
Hydroethanolic Extract of Morus nigra L. Leaves: A Dual PPAR-α/γ Agonist with Anti-Inflammatory Properties in Lipopolysaccharide-Stimulated RAW 264.7

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
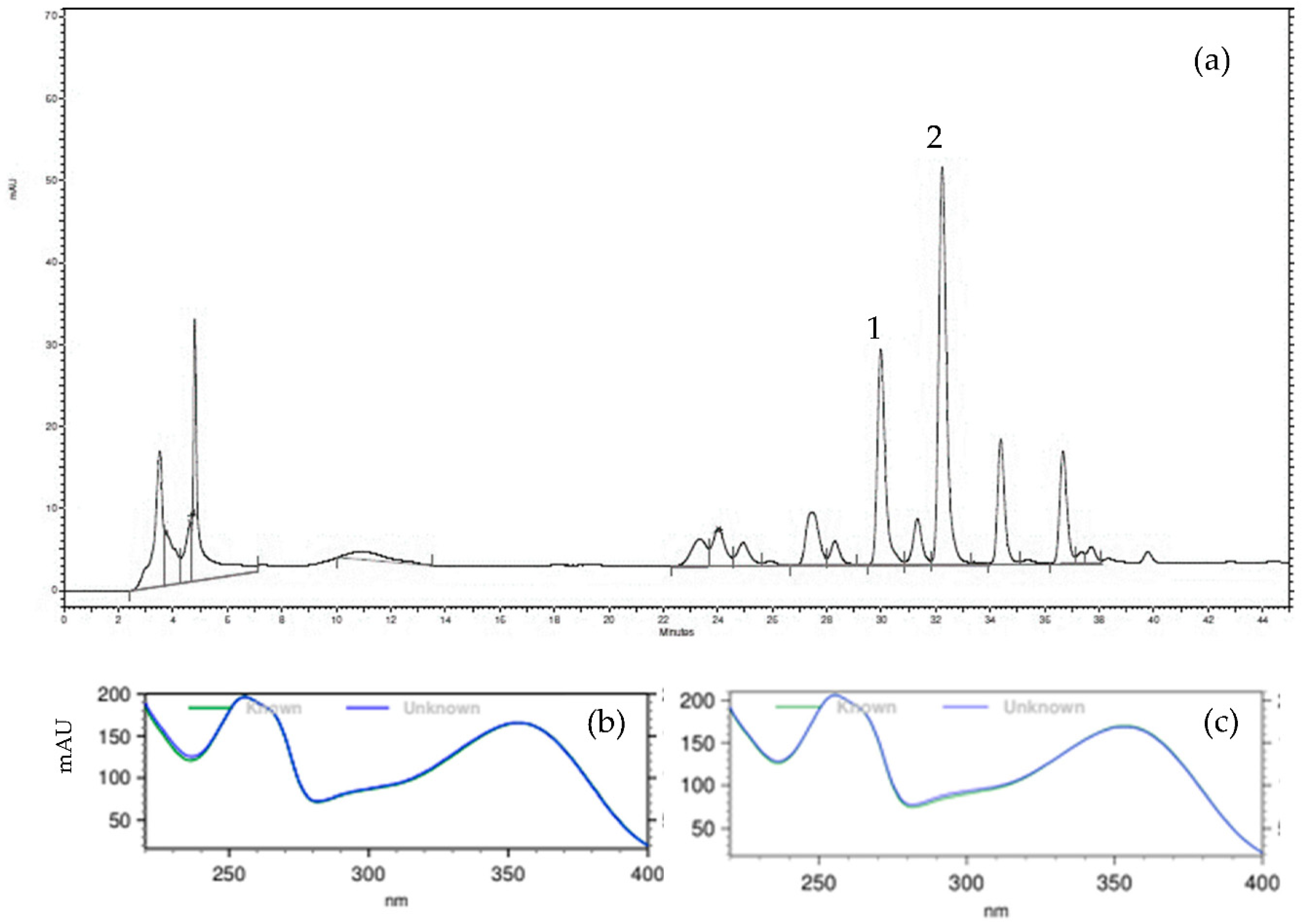
2.1. Chemical Profile
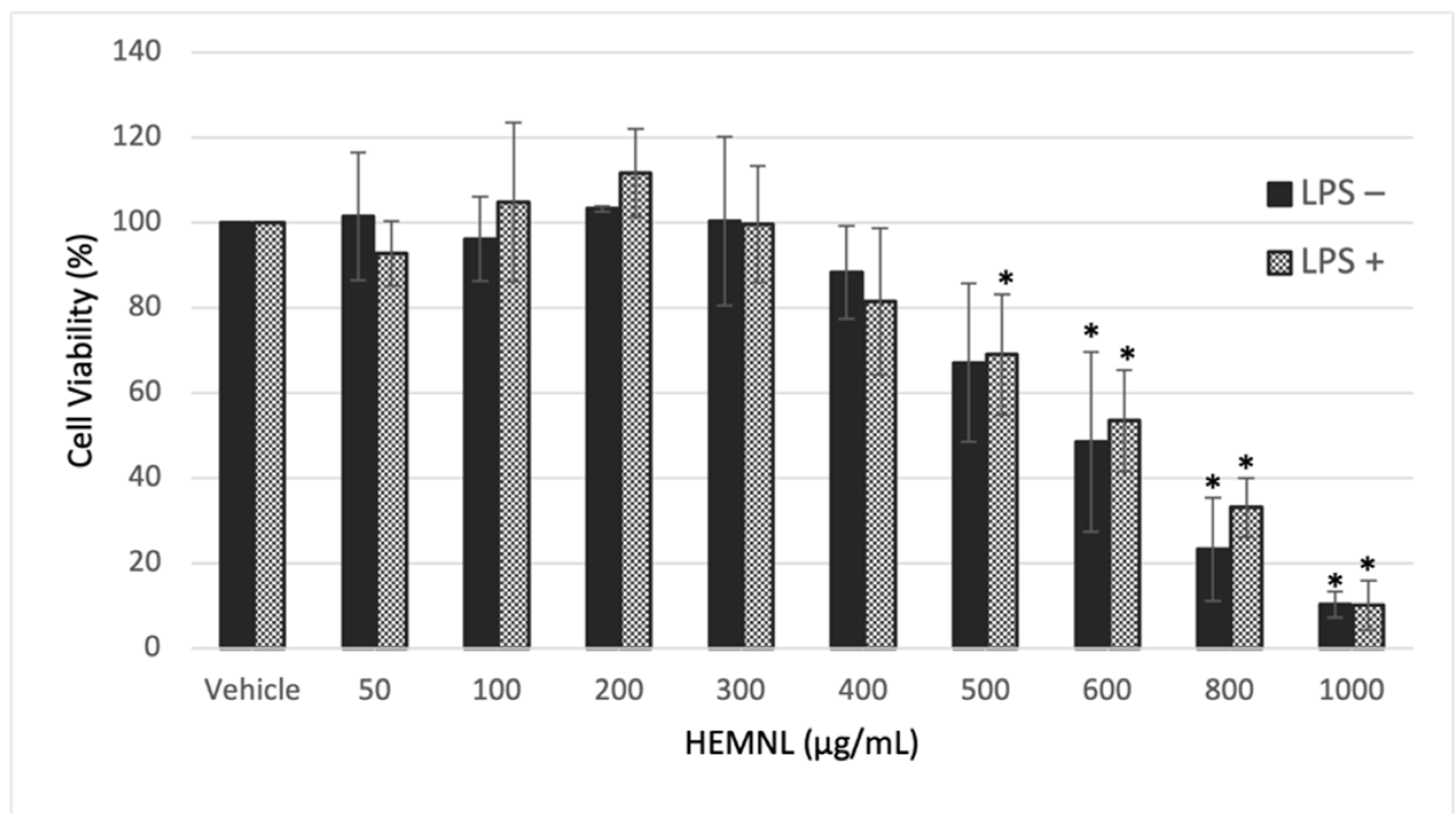
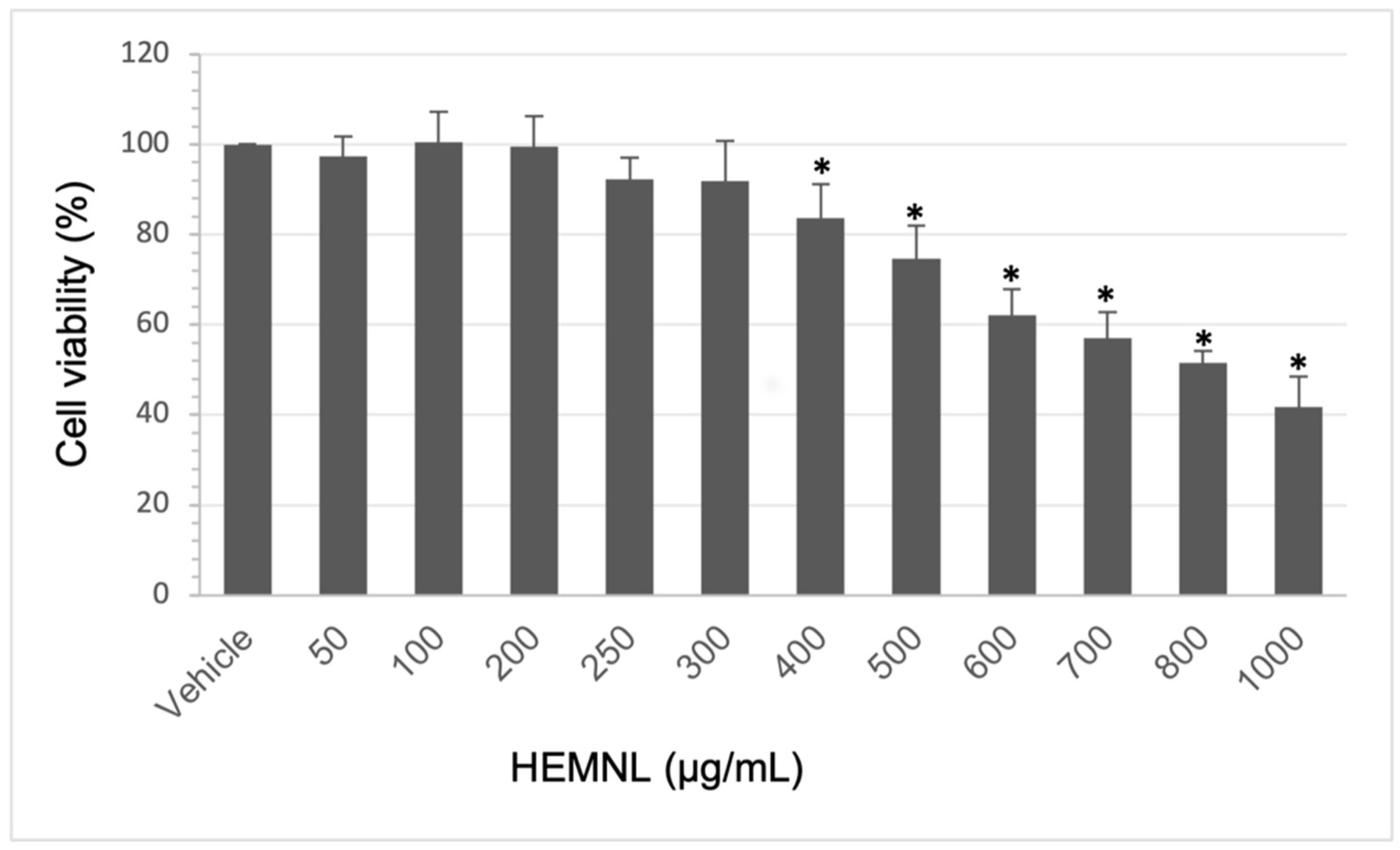
2.2. Extract Cytotoxicity
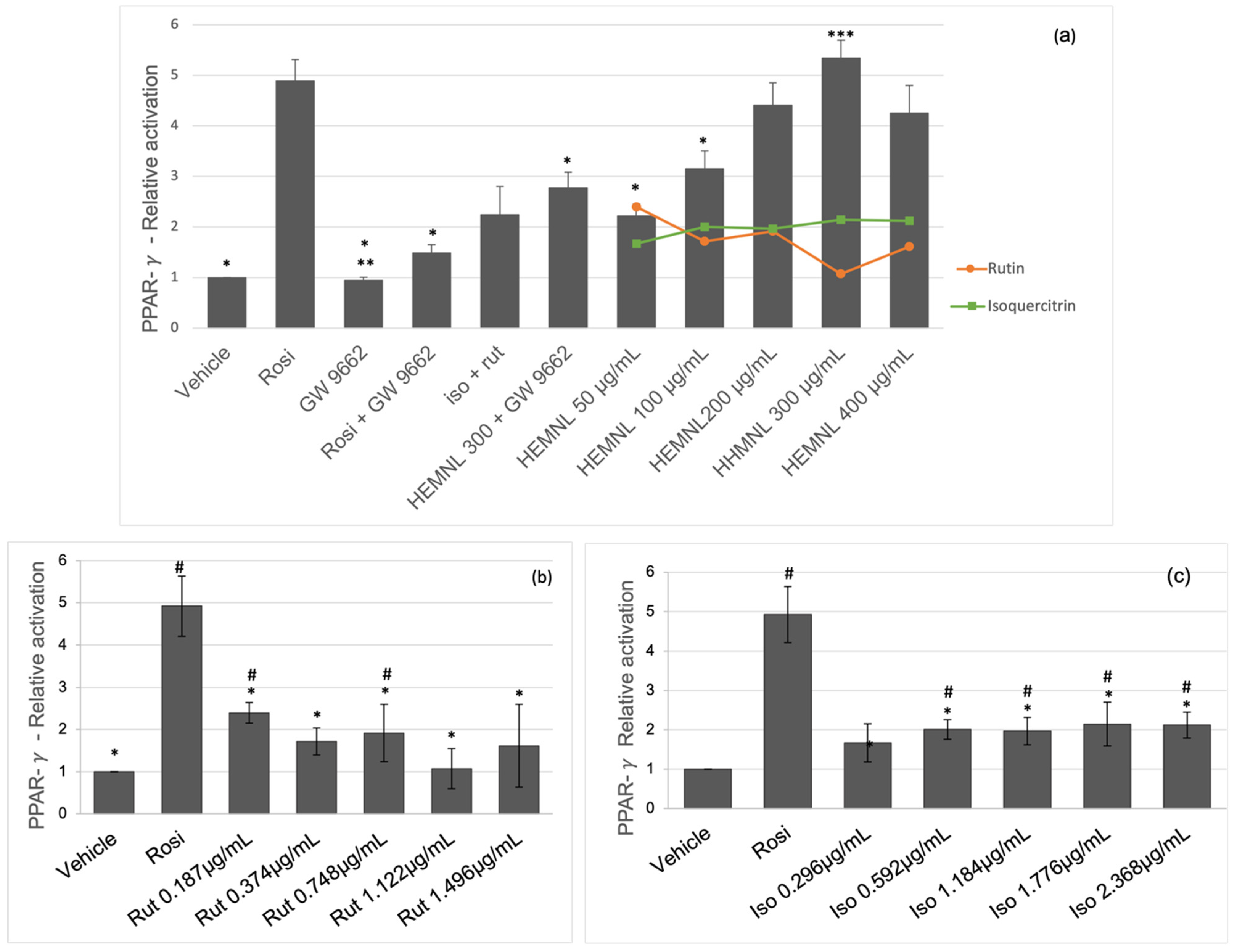
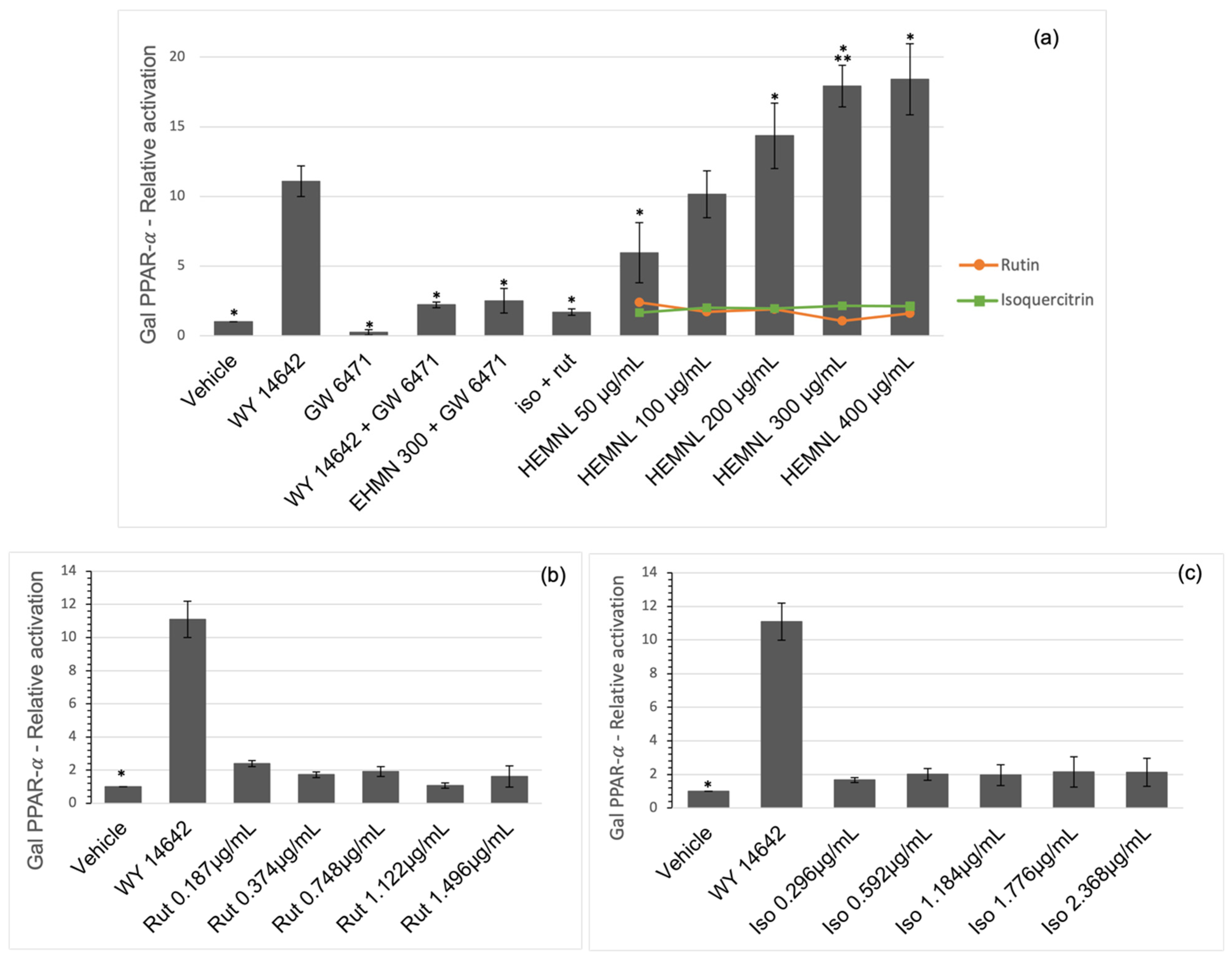
2.3. Activation of Nuclear Receptors PPAR-γ and PPAR-α
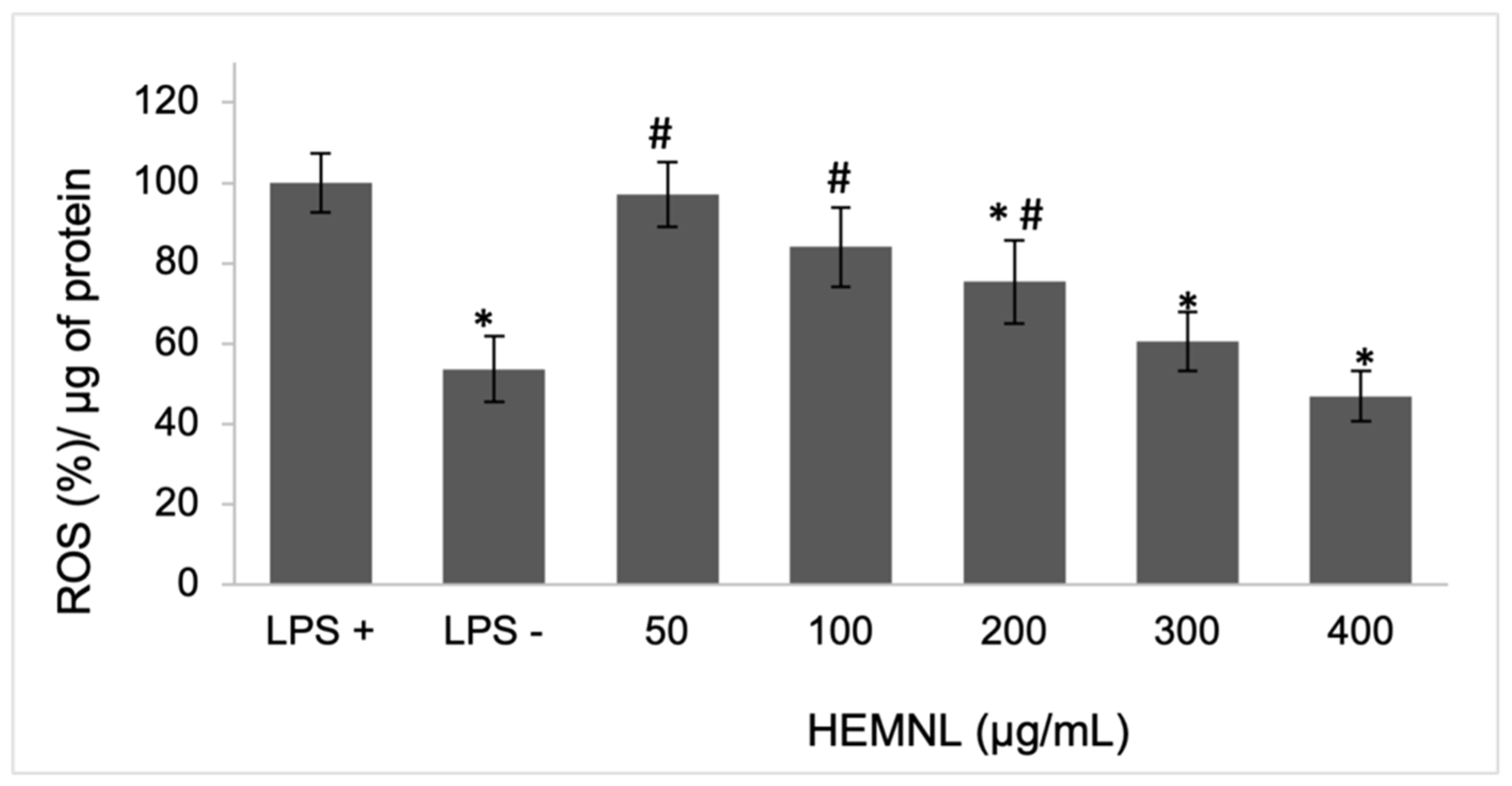
2.4. Intracellular ROS Reduction
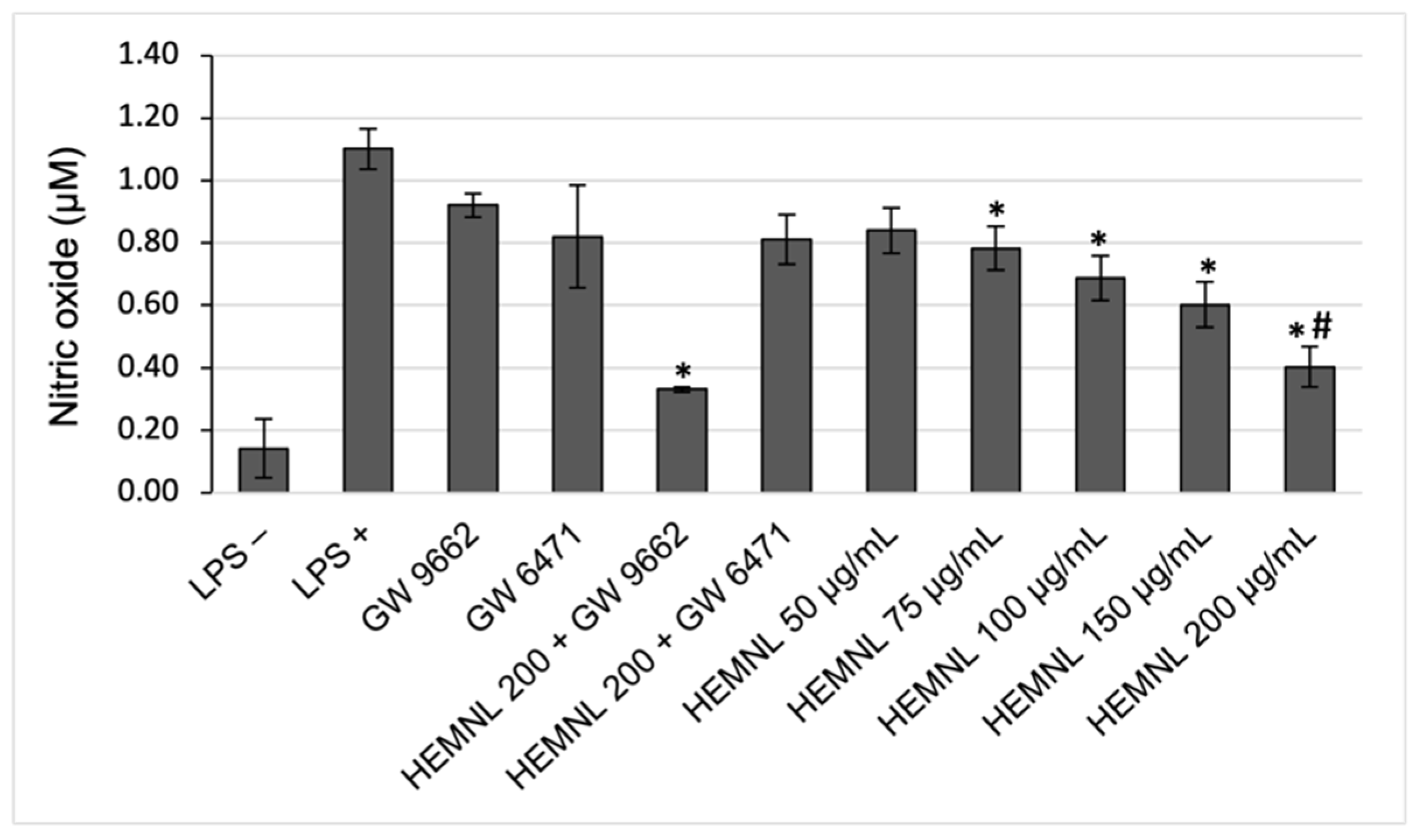
2.5. Reduction of NO Production
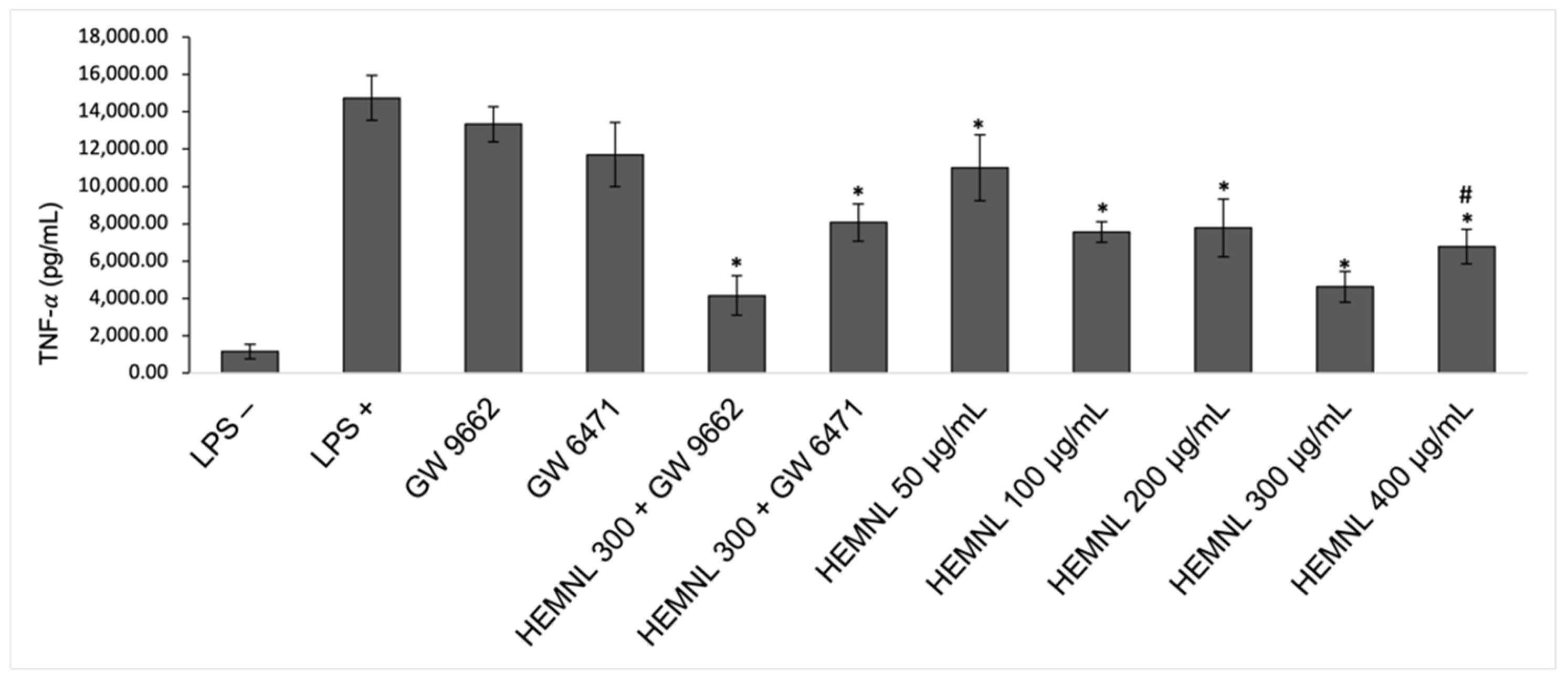
2.6. Reduction of Pro-Inflammatory Cytokine TNF-α
3. Discussion
4. Materials and Methods
4.1. Reagents and Standards
4.2. Plant Material and Extraction Procedure
4.3. HPLC-DAD Analysis
4.4. Cell Culture
4.5. Cell Viability Assay
4.6. PPAR Luciferase Reporter Assay
4.7. Measurement of Intracellular ROS
4.8. Nitrite Determination
4.9. TNF-α Determination
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Halim, M.; Halim, A. The Effects of Inflammation, Aging and Oxidative Stress on the Pathogenesis of Diabetes Mellitus (Type 2 Diabetes). Diabetes Metab. Syndr. Clin. Res. Rev. 2019, 13, 1165–1172. [Google Scholar] [CrossRef] [PubMed]
- Flynn, M.G.; Markofski, M.M.; Carrillo, A.E. Elevated Inflammatory Status and Increased Risk of Chronic Disease in Chronological Aging: Inflamm-Aging or Inflamm-Inactivity? Aging Dis. 2019, 10, 147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory Responses and Inflammation-Associated Diseases in Organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Libby, P.; Kobold, S. Inflammation: A Common Contributor to Cancer, Aging, and Cardiovascular Diseases—Expanding the Concept of Cardio-Oncology. Cardiovasc. Res. 2019, 115, 824–829. [Google Scholar] [CrossRef] [PubMed]
- Furman, D.; Campisi, J.; Verdin, E.; Carrera-Bastos, P.; Targ, S.; Franceschi, C.; Ferrucci, L.; Gilroy, D.W.; Fasano, A.; Miller, G.W.; et al. Chronic Inflammation in the Etiology of Disease across the Life Span. Nat. Med. 2019, 25, 1822–1832. [Google Scholar] [CrossRef] [PubMed]
- Vos, T.; Lim, S.S.; Abbafati, C.; Abbas, K.M.; Abbasi, M.; Abbasifard, M.; Abbasi-Kangevari, M.; Abbastabar, H.; Abd-Allah, F.; Abdelalim, A.; et al. Global Burden of 369 Diseases and Injuries in 204 Countries and Territories, 1990–2019: A Systematic Analysis for the Global Burden of Disease Study 2019. Lancet 2020, 396, 1204–1222. [Google Scholar] [CrossRef]
- van den Oever, I.A.M.; Baniaamam, M.; Simsek, S.; Raterman, H.G.; van Denderen, J.C.; van Eijk, I.C.; Peters, M.J.L.; van der Horst-Bruinsma, I.E.; Smulders, Y.M.; Nurmohamed, M.T. The Effect of Anti-TNF Treatment on Body Composition and Insulin Resistance in Patients with Rheumatoid Arthritis. Rheumatol. Int. 2021, 41, 319–328. [Google Scholar] [CrossRef]
- Burska, A.N.; Sakthiswary, R.; Sattar, N. Effects of Tumour Necrosis Factor Antagonists on Insulin Sensitivity/Resistance in Rheumatoid Arthritis: A Systematic Review and Meta-Analysis. PLoS ONE 2015, 10, e0128889. [Google Scholar] [CrossRef]
- Varga, T.; Czimmerer, Z.; Nagy, L. PPARs Are a Unique Set of Fatty Acid Regulated Transcription Factors Controlling Both Lipid Metabolism and Inflammation. Biochim. Biophys. Acta 2011, 1812, 1007–1022. [Google Scholar] [CrossRef]
- Martin, H. Role of PPAR-Gamma in Inflammation. Prospects for Therapeutic Intervention by Food Components. Mutat. Res. 2010, 690, 57–63. [Google Scholar] [CrossRef]
- Cheng, H.S.; Tan, W.R.; Low, Z.S.; Marvalim, C.; Lee, J.Y.H.; Tan, N.S. Tan Exploration and Development of PPAR Modulators in Health and Disease: An Update of Clinical Evidence. Int. J. Mol. Sci. 2019, 20, 5055. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization (WHO). Traditional Medicine Strategy 2014–2023; World Health Organization: Geneva, Switzerland, 2013; pp. 1–78. [Google Scholar]
- World Health Organization (WHO). WHO Global Report on Traditional and Complementary Medicine 2019; World Health Organization: Geneva, Switzerland, 2019. [Google Scholar]
- Ahmad Khan, M.S.; Ahmad, I. Herbal Medicine. In New Look to Phytomedicine; Elsevier: Amsterdam, The Netherlands, 2019; pp. 3–13. [Google Scholar]
- Miranda, M.A.; Vieira, G.D.-V.; Yamamoto, C.H.; de Sousa, O.V. Uso Etnomedicinal Do Chá de Morus Nigra L. No Tratamento Dos Sintomas Do Climatério de Mulheres de Muriaé, Minas Gerais, Brasil. Hu. Rev. 2010, 36, 61–68. [Google Scholar]
- Pio, I.D.S.L.; Lavor, A.L.; Damasceno, C.M.D.; Menezes, P.M.N.; Silva, F.S.; Maia, G.L.A. Traditional Knowledge and Uses of Medicinal Plants by the Inhabitants of the Islands of the São Francisco River, Brazil and Preliminary Analysis of Rhaphiodon Echinus (Lamiaceae). Braz. J. Biol. 2019, 79, 87–99. [Google Scholar] [CrossRef] [Green Version]
- Tng, D.Y.P.; Apgaua, D.M.G.; Lisboa, M.D.S.; El-Hani, C.N. Plants Uses in a Traditional Fisherman Community in Northeastern Brazil. bioRxiv 2019. [Google Scholar] [CrossRef]
- Hamza, N.; Berke, B.; Umar, A.; Cheze, C.; Gin, H. A Review of Algerian Medicinal Plants Used in the Treatment of Diabetes. J. Ethnopharmacol. 2019, 238, 111841. [Google Scholar] [CrossRef] [PubMed]
- Koncic, M.Z.; Bljaji, K. Traditional Herbal Products Used for the Management of Diabetes in Croatia: Linking Traditional Use With α -Glucosidase-Inhibitory Activity. In Bioactive Food as Dietary Interventions for Diabetes; Academic Press: Cambridge, MA, USA, 2019; pp. 647–664. ISBN 9780128138229. [Google Scholar]
- de Freitas, M.M.; Fontes, P.R.; Souza, P.M.; Fagg, C.W.; Guerra, E.N.S.; de Medeiros Nóbrega, Y.K.; Silveira, D.; Fonseca-Bazzo, Y.; Simeoni, L.A.; Homem-de-Mello, M.; et al. Extracts of Morus Nigra L Leaves Standardized in Chlorogenic Acid, Rutin and Isoquercitrin: Tyrosinase Inhibition and Cytotoxicity. PLoS ONE 2016, 11, e0163130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hago, S.; Mahrous, E.A.; Moawad, M.; Abdel-wahab, S.; Abdel-sattar, E. Evaluation of Antidiabetic Activity of Morus Nigra L. and Bauhinia Variegata L. Leaves as Egyptian Remedies Used for the Treatment of Diabetes. Nat. Prod. Res. 2019, 35, 829–835. [Google Scholar] [CrossRef]
- Ribeiro, A.E.A.S.; Soares, J.M.D.; Silva, H.A.L.; de Wanderley, C.W.S.; Moura, C.A.; Oliveira-Júnior, R.G.; Oliveira, A.P.; Rolim, L.A.; Costa, E.V.; da Almeida, J.R.G.S.; et al. Inhibitory Effects of Morus Nigra L. (Moraceae) against Local Paw Edema and Mechanical Hypernociception Induced by Bothrops Jararacussu Snake Venom in Mice. Biomed. Pharmacother. 2019, 111, 1046–1056. [Google Scholar] [CrossRef]
- Lim, H.; Heo, M.Y.; Kim, H.P. Flavonoids: Broad Spectrum Agents on Chronic Inflammation. Biomol. Ther. 2019, 27, 241–253. [Google Scholar] [CrossRef]
- Serino, A.; Salazar, G. Protective Role of Polyphenols against Vascular Inflammation, Aging and Cardiovascular Disease. Nutrients 2019, 11, 53. [Google Scholar] [CrossRef] [Green Version]
- 10993–5; 2009 Biological Evaluation of Medical Devices—Part 5: Tests for in Vitro Cytotoxicity. International Organization for Standardization: Geneva, Switzerland, 2009.
- Cavuldak, Ö.A.; Ertan, R.; Vural, N.; Akay, M.A. Optimization of Ultrasound-Assisted Water Extraction Conditions for the Extraction of Phenolic Compounds from Black Mulberry Leaves (Morus Nigra L.). J. Food Process. Eng. 2019, 42, e13132. [Google Scholar] [CrossRef]
- Schafranski, K.; Postigo, M.P.; Vitali, L.; Micke, G.A.; Richter, W.E. Avaliação de Compostos Bioativos e Atividade Antioxidante de Extratos de Folhas de Amoreira Preta (Morus Nigra L.) Utilizando Planejamento Experimental. Quim. Nova 2019, 42, 736–744. [Google Scholar] [CrossRef]
- Sánchez-Salcedo, E.M.; Mena, P.; García-Viguera, C.; Hernández, F.; Martinez, J.J. (Poly)Phenolic Compounds and Antioxidant Activity of White (Morus Alba) and Black (Morus Nigra) Mulberry Leaves: Their Potential for New Products Rich in Phytochemicals. J. Funct. Foods 2015, 18, 1039–1046. [Google Scholar] [CrossRef]
- Figueredo, K.C.; Guex, C.G.; Reginato, F.Z.; Haas, A.R.; Cassanego, G.B.; Lima, C.; Boligon, A.A.; Helena, G.; Lopes, H.; Bauermann, L.D.F.; et al. Safety Assessment of Morus Nigra L. Leaves: Acute and Subacute Oral Toxicity Studies in Wistar Rats. J. Ethnopharmacol. 2018, 224, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Zeni, A.N.A.L.B.; Moreira, T.D.; Dalmagro, A.N.A.P.; Camargo, A.; Bini, L.A.; Simionatto, E.; Scharf, D. Evaluation of Phenolic Compounds and Lipid-Lowering Effect of Morus Nigra Leaves Extract. An. Acad. Bras. Cienc. 2017, 89, 2805–2815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turgut, N.H.; Mert, D.G.; Kara, H.; Egilmez, H.R.; Arslanbas, E.; Tepe, B.; Gungor, H.; Yilmaz, N.; Baris, N. Effect of Black Mulberry (Morus Nigra) Extract Treatment on Cognitive Impairment and Oxidative Stress Status of d-Galactose-Induced Aging Mice. Pharm. Biol. 2016, 54, 1052–1064. [Google Scholar] [CrossRef]
- Simões, C.M.O.; Schenkel, E.P.; Mello, J.C.P.; Mentz, L.A.; Petrovick, P.R. Farmacognosia—Do Produto Natural Ao Medicamento, 1st ed.; Artmed: Porto Alegre, Brazil, 2017. [Google Scholar]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An Overview. J. Nutr. Sci. 2016, 5, 41. [Google Scholar] [CrossRef] [Green Version]
- Tag, H.M. Hepatoprotective Effect of Mulberry (Morus Nigra) Leaves Extract against Methotrexate Induced Hepatotoxicity in Male Albino Rat. BMC Complement. Altern. Med. 2015, 15, 252. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Yu, W.; Chen, G.; Meng, S.; Xiang, Z.; He, N. Antinociceptive and Antibacterial Properties of Anthocyanins and Flavonols from Fruits of Black and Non-Black Mulberries. Molecules 2018, 23, 4. [Google Scholar] [CrossRef] [Green Version]
- Evans, R.M.; Barish, G.D.; Wang, Y.X. PPARs and the Complex Journey to Obesity. Nat. Med. 2004, 10, 355–361. [Google Scholar] [CrossRef]
- Nelson, D.L.; Cox, M.M. Principios de Bioquímica de Lehninger, 6th ed.; Artmed: Porto Alegre, Brazil, 2014; ISBN 9781429234146. [Google Scholar]
- Tyagi, S.; Gupta, P.; Saini, A.; Kaushal, C.; Sharma, S. The Peroxisome Proliferator-Activated Receptor: A Family of Nuclear Receptors Role in Various Diseases. J. Adv. Pharm. Technol. Res. 2011, 2, 236. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.K.; Zhuang, Y.; Wahli, W. Synthetic and Natural Peroxisome Proliferator-Activated Receptor (PPAR) Agonists as Candidates for the Therapy of the Metabolic Syndrome. Expert. Opin. Ther. Targets 2017, 21, 333–348. [Google Scholar] [CrossRef]
- Xu, L.J.; Yu, M.H.; Huang, C.Y.; Niu, L.X.; Wang, Y.F.; Wu, C.Z.; Yang, P.M.; Hu, X. Isoprenylated Flavonoids from Morus Nigra and Their PPAR γ Agonistic Activities. Fitoterapia 2018, 127, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Berger, J.; Moller, D.E. The Mechanisms of Action of PPARs. Annu. Rev. Med. 2002, 53, 409–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, H.; Ma, Q.; Cui, H.; Liu, G.; Zhao, X.; Li, W.; Piao, G. How Can Synergism of Traditional Medicines Benefit from Network Pharmacology? Molecules 2017, 22, 1135. [Google Scholar] [CrossRef] [Green Version]
- Elekofehinti, O. In Silico Studies on Plant Derived Rutin as Potent Agonist of Peroxisome Proliferator-Activated Receptor Gamma (PPARγ). Br. J. Med. Med. Res. 2016, 14, 1–8. [Google Scholar] [CrossRef]
- Mandal, S.K.; Kumar, B.K.; Sharma, P.K.; Murugesan, S.; Deepa, P.R. In Silico and in Vitro Analysis of PPAR—α/γ Dual Agonists: Comparative Evaluation of Potential Phytochemicals with Anti-Obesity Drug Orlistat. Comput. Biol. Med. 2022, 147, 105796. [Google Scholar] [CrossRef]
- Seeley, J.J.; Ghosh, S. Molecular Mechanisms of Innate Memory and Tolerance to LPS. J. Leukoc. Biol. 2017, 101, 107–119. [Google Scholar] [CrossRef]
- Hoesel, B.; Schmid, J.A. The Complexity of NF-ΚB Signaling in Inflammation and Cancer. Mol. Cancer 2013, 12, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Serasanambati, M.; Chilakapati, S.R. Function of Nuclear Factor Kappa B (NF-KB) in Human Diseases-A Review. S. Indian J. Biol. Sci. 2016, 2, 368. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-ΚB Signaling in Inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [PubMed]
- Rius-Pérez, S.; Pérez, S.; Martí-Andrés, P.; Monsalve, M.; Sastre, J. Nuclear Factor Kappa B Signaling Complexes in Acute Inflammation. Antioxid. Redox Signal. 2020, 33, 145–165. [Google Scholar] [CrossRef] [PubMed]
- Page, M.J.; Kell, D.B.; Pretorius, E. The Role of Lipopolysaccharide-Induced Cell Signalling in Chronic Inflammation. Chronic Stress 2022, 6, 247054702210763. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Abbas, A.K.; Aster, J.C. Inflamação e Reparo. In Robbins Patologia Básica; Elsevier: Amsterdam, The Netherlands, 2013. [Google Scholar]
- Rendra, E.; Riabov, V.; Mossel, D.M.; Sevastyanova, T.; Harmsen, M.C.; Kzhyshkowska, J. Reactive Oxygen Species (ROS) in Macrophage Activation and Function in Diabetes. Immunobiology 2019, 224, 242–253. [Google Scholar] [CrossRef]
- Eruygur, N.; Dural, E. Determination of 1-Deoxynojirimycin by a Developed and Validated HPLC-FLD Method and Assessment of In-Vitro Antioxidant, α-Amylase and α-Glucosidase Inhibitory Activity in Mulberry Varieties from Turkey. Phytomedicine 2019, 53, 234–242. [Google Scholar] [CrossRef]
- Dalmagro, A.P.; Camargo, A.; Lúcia, A.; Zeni, B. Morus Nigra and Its Major Phenolic, Syringic Acid, Have Antidepressant-like and Neuroprotective Effects in Mice. Metab. Brain Dis. 2017, 32, 1963–1973. [Google Scholar] [CrossRef]
- Montenote, M.C.; Wajsman, V.Z.; Konno, Y.T.; Ferreira, P.C.; Silva, R.M.G.; Therezo, A.L.S.; Silva, L.P.; Martins, L.P.A. Antioxidant Effect of Morus Nigra on Chagas Disease Progression. Rev. Inst. Med. Trop. Sao Paulo 2017, 59, e73. [Google Scholar] [CrossRef] [Green Version]
- Biswas, S.K. Does the Interdependence between Oxidative Stress and Inflammation Explain the Antioxidant Paradox? Oxid. Med. Cell. Longev. 2016, 2016, 17–19. [Google Scholar] [CrossRef] [Green Version]
- Marchio, P.; Guerra-Ojeda, S.; Vila, J.M.; Aldasoro, M.; Victor, V.M.; Mauricio, M.D. Targeting Early Atherosclerosis: A Focus on Oxidative Stress and Inflammation. Oxid. Med. Cell. Longev. 2019, 2019, 8563845. [Google Scholar] [CrossRef] [Green Version]
- Gill, R.; Tsung, A.; Billiar, T. Linking Oxidative Stress to Inflammation: Toll-like Receptors. Free Radic. Biol. Med. 2010, 48, 1121–1132. [Google Scholar] [CrossRef] [Green Version]
- Cinelli, M.A.; Do, H.T.; Miley, G.P.; Silverman, R.B. Inducible Nitric Oxide Synthase: Regulation, Structure, and Inhibition. Med. Res. Rev. 2020, 40, 158–189. [Google Scholar] [CrossRef] [PubMed]
- Spiller, F.; Oliveira Formiga, R.; Fernandes da Silva Coimbra, J.; Alves-Filho, J.C.; Cunha, T.M.; Cunha, F.Q. Targeting Nitric Oxide as a Key Modulator of Sepsis, Arthritis and Pain. Nitric Oxide 2019, 89, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Kashfi, K.; Vannini, F. Nitric Oxide and Cancer: To Inhibit or To Induce INOS: That Is the Question? Elsevier: Amsterdam, The Netherlands, 2019; ISBN 9780128165454. [Google Scholar]
- Oppenheim, J.J. Cytokines, Their Receptors and Signals; Elsevier: Amsterdam, The Netherlands, 2020; ISBN 9780128121023. [Google Scholar]
- Lewis, D.E.; Blutt, S.E. Organization of the Immune System, 5th ed.; Elsevier: Amsterdam, The Netherlands, 2019; Volume 1, ISBN 9780702068966. [Google Scholar]
- Zhang, C.; Deng, J.; Liu, D.; Tuo, X.; Yu, Y.; Yang, H.; Wang, N. Nuciferine Inhibits Proinflammatory Cytokines via the PPARs in LPS-Induced RAW264.7 Cells. Molecules 2018, 23, 2723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lúcio, K.D.P.; Carolina, A.; Rabelo, S.; Araújo, C.M.; Brandão, G.C.; Henrique, G.; de Souza, B.; Gomes, R.; Maria, D.; de Souza, S.; et al. Anti-Inflammatory and Antioxidant Properties of Black Mulberry (Morus Nigra L.) in a Model of LPS-Induced Sepsis. Oxid. Med. Cell. Longev. 2018, 2018, 5048031. [Google Scholar] [CrossRef] [Green Version]
- Hou, L.S.; Cui, Z.Y.; Sun, P.; Piao, H.Q.; Han, X.; Song, J.; Wang, G.; Zheng, S.; Dong, X.X.; Gao, L.; et al. Rutin Mitigates Hepatic Fibrogenesis and Inflammation through Targeting TLR4 and P2X7 Receptor Signaling Pathway in Vitro and in Vivo. J. Funct. Foods 2020, 64, 103700. [Google Scholar] [CrossRef]
- Nikfarjam, B.A.; Adineh, M.; Hajiali, F.; Nassiri-Asl, M. Treatment with Rutin—A Therapeutic Strategy for Neutrophil-Mediated Inflammatory and Autoimmune Diseases: Anti-Inflammatory Effects of Rutin on Neutrophils. J. Pharmacopunct. 2017, 20, 52–56. [Google Scholar] [CrossRef]
- Al-Harbi, N.O.; Imam, F.; Al-Harbi, M.M.; Al-Shabanah, O.A.; Alotaibi, M.R.; As Sobeai, H.M.; Afzal, M.; Kazmi, I.; al Rikabi, A.C. Rutin Inhibits Carfilzomib-Induced Oxidative Stress and Inflammation via the NOS-Mediated NF-ΚB Signaling Pathway. Inflammopharmacology 2019, 27, 817–827. [Google Scholar] [CrossRef]
- Nkpaa, K.W.; Onyeso, G.I. Rutin Attenuates Neurobehavioral Deficits, Oxidative Stress, Neuro-Inflammation and Apoptosis in Fluoride Treated Rats. Neurosci. Lett. 2018, 682, 92–99. [Google Scholar] [CrossRef]
- Taşlı, N.G.; Çimen, F.K.; Karakurt, Y.; Uçak, T.; Mammadov, R.; Süleyman, B.; Kurt, N.; Süleyman, H. Protective Effects of Rutin against Methanol Induced Acute Toxic Optic Neuropathy: An Experimental Study. Int. J. Ophthalmol. 2018, 11, 780–785. [Google Scholar] [CrossRef]
- Xie, P.S.; Leung, A.Y. Understanding the Traditional Aspect of Chinese Medicine in Order to Achieve Meaningful Quality Control of Chinese Materia Medica. J. Chromatogr. A 2009, 1216, 1933–1940. [Google Scholar] [CrossRef]
- Huang, S.H.; Xu, M.; Wu, H.M.; Wan, C.X.; Wang, H.B.; Wu, Q.Q.; Liao, H.H.; Deng, W.; Tang, Q.Z. Isoquercitrin Attenuated Cardiac Dysfunction Via AMPKα-Dependent Pathways in LPS-Treated Mice. Mol. Nutr. Food Res. 2018, 62, 1800955. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T. Rapid Colorimetric Assay for Cellular Growth and Survival: Application to Proliferation and Cytotoxicity Assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time (min) | Phosphoric Acid 1% (%) | Acetonitrile (%) |
|---|---|---|
| 0 | 90 | 10 |
| 40 | 70 | 30 |
| 45 | 90 | 10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carneiro, A.d.A.; Sinoti, S.B.P.; de Freitas, M.M.; Simeoni, L.A.; Fagg, C.W.; Magalhães, P.d.O.; Silveira, D.; Fonseca-Bazzo, Y.M. Hydroethanolic Extract of Morus nigra L. Leaves: A Dual PPAR-α/γ Agonist with Anti-Inflammatory Properties in Lipopolysaccharide-Stimulated RAW 264.7. Plants 2022, 11, 3147. https://doi.org/10.3390/plants11223147
Carneiro AdA, Sinoti SBP, de Freitas MM, Simeoni LA, Fagg CW, Magalhães PdO, Silveira D, Fonseca-Bazzo YM. Hydroethanolic Extract of Morus nigra L. Leaves: A Dual PPAR-α/γ Agonist with Anti-Inflammatory Properties in Lipopolysaccharide-Stimulated RAW 264.7. Plants. 2022; 11(22):3147. https://doi.org/10.3390/plants11223147
Chicago/Turabian StyleCarneiro, Amanda de Assis, Simone Batista Pires Sinoti, Marcela Medeiros de Freitas, Luiz Alberto Simeoni, Christopher William Fagg, Pérola de Oliveira Magalhães, Dâmaris Silveira, and Yris Maria Fonseca-Bazzo. 2022. "Hydroethanolic Extract of Morus nigra L. Leaves: A Dual PPAR-α/γ Agonist with Anti-Inflammatory Properties in Lipopolysaccharide-Stimulated RAW 264.7" Plants 11, no. 22: 3147. https://doi.org/10.3390/plants11223147