G.; Eriksson, O. Plant species diversity and grazing in the Scandinavian mountains—Patterns and processes at different spatial scales. Ecography 2001, 24, 683–695. [Google Scholar] [CrossRef]
- Wright, D.H. Species- Energy Theory: An Extension of Species-Area Theory (1983). In Foundations of Macroecology: Classic Papers with Commentaries; Smith, F., Gittleman, J., Brown, J., Eds.; University of Chicago Press: Chicago, IL, USA, 2014; pp. 593–605. [Google Scholar] [CrossRef]
- Davies, R.G.; Orme, C.D.L.; Storch, D.; Olson, V.A.; Thomas, G.; Ross, S.G.; Ding, T.-S.; Rasmussen, P.C.; Bennett, P.M.; Owens, I.; et al. Topography, energy and the global distribution of bird species richness. Proc. R. Soc. B Boil. Sci. 2007, 274, 1189–1197. [Google Scholar] [CrossRef] [PubMed]
- Perea, R.; Schroeder, J.W.; Dirzo, R. The Herbaceous Understory Plant Community in the Context of the Overstory: An Overlooked Component of Tropical Diversity. Diversity 2022, 14, 800. [Google Scholar] [CrossRef]
- Barbier, S.; Gosselin, F.; Balandier, P. Influence of tree species on understory vegetation diversity and mechanisms involved—A critical review for temperate and boreal forests. For. Ecol. Manag. 2008, 254, 38. [Google Scholar] [CrossRef]
- Liu, Q.; Deng, H.; Li, Z.; Liang, S.; Li, Q.; Ni, D. Characteristics of plant communities in the Guizhou Chishui Cyathea National Nature Reserve. J. Beijing For. Univ. 2019, 41, 19–3110. [Google Scholar] [CrossRef]
- Aussenac, R.; Bergeron, Y.; Gravel, M.; Drobyshev, I. Interactions among trees: A key element in the stabilising effect of species diversity on forest growth. Funct. Ecol. 2019, 33, 360–367. [Google Scholar] [CrossRef]
- Deng, N.; Song, Q.; Ma, F.; Tian, Y. Patterns and Driving Factors of Diversity in the Shrub Community in Central and Southern China. Forests 2022, 13, 1090. [Google Scholar] [CrossRef]
- Zhu, G.; Xu, Q.; Lv, Y. Effects of stand spatial structure on species diversity of shrubs in Quercus spp. natural secondary forests in Hunan Province. Acta Ecol. Sin. 2018, 38, 5404–5412. [Google Scholar] [CrossRef]
- Xiang, Q.; Guo, Q.; Ai, X.; Yao, L.; Zhu, J.; Xue, Z.; Zhou, Y.; Zhao, H.; Wu, J. Spatial structure of forest stands and the pattern of species diversity with spatial scale. For. Sci. Res. 2022, 35, 151–160. [Google Scholar] [CrossRef]
- Benthien, O.; Braun, M.; Riemann, J.C.; Stolter, C. Long-term effect of sheep and goat grazing on plant diversity in a semi-natural dry grassland habitat. Heliyon 2018, 4, e00556. [Google Scholar] [CrossRef] [Green Version]
- Shao, D.X.H. Influence of traditional Wa culture on vegetable germplasm diversity in Yunnan Province. Biodiversity 2017, 29, 1120–1127. [Google Scholar] [CrossRef] [Green Version]
- Niemelä, J.; Saarela, S.-R.; Söderman, T.; Kopperoinen, L.; Yli-Pelkonen, V.; Väre, S.; Kotze, D.J. Using the ecosystem services approach for better planning and conservation of urban green spaces: A Finland case study. Biodivers. Conserv. 2010, 19, 3225–3243. [Google Scholar] [CrossRef]
- Chen, M.; Hu, G.; Hu, Y.; Zhang, G.; Liu, R.; Yang, A.; Zhao, Z.; Wang, H. Effects of structured forest management on forest quality of broadleaf red pine forests in Northeast China. J. Beijing For. Univ. 2019, 41, 19–30. [Google Scholar] [CrossRef]
- Chen, Y.; Cao, Y. Response of tree regeneration and understory plant species diversity to stand density in mature Pinus tabulaeformis plantations in the hilly area of the Loess Plateau, China. Ecol. Eng. 2014, 73, 238–245. [Google Scholar] [CrossRef]







| Village Name | Longitude and Latitude | Nation | Area (hm2) | Population | Representative Features |
|---|---|---|---|---|---|
| Longtan | 28°36′42″ N, 107°57′51″ E | Gelao, Miao nationality | 31.71 | 436 | It was one of the first Chinese traditional village. The natural pattern is “living based on the mountain, with the stone forest reflecting the mask”. |
| Sankeng | 28°36′09″ N, 108°00′06″ E | Gelao, Miao nationality | 0.67 | 560 | It is the fifth batch of traditional Chinese villages. A long history and a profound cultural heritage. Most of the ancient wooden structure houses are well preserved. |
| Manao | 28°06′51″ N, 107°44′40″ E | Han, Gelao nationality | 19.99 | 148 | In December 2012, Manao Village was included in the first batch of Chinese traditional village list. |
| Heixi | 28°16′19″ N, 107°42′59″ E | Han, Gelao, Tujia nationality | 0.40 | 4868 | In November 2014, Heixi Ancient Village was selected as the third batch of traditional villages in China, and the ancient buildings are relatively well preserved. |
| Shijiazhai | 28°06′36″ N, 107°25′42″ E | Han, Gelao nationality | 3.99 | 3552 | It is the second batch of traditional Chinese villages. Preserves the people’s commune buildings and large areas of traditional residential buildings. |
| Gouba | 27°38′19″ N, 106°34′07″ E | Han, Gelao nationality | 16.70 | 3412 | It is the third batch of traditional Chinese villages. There are famous scenic spots such as Gouba Conference site, Gouba Conference Exhibition Hall, ingenuity garden, “Dream 1935” light and shadow show. |
| Altitude (m) | Summation | Longtan | Sankeng | Manao | Heixi | Shijiazhai | Gouba |
|---|---|---|---|---|---|---|---|
| 600–700 | 10 | 10 | —— | —— | —— | —— | —— |
| 700–800 | 10 | —— | —— | —— | 5 | 5 | —— |
| 800–900 | 18 | —— | —— | 9 | 5 | 4 | —— |
| 1000–1100 | 12 | —— | 7 | —— | —— | —— | 5 |
| 1100–1160 | 7 | —— | 3 | —— | —— | —— | 4 |
| PLE Spaces | The Type | The Number | Longtan | Sankeng | Manao | Heixi | Shijiazhai | Gouba |
|---|---|---|---|---|---|---|---|---|
| 57 | 10 | 9 | 9 | 11 | 9 | 9 | ||
| Produce | Vegetable field | 6 | 1 | 2 | —— | 1 | 1 | 1 |
| Tea garden | 2 | —— | —— | 1 | —— | 1 | —— | |
| Farmland | 2 | —— | 1 | —— | —— | 1 | —— | |
| Fruit forest | 2 | 1 | —— | —— | —— | —— | 1 | |
| Living | Courtyard | 6 | 1 | 1 | 1 | 1 | 1 | 1 |
| House side | 7 | 1 | 1 | 1 | 2 | 1 | 1 | |
| Roads | 7 | 1 | 1 | 2 | 1 | 2 | ||
| Square | 2 | 1 | —— | —— | 1 | —— | —— | |
| Sacrifice | 5 | 1 | 1 | 1 | 1 | 1 | —— | |
| Ecology | Forest | 12 | 3 | 1 | 4 | 2 | 2 | 1 |
| River channel | 6 | 1 | 1 | 2 | 1 | 1 |
| Index | Formula | Value | Index Meaning | ||||
|---|---|---|---|---|---|---|---|
| 0.00 | 0.25 | 0.50 | 0.75 | 1.00 | |||
| Uniform angle index (Wi) | very regular | regular | random | clumped | very clumped | Wi, i is the core wood, j is the adjacent wood. When the jth angle is smaller than the standard angle (α0 = 72°), Zij = 1, but not Zij = 0. Wi = 0, very regular distribution. Wi = 0.25, regular distribution. Wi = 0.50, random distribution. Wi = 0.75, clumped distribution. Wi = 1.00, very clumped distribution. | |
| Dominance (Ui) | predominant | sub-dominant | medium | disadvantaged | Absolutely disadvantaged | Ui, when the core wood i is more adjacent than the j strain. Kij = 1, and vice versa, Kij = 0. Ui = 0, in the advantage. Ui = 0.25, which is the sub-advantage. Ui = 0.50, in Medium. Ui = 0.75, in the survival of the fittest. Ui = 1.00, in the absolute survival of the fittest. | |
| Mingling (Mi) | no mingling | weak mingling | moderate mingling | high mingling | very high mingling | Mi, when core wood i is not adjacent to the j th strain. Vij = 1, and vice versa, Vij = 0. Mi = 0, no mingling. Mi = 0.25, weak degree mingling. Mi = 0.50, moderate mingling. Mi = 0.75, high mingling. Mi = 1.00, very high mingling. | |
| Competition Index (CIi) | —— | —— | —— | —— | —— | Competition index CIi describing the size of forest tree competitiveness. Dij represents the chest diameter of adjacent wood j, and Di indicates the chest diameter of core wood i, DISij, it ote the European distance between core wood i and adjacent wood j. | |
| Growth Vitality (DCi) | —— | —— | —— | —— | —— | The DCi that determines the growth and vitality of forest trees. CWi and Di, the crown length and chest diameter of the core wood i, respectively. | |
| Stability (DHi) | —— | —— | —— | —— | —— | The DHi that reflects the tree stability, Hi and Di, tree height and chest diameter of the core wood i, respectively. | |
| Distribution Level | The Margalef Index (R) | The Shannon-Wiener Index (H) | The Simpson Index (D) | The Pielou Index (J) |
|---|---|---|---|---|
| Tree layer | 2.330 ± 1.101 | 0.746 ± 0.272 | 0.720 ± 0.195 | 0.345 ± 0.067 |
| Shrub layer | 2.510 ± 1.196 | 0.830 ± 0.252 | 0.753 ± 0.158 | 0.350 ± 0.071 |
| Herb layer | 4.151 ± 1.122 | 1.083 ± 0.285 | 0.820 ± 0.173 | 0.324 ± 0.077 |
| average value | 2.996 ± 1.401 | 0.886 ± 0.304 | 0.764 ± 0.180 | 0.340 ± 0.072 |
| Altitude Gradients (m) | The Margalef Index (R) | The Shannon-Wiener Index (H) | The Simpson Index (D) | The Pielou Index (J) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Tree Layer | Shrub Layer | Herb Layer | Tree Layer | Shrub Layer | Herb Layer | Tree Layer | Shrub Layer | Herb Layer | Tree Layer | Shrub Layer | Herb Layer | |
| 600–700 | 1.811 ± 0.840a | 2.500 ± 1.334ab | 4.000 ± 1.200a | 0.643 ± 0.240a | 0.832 ± 0.272a | 0.930 ± 0.295a | 0.671 ± 0.190ab | 0.770 ± 0.113a | 0.723 ± 0.180a | 0.340 ± 0.055ab | 0.360 ± 0.051a | 0.280 ± 0.080a |
| 700–800 | 2.910 ± 1.393b | 2.610 ± 0.900ab | 4.330 ± 1.444a | 0.873 ± 0.264b | 0.810 ± 0.263a | 1.182 ± 0.273b | 0.804 ± 0.120b | 0.726 ± 0.220a | 0.870 ± 0.161b | 0.376 ± 0.033a | 0.310 ± 0.091a | 0.360 ± 0.067b |
| 800–900 | 2.900 ± 2.840b | 2.780 ± 1.323a | 4.390 ± 1.076a | 0.865 ± 0.200b | 0.880 ± 0.260a | 1.200 ± 1.180b | 0.800 ± 0.140b | 0.764 ± 0.164a | 0.884 ± 0.076b | 0.356 ± 0.054ab | 0.350 ± 0.085a | 0.355 ± 0.044b |
| 1000–1100 | 1.640 ± 0.860a | 1.900 ± 1.070b | 3.880 ± 0.901a | 0.560 ± 0.300a | 0.730 ± 0.233a | 1.030 ± 0.320ab | 0.572 ± 0.270a | 0.704 ± 0.151a | 0.800 ± 0.193ab | 0.315 ± 0.112b | 0.350 ± 0.058a | 0.309 ± 0.086ab |
| 1100–1160 | 2.300 ± 0.683ab | 2.900 ± 1.030ab | 4.124 ± 0.900a | 0.820 ± 0.190b | 0.950 ± 0.181a | 1.113 ± 0.170ab | 0.800 ± 0.123b | 0.840 ± 0.080a | 0.855 ± 0.076ab | 0.370 ± 0.035ab | 0.374 ± 0.012a | 0.330 ± 0.051ab |
| “PLE Spaces” | The Margalef Index (R) | The Shannon-Wiener Index (H) | The Simpson Index (D) | The Pielou Index (J) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Tree Layer | Shrub Layer | Herb Layer | Tree Layer | Shrub Layer | Herb Layer | Tree Layer | Shrub Layer | Herb Layer | Tree Layer | Shrub Layer | Herb Layer | |
| Produce | 1.540 ± 0.740a | 1.437 ± 0.656a | 3.806 ± 1.330a | 0.580 ± 0.246a | 0.664 ± 0.232a | 0.858 ± 0.370a | 0.650 ± 0.198a | 0.680 ± 0.204a | 0.658 ± 0.253a | 0.377 ± 0.068a | 0.350 ± 0.088a | 0.256 ± 0.103a |
| Living | 2.730 ± 0.900b | 2.803 ± 1.089b | 4.430 ± 1.150a | 0.840 ± 0.223b | 0.882 ± 0.224b | 0.171 ± 0.245b | 0.780 ± 0.144b | 0.791 ± 0.120b | 0.862 ± 0.25b | 0.360 ± 0.051a | 0.346 ± 0.057a | 0.345 ± 0.062b |
| Ecology | 2.260 ± 1.315b | 2.815 ± 1.257b | 3.950 ± 0.811a | 0.720 ± 0.307ab | 0.866 ± 0.268b | 1.105 ± 0.180b | 0.676 ± 0.242ab | 0.746 ± 0.167ab | 0.870 ± 0.080b | 0.313 ± 0.080b | 0.340 ± 0.082a | 0.340 ± 0.045b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, C.; Huang, Z.; Zhang, X.; Zhao, H.; Jiang, S.; Ren, Y. Correlation between Vegetation Structure and Species Diversity in Traditional Villages in Karst Topographic Regions of the Zunyi City, China. Plants 2022, 11, 3161. https://doi.org/10.3390/plants11223161
Deng C, Huang Z, Zhang X, Zhao H, Jiang S, Ren Y. Correlation between Vegetation Structure and Species Diversity in Traditional Villages in Karst Topographic Regions of the Zunyi City, China. Plants. 2022; 11(22):3161. https://doi.org/10.3390/plants11223161
Chicago/Turabian StyleDeng, Caijie, Zongsheng Huang, Xiaojing Zhang, Hua Zhao, Siyu Jiang, and Yuxin Ren. 2022. "Correlation between Vegetation Structure and Species Diversity in Traditional Villages in Karst Topographic Regions of the Zunyi City, China" Plants 11, no. 22: 3161. https://doi.org/10.3390/plants11223161