
Fruit Morphology and Ripening-Related QTLs in a Newly Developed Introgression Line Collection of the Elite Varieties ‘Védrantais’ and ‘Piel de Sapo’

 , , , and
, , , and
Abstract
:1. Introduction
2. Results and Discussion
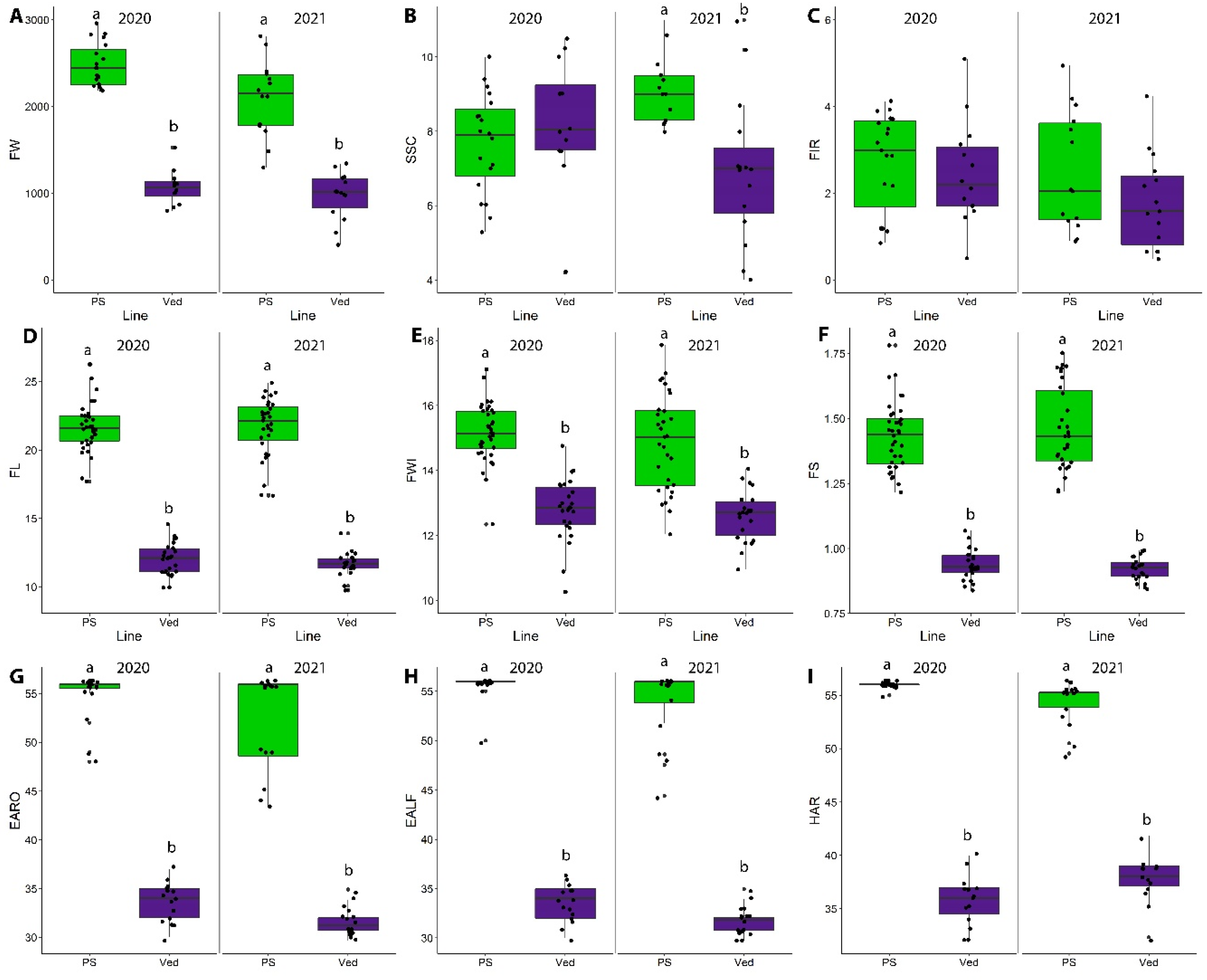
2.1. Phenotyping of the Parental Lines
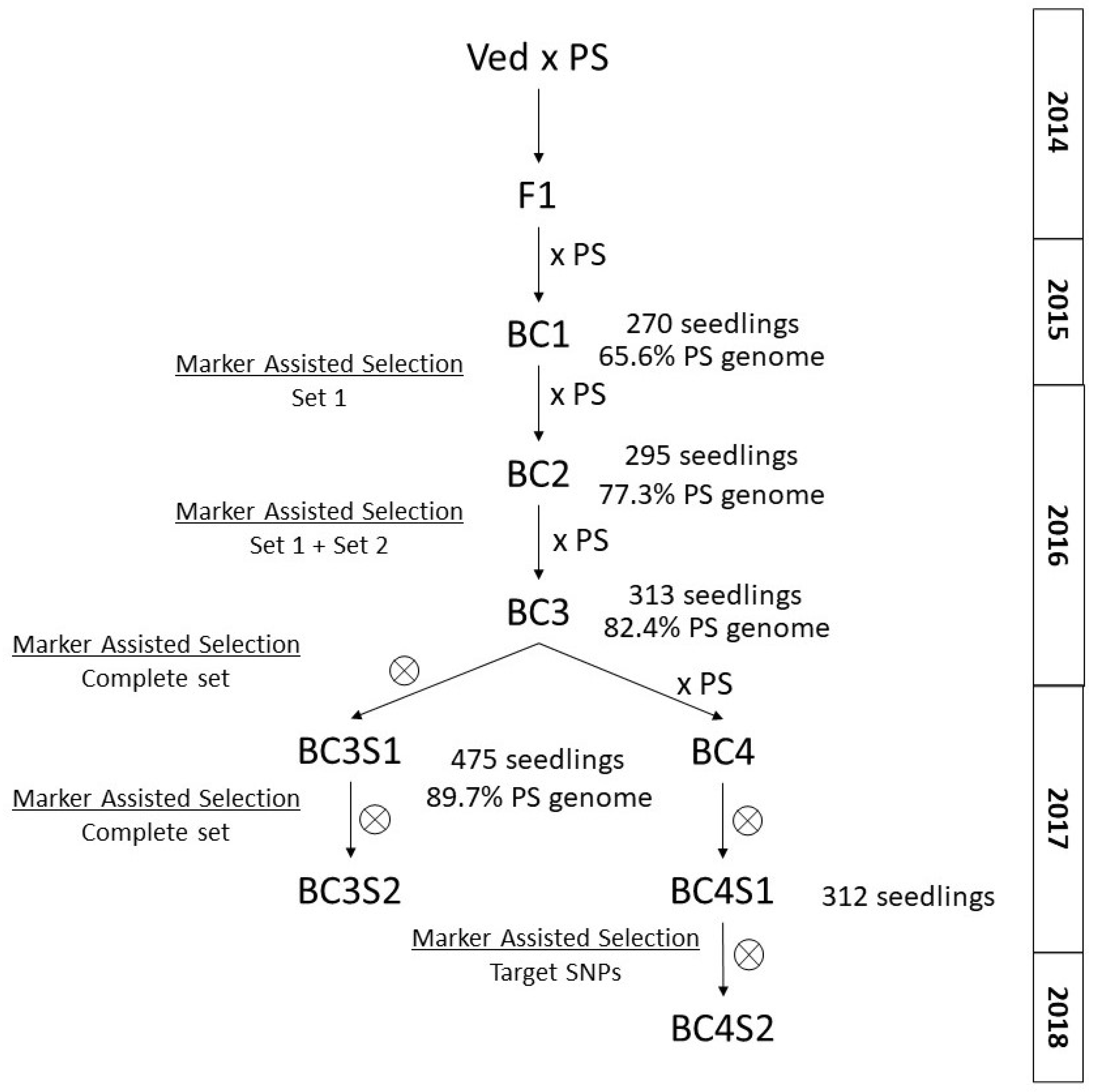
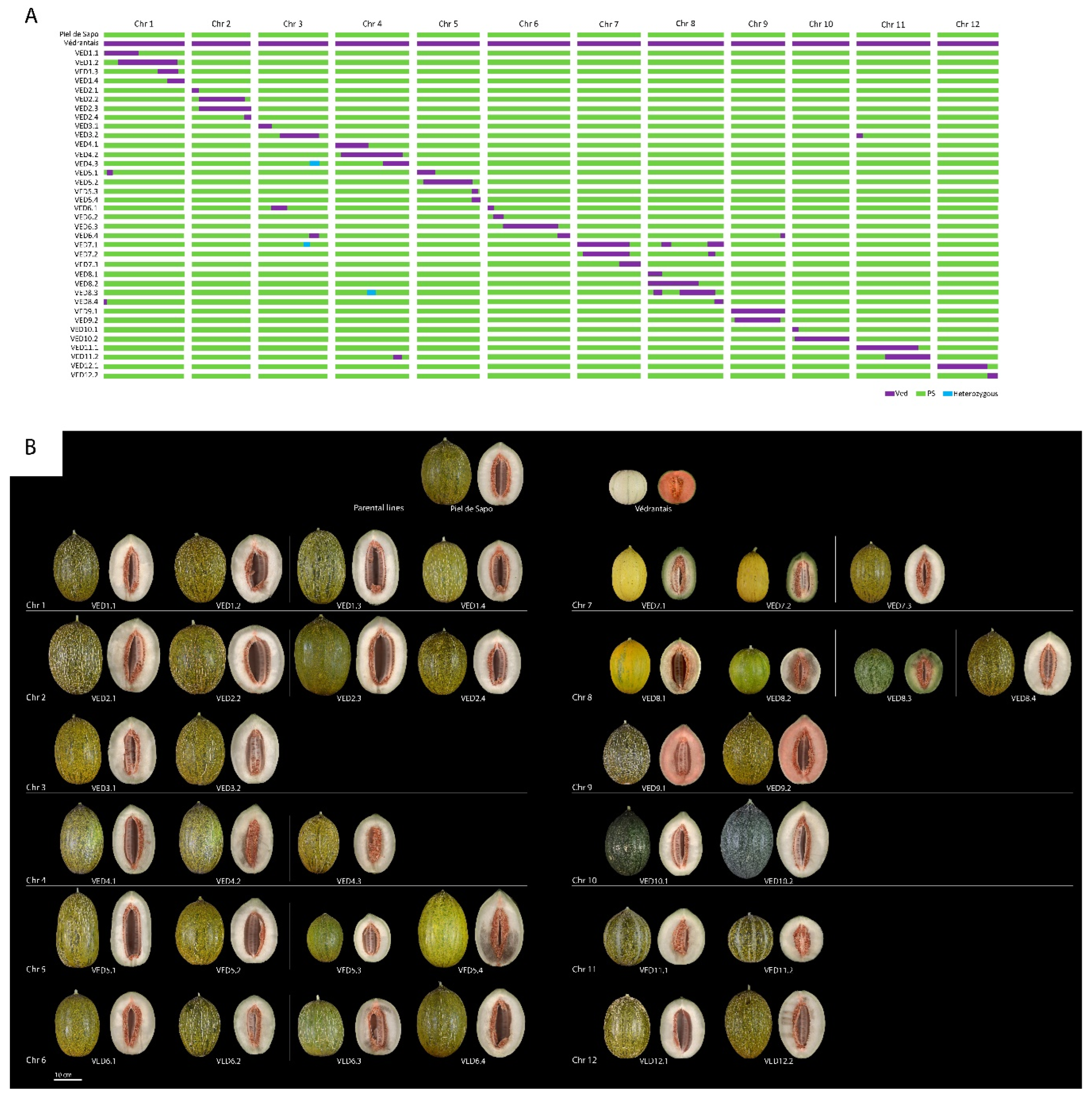
2.2. Development of the IL Collection
2.3. Phenotyping of the IL Population and QTL Mapping
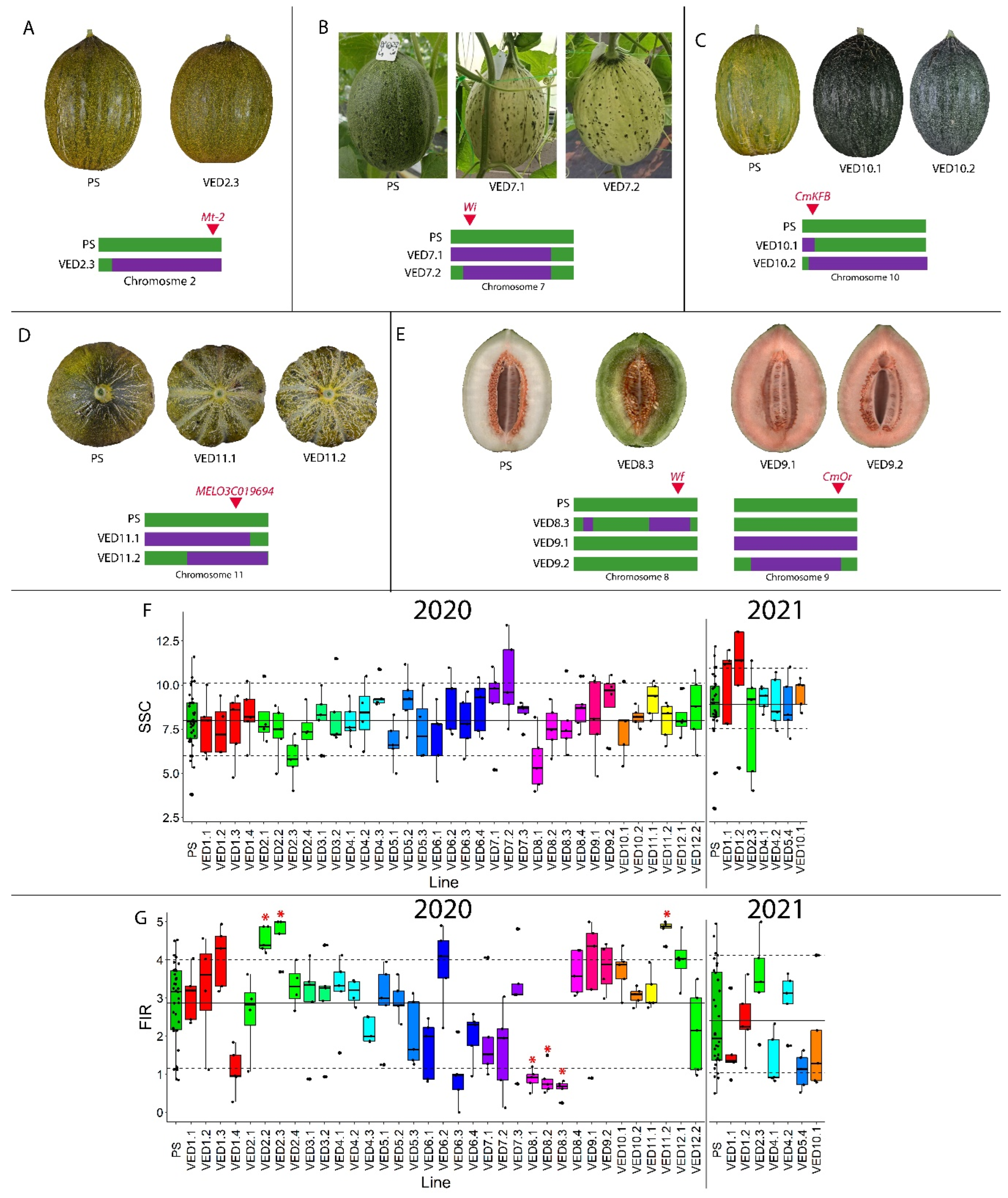
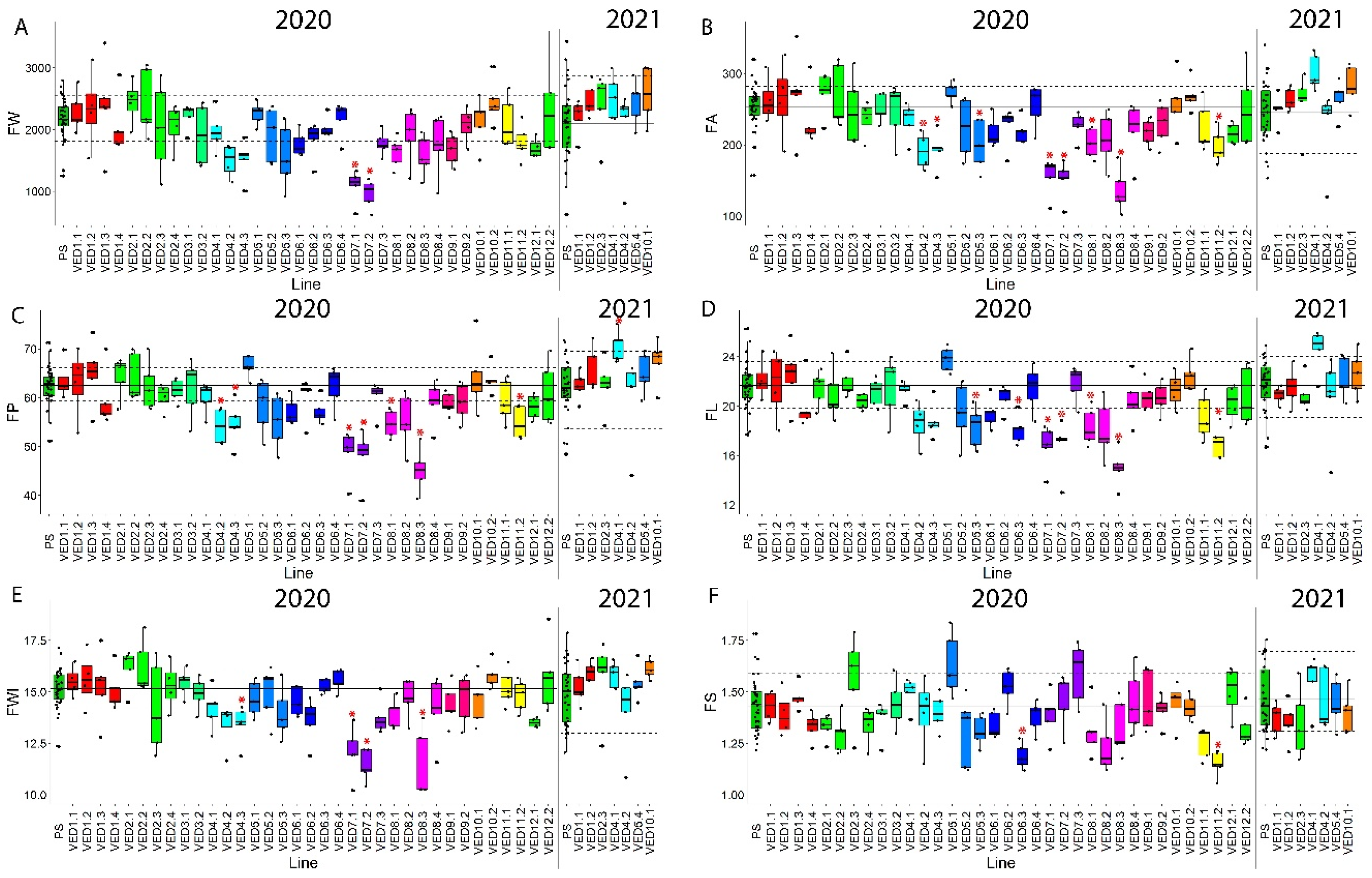
2.3.1. Fruit Quality Traits
2.3.2. Fruit Morphology Traits
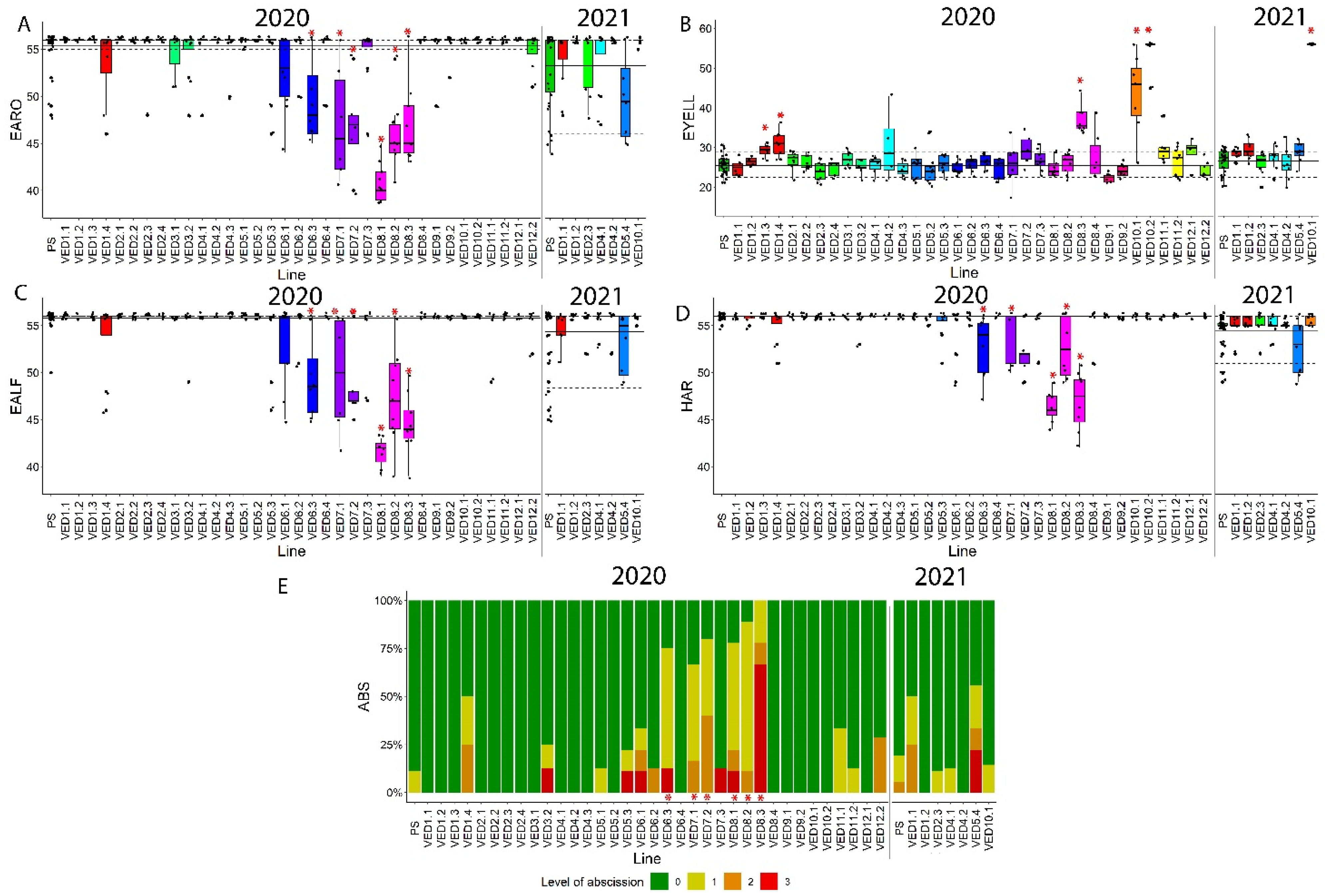
2.3.3. Ripening Traits
3. Materials and Methods
3.1. Plant Material and Breeding Scheme
3.2. In Vitro Plant Culture
3.3. DNA Extraction and Genotyping
3.4. Experimental Design and Phenotyping
3.5. QTL Mapping and Data Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bhoite, R.; Onyemaobi, I.; Si, P.; Siddique, K.H.M.; Yan, G. Identification and Validation of QTL and Their Associated Genes for Pre-Emergent Metribuzin Tolerance in Hexaploid Wheat (Triticum aestivum L.). BMC Genet. 2018, 19, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Z.; Cheng, Y.; Xian, P.; Lin, R.; Xia, Q.; He, X.; Liang, Q.; Lian, T.; Ma, Q.; Nian, H. Fine-Mapping QTLs and the Validation of Candidate Genes for Aluminum Tolerance Using a High-Density Genetic Map. Plant Soil 2019, 444, 119–137. [Google Scholar] [CrossRef]
- Zhaoming, Q.; Xiaoying, Z.; Huidong, Q.; Dawei, X.; Xue, H.; Hongwei, J.; Zhengong, Y.; Zhanguo, Z.; Jinzhu, Z.; Rongsheng, Z.; et al. Identification and Validation of Major QTLs and Epistatic Interactions for Seed Oil Content in Soybeans under Multiple Environments Based on a High-Density Map. Euphytica 2017, 213, 162. [Google Scholar] [CrossRef]
- Fukino, N.; Kunihisa, M.; Matsumoto, S. Characterization of Recombinant Inbred Lines Derived from Crosses in Melon (Cucumis melo L.), ‘PMAR No. 5′ × ‘Harukei No. 3’. Breed. Sci. 2004, 54, 141–145. [Google Scholar] [CrossRef] [Green Version]
- Kuang, L.; Ahmad, N.; Su, B.; Huang, L.; Li, K.; Wang, H.; Wang, X.; Dun, X. Discovery of Genomic Regions and Candidate Genes Controlling Root Development Using a Recombinant Inbred Line Population in Rapeseed (Brassica napus L.). Int. J. Mol. Sci. 2022, 23, 4781. [Google Scholar] [CrossRef]
- Clark, C.B.; Wang, W.; Wang, Y.; Fear, G.J.; Wen, Z.; Wang, D.; Ren, B.; Ma, J. Identification and Molecular Mapping of a Major Quantitative Trait Locus Underlying Branch Angle in Soybean. Theor. Appl. Genet. 2022, 135, 777–784. [Google Scholar] [CrossRef] [PubMed]
- Endelman, J.B.; Jansky, S.H. Genetic Mapping with an Inbred Line-Derived F2 Population in Potato. Theor. Appl. Genet. 2016, 129, 935–943. [Google Scholar] [CrossRef]
- Zhang, H.; Zang, J.; Huo, Y.; Zhang, Z.; Chen, H.; Chen, X.; Liu, J. Identification of the Potential Genes Regulating Seed Germination Speed in Maize. Plants 2022, 11, 556. [Google Scholar] [CrossRef]
- Wang, R.X.; Hai, L.; Zhang, X.Y.; You, G.X.; Yan, C.S.; Xiao, S.H. QTL Mapping for Grain Filling Rate and Yield-Related Traits in RILs of the Chinese Winter Wheat Population Heshangmai × Yu8679. Theor. Appl. Genet. 2009, 118, 313–325. [Google Scholar] [CrossRef]
- Wang, L.; Cheng, Y.; Ma, Q.; Mu, Y.; Huang, Z.; Xia, Q.; Zhang, G.; Nian, H. QTL Fine-Mapping of Soybean (Glycine max L.) Leaf Type Associated Traits in Two RILs Populations. BMC Genom. 2019, 20, 260. [Google Scholar] [CrossRef]
- Li, J.X.; Yu, S.B.; Xu, C.G.; Tan, Y.F.; Gao, Y.J.; Li, X.H.; Zhang, Q. Analyzing Quantitative Trait Loci for Yield Using a Vegetatively Replicated F2 Population from a Cross between the Parents of an Elite Rice Hybrid. Theor. Appl. Genet. 2000, 101, 248–254. [Google Scholar] [CrossRef]
- Harel-Beja, R.; Sherman, A.; Rubinstein, M.; Eshed, R.; Bar-Ya’akov, I.; Trainin, T.; Ophir, R.; Holland, D. A Novel Genetic Map of Pomegranate Based on Transcript Markers Enriched with QTLs for Fruit Quality Traits. Tree Genet. Genomes 2015, 11, 109. [Google Scholar] [CrossRef]
- Eshed, Y.; Zamir, D. An Introgression Line Population of Lycopersicon Pennellii in the Cultivated Tomato Enables the Identification and Fine Mapping of Yield-Associated QTL. Genetics 1995, 141, 1147–1162. [Google Scholar] [CrossRef] [PubMed]
- Perpiñá, G.; Esteras, C.; Gibon, Y.; Monforte, A.J.; Picó, B. A New Genomic Library of Melon Introgression Lines in a Cantaloupe Genetic Background for Dissecting Desirable Agronomical Traits. BMC Plant Biol. 2016, 16, 154. [Google Scholar] [CrossRef] [PubMed]
- Balakrishnan, D.; Surapaneni, M.; Mesapogu, S.; Neelamraju, S. Development and Use of Chromosome Segment Substitution Lines as a Genetic Resource for Crop Improvement. Theor. Appl. Genet. 2019, 132, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Zhao, Q.; Du, P.; Xu, C.; Wang, B.; Feng, Q.; Liu, Q.; Tang, S.; Gu, M.; Han, B.; et al. Developing High Throughput Genotyped Chromosome Segment Substitution Lines Based on Population Whole-Genome Re-Sequencing in Rice (Oryza sativa L.). BMC Genom. 2010, 11, 656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szalma, S.J.; Hostert, B.M.; LeDeaux, J.R.; Stuber, C.W.; Holland, J.B. QTL Mapping with Near-Isogenic Lines in Maize. Theor. Appl. Genet. 2007, 114, 1211–1228. [Google Scholar] [CrossRef]
- Fujita, D.; Tagle, A.G.; Koide, Y.; Simon, E.V.; Fukuta, Y.; Ishimaru, T.; Kobayashi, N. Characterization of QTLs for Grain Weight from New Plant Type Rice Cultivars through the Development of Near-Isogenic Lines with an IR 64 Background. Euphytica 2022, 218, 50. [Google Scholar] [CrossRef]
- Essafi, A.; Díaz-Pendón, J.A.; Moriones, E.; Monforte, A.J.; Garcia-Mas, J.; Martín-Hernández, A.M. Dissection of the Oligogenic Resistance to Cucumber Mosaic Virus in the Melon Accession PI 161375. Theor. Appl. Genet. 2009, 118, 275–284. [Google Scholar] [CrossRef]
- Giner, A.; Pascual, L.; Bourgeois, M.; Gyetvai, G.; Rios, P.; Picó, B.; Troadec, C.; Bendahmane, A.; Garcia-Mas, J.; Martín-Hernández, A.M. A Mutation in the Melon Vacuolar Protein Sorting 41 prevents Systemic Infection of Cucumber Mosaic Virus. Sci. Rep. 2017, 7, 10471. [Google Scholar] [CrossRef]
- Babar, A.D.; Zaka, A.; Naveed, S.A.; Ahmad, N.; Aslam, K.; Asif, M.; Maqsood, U.; Vera Cruz, C.M.; Arif, M. Development of Basmati Lines by the Introgression of Three Bacterial Blight Resistant Genes through Marker-Assisted Breeding. Euphytica 2022, 218, 59. [Google Scholar] [CrossRef]
- Lavaud, C.; Baviere, M.; Le Roy, G.; Hervé, M.R.; Moussart, A.; Delourme, R.; Pilet-Nayel, M.-L. Single and Multiple Resistance QTL Delay Symptom Appearance and Slow down Root Colonization by Aphanomyces Euteiches in Pea near Isogenic Lines. BMC Plant Biol. 2016, 16, 166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, L.; Ruggieri, V.; Pérez, S.; Alexiou, K.G.; Fernández, M.; Jahrmann, T.; Pujol, M.; Garcia-Mas, J. QTL Mapping of Melon Fruit Quality Traits Using a High-Density GBS-Based Genetic Map. BMC Plant Biol. 2018, 18, 324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oren, E.; Tzuri, G.; Dafna, A.; Meir, A.; Kumar, R.; Katzir, N.; Elkind, Y.; Freilich, S.; Schaffer, A.A.; Tadmor, Y.; et al. High-Density NGS-Based Map Construction and Genetic Dissection of Fruit Shape and Rind Netting in Cucumis melo. Theor. Appl. Genet. 2020, 133, 1927–1945. [Google Scholar] [CrossRef] [PubMed]
- Dai, D.; Zeng, S.; Wang, L.; Li, J.; Ji, P.; Liu, H.; Sheng, Y. Identification of Fruit Firmness QTL Ff2.1 by SLAF-BSA and QTL Mapping in Melon. Euphytica 2022, 218, 52. [Google Scholar] [CrossRef]
- Argyris, J.M.; Díaz, A.; Ruggieri, V.; Fernández, M.; Jahrmann, T.; Gibon, Y.; Picó, B.; Martín-Hernández, A.M.; Monforte, A.J.; Garcia-Mas, J. QTL Analyses in Multiple Populations Employed for the Fine Mapping and Identification of Candidate Genes at a Locus Affecting Sugar Accumulation in Melon (Cucumis melo L.). Front. Plant Sci. 2017, 8, 1679. [Google Scholar] [CrossRef] [Green Version]
- Pereira, L.; Santo Domingo, M.; Argyris, J.; Mayobre, C.; Valverde, L.; Martín-Hernández, A.M.; Pujol, M.; Garcia-Mas, J. A Novel Introgression Line Collection to Unravel the Genetics of Climacteric Ripening and Fruit Quality in Melon. Sci. Rep. 2021, 11, 11364. [Google Scholar] [CrossRef]
- Castro, G.; Perpiñá, G.; Monforte, A.J.; Picó, B.; Esteras, C. New Melon Introgression Lines in a Piel de Sapo Genetic Background with Desirable Agronomical Traits from Dudaim Melons. Euphytica 2019, 215, 169. [Google Scholar] [CrossRef]
- Martínez-Martínez, C.; Gonzalo, M.J.; Sipowicz, P.; Campos, M.; Martínez-Fernández, I.; Leida, C.; Zouine, M.; Alexiou, K.G.; Garcia-Mas, J.; Gómez, M.D.; et al. A Cryptic Variation in a Member of the Ovate Family Proteins Is Underlying the Melon Fruit Shape QTL Fsqs8.1. Theor. Appl. Genet. 2022, 135, 785–801. [Google Scholar] [CrossRef]
- Pereira, L.; Santo Domingo, M.; Ruggieri, V.; Argyris, J.; Phillips, M.A.; Zhao, G.; Lian, Q.; Xu, Y.; He, Y.; Huang, S.; et al. Genetic Dissection of Climacteric Fruit Ripening in a Melon Population Segregating for Ripening Behavior. Hortic. Res. 2020, 7, 187. [Google Scholar] [CrossRef]
- Perpiñá, G.; Cebolla-Cornejo, J.; Esteras, C.; Monforte, A.J.; Picó, B. ‘MAK-10′: A Long Shelf-Life Charentais Breeding Line Developed by Introgression of a Genomic Region from Makuwa Melon. HortScience 2017, 52, 1633–1638. [Google Scholar] [CrossRef]
- Giordano, A.; Santo Domingo, M.; Quadrana, L.; Pujol, M.; Martín-Hernández, A.M.; Garcia-Mas, J. CRISPR/Cas9 Gene Editing Uncovers the Roles of CONSTITUTIVE TRIPLE RESPONSE 1 and REPRESSOR OF SILENCING 1 in Melon Fruit Ripening and Epigenetic Regulation. J. Exp. Bot. 2022, 73, 4022–4033. [Google Scholar] [CrossRef]
- Liu, B.; Santo Domingo, M.; Mayobre, C.; Martín-Hernández, A.M.; Pujol, M.; Garcia-Mas, J. Knock-Out of CmNAC-NOR Affects Melon Climacteric Fruit Ripening. Front. Plant Sci. 2022, 13, 878037. [Google Scholar] [CrossRef] [PubMed]
- Santo Domingo, M.; Areco, L.; Mayobre, C.; Valverde, L.; Martín-Hernández, A.M.; Pujol, M.; Garcia-Mas, J. Modulating Climacteric Intensity in Melon through QTL Stacking. Hortic. Res. 2022, 9, uhac131. [Google Scholar] [CrossRef] [PubMed]
- Eduardo, I.; Arús, P.; Monforte, A.J.; Obando, J.; Fernández-Trujillo, J.P.; Martínez, J.A.; Alarcón, A.L.; Álvarez, J.M.; Knaap, E. van der Estimating the Genetic Architecture of Fruit Quality Traits in Melon Using a Genomic Library of Near Isogenic Lines. J. Am. Soc. Hortic. Sci. 2007, 132, 80–89. [Google Scholar] [CrossRef] [Green Version]
- Leida, C.; Moser, C.; Esteras, C.; Sulpice, R.; Lunn, J.E.; de Langen, F.; Monforte, A.J.; Picó, B. Variability of Candidate Genes, Genetic Structure and Association with Sugar Accumulation and Climacteric Behavior in a Broad Germplasm Collection of Melon (Cucumis melo L.). BMC Genet. 2015, 16, 28. [Google Scholar] [CrossRef] [Green Version]
- Nimmakayala, P.; Tomason, Y.R.; Abburi, V.L.; Alvarado, A.; Saminathan, T.; Vajja, V.G.; Salazar, G.; Panicker, G.K.; Levi, A.; Wechter, W.P.; et al. Genome-Wide Differentiation of Various Melon Horticultural Groups for Use in GWAS for Fruit Firmness and Construction of a High Resolution Genetic Map. Front. Plant Sci. 2016, 7, 1437. [Google Scholar] [CrossRef] [Green Version]
- Périn, C.; Dogimont, C.; Giovinazzo, N.; Besombes, D.; Guitton, L.; Hagen, L.; Pitrat, M. Genetic Control and Linkages of Some Fruit Characters in Melon–The Cucurbit Genetics Cooperative (CGC). 1999. Available online: https://cucurbit.info/1999/07/genetic-control-and-linkages-of-some-fruit-characters-in-melon/ (accessed on 16 May 2022).
- Shen, J.; Xu, X.; Zhang, Y.; Niu, X.; Shou, W. Genetic Mapping and Identification of the Candidate Genes for Mottled Rind in Cucumis melo L. Front. Plant Sci. 2021, 12, 2563. [Google Scholar] [CrossRef]
- Kubicki, B. Inheritance of some characters in muskmelons (Cucumis melo). Genet. Pol. 1962, 3, 265–274. [Google Scholar]
- Feder, A.; Burger, J.; Gao, S.; Lewinsohn, E.; Katzir, N.; Schaffer, A.A.; Meir, A.; Davidovich-Rikanati, R.; Portnoy, V.; Gal-On, A.; et al. A Kelch Domain-Containing F-Box Coding Gene Negatively Regulates Flavonoid Accumulation in Muskmelon. Plant Physiol. 2015, 169, 1714–1726. [Google Scholar] [CrossRef] [Green Version]
- Zhao, G.; Lian, Q.; Zhang, Z.; Fu, Q.; He, Y.; Ma, S.; Ruggieri, V.; Monforte, A.J.; Wang, P.; Julca, I.; et al. A Comprehensive Genome Variation Map of Melon Identifies Multiple Domestication Events and Loci Influencing Agronomic Traits. Nat Genet 2019, 51, 1607–1615. [Google Scholar] [CrossRef] [PubMed]
- Monforte, A.J.; Oliver, M.; Gonzalo, M.J.; Alvarez, J.M.; Dolcet-Sanjuan, R.; Arús, P. Identification of Quantitative Trait Loci Involved in Fruit Quality Traits in Melon (Cucumis melo L.). Theor. Appl. Genet. 2004, 108, 750–758. [Google Scholar] [CrossRef] [PubMed]
- Cuevas, H.E.; Staub, J.E.; Simon, P.W.; Zalapa, J.E. A Consensus Linkage Map Identifies Genomic Regions Controlling Fruit Maturity and Beta-Carotene-Associated Flesh Color in Melon (Cucumis melo L.). Theor. Appl. Genet. 2009, 119, 741–756. [Google Scholar] [CrossRef] [PubMed]
- Galpaz, N.; Gonda, I.; Shem-Tov, D.; Barad, O.; Tzuri, G.; Lev, S.; Fei, Z.; Xu, Y.; Mao, L.; Jiao, C.; et al. Deciphering Genetic Factors That Determine Melon Fruit-Quality Traits Using RNA-Seq-Based High-Resolution QTL and EQTL Mapping. Plant J. 2018, 94, 169–191. [Google Scholar] [CrossRef] [Green Version]
- Tzuri, G.; Zhou, X.; Chayut, N.; Yuan, H.; Portnoy, V.; Meir, A.; Sa’ar, U.; Baumkoler, F.; Mazourek, M.; Lewinsohn, E.; et al. A ‘Golden’ SNP in CmOr Governs the Fruit Flesh Color of Melon (Cucumis melo). Plant J. 2015, 82, 267–279. [Google Scholar] [CrossRef]
- Harel-Beja, R.; Tzuri, G.; Portnoy, V.; Lotan-Pompan, M.; Lev, S.; Cohen, S.; Dai, N.; Yeselson, L.; Meir, A.; Libhaber, S.E.; et al. A Genetic Map of Melon Highly Enriched with Fruit Quality QTLs and EST Markers, Including Sugar and Carotenoid Metabolism Genes. Theor. Appl. Genet. 2010, 121, 511–533. [Google Scholar] [CrossRef]
- Ramamurthy, R.K.; Waters, B.M. Identification of Fruit Quality and Morphology QTLs in Melon (Cucumis melo) Using a Population Derived from Flexuosus and Cantalupensis Botanical Groups. Euphytica 2015, 204, 163–177. [Google Scholar] [CrossRef]
- Gur, A.; Tzuri, G.; Meir, A.; Sa’ar, U.; Portnoy, V.; Katzir, N.; Schaffer, A.A.; Li, L.; Burger, J.; Tadmor, Y. Genome-Wide Linkage-Disequilibrium Mapping to the Candidate Gene Level in Melon (Cucumis melo). Sci. Rep. 2017, 7, 9770. [Google Scholar] [CrossRef]
- Díaz, A.; Martín-Hernández, A.M.; Dolcet-Sanjuan, R.; Garcés-Claver, A.; Álvarez, J.M.; Garcia-Mas, J.; Picó, B.; Monforte, A.J. Quantitative Trait Loci Analysis of Melon (Cucumis melo L.) Domestication-Related Traits. Theor. Appl. Genet. 2017, 130, 1837–1856. [Google Scholar] [CrossRef]
- Barrantes, W.; López-Casado, G.; García-Martínez, S.; Alonso, A.; Rubio, F.; Ruiz, J.J.; Fernández-Muñoz, R.; Granell, A.; Monforte, A.J. Exploring New Alleles Involved in Tomato Fruit Quality in an Introgression Line Library of Solanum Pimpinellifolium. Front. Plant Sci. 2016, 7, 1172. [Google Scholar] [CrossRef] [Green Version]
- Snouffer, A.; Kraus, C.; van der Knaap, E. The Shape of Things to Come: Ovate Family Proteins Regulate Plant Organ Shape. Curr. Opin. Plant Biol. 2020, 53, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Moreno, E.; Obando, J.M.; Dos-Santos, N.; Fernández-Trujillo, J.P.; Monforte, A.J.; Garcia-Mas, J. Candidate Genes and QTLs for Fruit Ripening and Softening in Melon. Theor. Appl. Genet. 2008, 116, 589–602. [Google Scholar] [CrossRef] [PubMed]
- Vegas, J.; Garcia-Mas, J.; Monforte, A.J. Interaction between QTLs Induces an Advance in Ethylene Biosynthesis during Melon Fruit Ripening. Theor. Appl. Genet. 2013, 126, 1531–1544. [Google Scholar] [CrossRef]
- Ríos, P.; Argyris, J.; Vegas, J.; Leida, C.; Kenigswald, M.; Tzuri, G.; Troadec, C.; Bendahmane, A.; Katzir, N.; Picó, B.; et al. ETHQV6.3 Is Involved in Melon Climacteric Fruit Ripening and Is Encoded by a NAC Domain Transcription Factor. Plant J. 2017, 91, 671–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oren, E.; Tzuri, G.; Dafna, A.; Rees, E.R.; Song, B.; Freilich, S.; Elkind, Y.; Isaacson, T.; Schaffer, A.A.; Tadmor, Y.; et al. QTL Mapping and Genomic Analyses of Earliness and Fruit Ripening Traits in a Melon Recombinant Inbred Lines Population Supported by de Novo Assembly of Their Parental Genomes. Hortic. Res. 2022, 9, uhab081. [Google Scholar] [CrossRef]
- Doyle, J.J. Isolation of plant DNA from fresh tissue. Focus 1990, 12, 13–15. [Google Scholar]
- Lu, J.; Hou, J.; Ouyang, Y.; Luo, H.; Zhao, J.; Mao, C.; Han, M.; Wang, L.; Xiao, J.; Yang, Y.; et al. A Direct PCR–Based SNP Marker–Assisted Selection System (D-MAS) for Different Crops. Mol. Breed. 2020, 40, 9. [Google Scholar] [CrossRef]
- Sanseverino, W.; Hénaff, E.; Vives, C.; Pinosio, S.; Burgos-Paz, W.; Morgante, M.; Ramos-Onsins, S.E.; Garcia-Mas, J.; Casacuberta, J.M. Transposon Insertions, Structural Variations, and SNPs Contribute to the Evolution of the Melon Genome. Mol. Biol. Evol. 2015, 32, 2760–2774. [Google Scholar] [CrossRef] [Green Version]
- Brewer, M.T.; Lang, L.; Fujimura, K.; Dujmovic, N.; Gray, S.; van der Knaap, E. Development of a Controlled Vocabulary and Software Application to Analyze Fruit Shape Variation in Tomato and Other Plant Species. Plant Physiol. 2006, 141, 15–25. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 23 February 2022).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chr | Number of ILs | Introgression Size (bp) | Introgression Size * (cM) | ||||
|---|---|---|---|---|---|---|---|
| Average | Min | Max | Average | Min | Max | ||
| 1 | 4 | 15,012,079 | 7,083,403 | 23,611,517 | 49.5 | 37.7 | 58.3 |
| 2 | 4 | 12,741,987 | 2,293,836 | 23,903,255 | 57.1 | 14.7 | 100.6 |
| 3 | 2 | 11,857,082 | 6,062,824 | 17,651,340 | 53.2 | 36.8 | 69.7 |
| 4 | 3 | 17,188,567 | 11,842,362 | 24,802,312 | 69.6 | 65.5 | 75.1 |
| 5 | 4 | 9,353,072 | 2,799,579 | 22,919,604 | 45.5 | 24.9 | 63.3 |
| 6 | 4 | 9,574,343 | 2,430,563 | 27,849,153 | 38.2 | 21.9 | 66.7 |
| 7 | 3 | 19,189,571 | 8,232,872 | 25,979,214 | 81.8 | 67.5 | 100.8 |
| 8 | 4 | 12,919,496 | 1,895,546 | 25,002,865 | 47.8 | 12.9 | 92.1 |
| 9 | 2 | 23,045,254 | 20,847,233 | 25,243,276 | 85.6 | 61.7 | 109.5 |
| 10 | 2 | 15,007,760 | 5,108,239 | 24,907,280 | 70.3 | 54.0 | 86.6 |
| 11 | 2 | 25,057,113 | 20,321,128 | 29,793,099 | 89.6 | 54.0 | 99.4 |
| 12 | 2 | 13,781,830 | 4,579,847 | 22,983,813 | 51.9 | 47.1 | 56.8 |
| Total | 36 | 14,584,344 | 1,895,546 | 29,793,099 | 58.5 | 12.9 | 109.5 |
| Trait | QTL | IL | Year | Chr. | Genomic Interval (Mb) | IL Mean | PS Mean | % Difference a | p-Value |
|---|---|---|---|---|---|---|---|---|---|
| FW | FWQW7.1 | VED7.1 | 2020 | 7 | 2.62–20.73 | 1090.4 | 2175.6 | −50.0 | 0.0131 |
| VED7.2 | 2020 | 963.6 | −55.7 | 0.0148 | |||||
| FP | FPQW4.1 | VED4.2 | 2020 | 4 | 22.48–27.25 | 54.17 | 62.48 | −13.30 | 0.0183 |
| VED4.3 | 2020 | 54.58 | −12.64 | 0.0259 | |||||
| FPQW8.1 | VED8.1 | 2020 | 8 | 0–6.89 | 54.65 | −12.53 | 0.0053 | ||
| FPQW7.1 | VED7.1 | 2020 | 7 | 2.62–20.73 | 48.66 | −22.12 | 0.0004 | ||
| VED7.2 | 2020 | 48.08 | −23.05 | 0.0002 | |||||
| FPQW8.2 | VED8.3 | 2020 | 8 | 31.12–34.62 | 45.2 | −27.66 | 0.0002 | ||
| FPQW11.1 | VED11.2 | 2020 | 11 | 29.79–34.46 | 54.89 | −12.15 | 0.0071 | ||
| FPQW4.2 | VED4.1 | 2021 | 4 | 0–14.92 | 71.52 | 62.2 | 14.98 | 0.0216 | |
| FA | FAQW4.1 | VED4.2 | 2020 | 4 | 22.48–27.25 | 191.39 | 253.8 | −24.59 | 0.0336 |
| VED4.3 | 2020 | 193.09 | −23.92 | 0.0053 | |||||
| FAQW5.1 | VED5.3 | 2020 | 27.63–29.32 | 200.19 | −21.12 | 0.0203 | |||
| FAQW8.1 | VED8.1 | 2020 | 8 | 0–6.89 | 202.38 | −20.26 | 0.0163 | ||
| FAQW7.1 | VED7.1 | 2020 | 7 | 2.62–20.73 | 156.19 | −38.46 | 0.0006 | ||
| VED7.2 | 2020 | 150.7 | −40.62 | 0.0004 | |||||
| FAQW8.2 | VED8.3 | 2020 | 8 | 31.12–34.62 | 136.38 | −46.26 | 0.0002 | ||
| FAQW11.1 | VED11.2 | 2020 | 11 | 29.79–34.46 | 197.52 | −22.17 | 0.0097 | ||
| FL | FLQW5.1 | VED5.3 | 2020 | 5 | 27.63–29.32 | 18.3 | 21.66 | −15.51 | 0.0477 |
| FLQW6.1 | VED6.3 | 2020 | 6 | 7.47–35.32 | 18.1 | −16.44 | 0.0429 | ||
| FLQW8.1 | VED8.1 | 2020 | 8 | 0–6.89 | 18.33 | −15.37 | 0.0461 | ||
| FLQW7.1 | VED7.1 | 2020 | 7 | 2.62–20.73 | 16.71 | −22.85 | 0.0187 | ||
| VED7.2 | 2020 | 16.76 | −22.62 | 0.0167 | |||||
| FLQW8.2 | VED8.3 | 2020 | 8 | 31.12–34.62 | 15.02 | −30.66 | 0.0131 | ||
| FLQW11.1 | VED11.2 | 2020 | 11 | 29.79–34.46 | 17.04 | −21.33 | 0.0171 | ||
| FWI | FWIQW4.1 | VED4.3 | 2020 | 4 | 30.00–34.31 | 13.42 | 15.16 | −11.48 | 0.0473 |
| FWIQW7.1 | VED7.1 | 2020 | 7 | 2.62–20.73 | 12.07 | −20.38 | 0.0171 | ||
| VED7.2 | 2020 | 11.41 | −24.73 | 0.0130 | |||||
| FWIQW8.1 | VED8.3 | 2020 | 8 | 31.12–34.62 | 11.45 | −24.47 | 0.0179 | ||
| FS | FSQW6.1 | VED6.3 | 2020 | 6 | 7.47–35.32 | 1.19 | 1.43 | −16.78 | 0.0011 |
| FSQW11.1 | VED11.2 | 2020 | 11 | 29.79–34.46 | 1.15 | −19.58 | <0.0001 | ||
| FIR | FIRQW2.1 | VED2.2 | 2020 | 2 | 3.16–24.77 | 4.52 | 2.86 | 58.04 | 0.0298 |
| VED2.3 | 2020 | 4.59 | 60.49 | 0.0475 | |||||
| FIRQW8.1 | VED8.1 | 2020 | 8 | 0–6.89 | 0.88 | −69.23 | 0.0459 | ||
| FIRQW8.2 | VED8.2 | 2020 | 8 | 6.89–14.98 | 0.85 | −70.03 | 0.0334 | ||
| FIRQW8.3 | VED8.3 | 2020 | 8 | 14.96–34.62 | 0.62 | −78.32 | 0.0130 | ||
| FIRQW11.1 | VED11.2 | 2020 | 11 | 29.79–34.46 | 4.8 | 67.83 | 0.0171 | ||
| EARO | EAROQW6.1 | VED6.3 | 2020 | 6 | 7.47–35.32 | 49.5 | 55.36 | −10.59 | 0.0078 |
| EAROQW8.1 | VED8.1 | 2020 | 8 | 0–6.89 | 40.89 | −26.14 | <0.0001 | ||
| EAROQW8.2 | VED8.2 | 2020 | 8 | 6.89–14.98 | 46.1 | −16.73 | <0.0001 | ||
| EAROQW8.3 | VED7.1 | 2020 | 8 | 31.12–34.62 | 47.17 | −14.79 | 0.0063 | ||
| VED7.2 | 2020 | 46.8 | −15.46 | 0.0007 | |||||
| VED8.3 | 2020 | 46.9 | −15.28 | 0.0001 | |||||
| EYELL | EYELLQW1.1 | VED1.3 | 2020 | 1 | 29.95–34.47 | 29 | 26 | 0.12 | 0.0181 |
| VED1.4 | 2020 | 31 | 0.19 | 0.0076 | |||||
| EYELLQW8.1 | VED8.3 | 2020 | 8 | 25.00–29.53 | 37 | 0.42 | 0.0004 | ||
| EYELLQW10.1 | VED10.1 | 2020 | 10 | 1.76–5.11 | 43 | 0.65 | 0.0083 | ||
| VED10.2 | 2020 | 54 | 1.08 | 0.0011 | |||||
| EYELLQW10.1 | VED10.1 | 2021 | 10 | 1.76–5.11 | 56 | 27 | 1.07 | 0.0002 | |
| EALF | EALFQW6.1 | VED6.3 | 2020 | 6 | 7.47–35.32 | 49.38 | 55.81 | −11.52 | <0.0001 |
| EALFQW8.1 | VED8.1 | 2020 | 8 | 2.63–6.89 | 41.43 | −25.76 | <0.0001 | ||
| VED8.3 | 2020 | 44.56 | −20.16 | <0.0001 | |||||
| EALFQW8.2 | VED8.2 | 2020 | 8 | 6.89–14.98 | 47.33 | −15.19 | <0.0001 | ||
| EALFQW8.3 | VED7.1 | 2020 | 8 | 31.12–34.62 | 49.83 | −10.71 | 0.0016 | ||
| VED7.2 | 2020 | 48.6 | −12.92 | 0.0002 | |||||
| VED8.3 | 2020 | 44.56 | −20.16 | <0.0001 | |||||
| ABS | ABSQW6.1 | VED6.3 | 2020 | 6 | 7.47–35.32 | 1 | 0.11 | 0.89 | 0.0031 |
| ABSQW8.1 | VED8.1 | 2020 | 8 | 2.63–6.89 | 1.11 | 1.00 | 0.0008 | ||
| VED8.3 | 2020 | 2.44 | 2.33 | <0.0001 | |||||
| ABSQW8.2 | VED8.2 | 2020 | 8 | 6.89–14.98 | 1 | 0.89 | <0.0001 | ||
| ABSQW8.3 | VED7.1 | 2020 | 8 | 31.12–34.62 | 0.83 | 0.72 | 0.0369 | ||
| VED7.2 | 2020 | 1.2 | 1.09 | 0.0049 | |||||
| VED8.3 | 2020 | 2.44 | 2.33 | <0.0001 | |||||
| HAR | HARQW6.1 | VED6.3 | 2020 | 6 | 7.47–35.32 | 52.75 | 55.97 | −5.75 | <0.0001 |
| HARQW8.1 | VED8.1 | 2020 | 8 | 2.63–6.89 | 46.43 | −17.05 | <0.0001 | ||
| VED8.3 | 2020 | 47 | −16.02 | <0.0001 | |||||
| HARQW8.2 | VED8.2 | 2020 | 8 | 6.89–14.98 | 52.63 | −5.97 | 0.0002 | ||
| HARQW8.3 | VED7.2 | 2020 | 8 | 31.12–34.62 | 52 | −7.09 | <0.0001 | ||
| VED8.3 | 2020 | 47 | −16.02 | <0.0001 |
| Group | Phenotype | Units * | Symbol |
|---|---|---|---|
| Fruit quality traits | Mottled rind | MOT | |
| Yellowing of the rind | YELL | ||
| Presence of sutures | SUT | ||
| Flesh color | FC | ||
| External color of immature fruit | ECOL | ||
| Soluble solid content | ° Brix | SSC | |
| Firmness | kg cm−2 | FIR | |
| Fruit morphology traits | Fruit area | cm2 | FA |
| Fruit perimeter | cm | FP | |
| Fruit length | cm | FL | |
| Fruit width | cm | FWI | |
| Fruit shape | FS | ||
| Fruit weight | g | FWI | |
| Fruit ripening traits | Presence of aroma | DAP | EARO |
| Earliness of yellowing of the rind | DAP | EYELL | |
| Abscission layer formation | DAP | EALF | |
| Level of abscission | ** | ABS | |
| Harvest date | DAP | HAR |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santo Domingo, M.; Mayobre, C.; Pereira, L.; Argyris, J.; Valverde, L.; Martín-Hernández, A.M.; Garcia-Mas, J.; Pujol, M. Fruit Morphology and Ripening-Related QTLs in a Newly Developed Introgression Line Collection of the Elite Varieties ‘Védrantais’ and ‘Piel de Sapo’. Plants 2022, 11, 3120. https://doi.org/10.3390/plants11223120
Santo Domingo M, Mayobre C, Pereira L, Argyris J, Valverde L, Martín-Hernández AM, Garcia-Mas J, Pujol M. Fruit Morphology and Ripening-Related QTLs in a Newly Developed Introgression Line Collection of the Elite Varieties ‘Védrantais’ and ‘Piel de Sapo’. Plants. 2022; 11(22):3120. https://doi.org/10.3390/plants11223120
Chicago/Turabian StyleSanto Domingo, Miguel, Carlos Mayobre, Lara Pereira, Jason Argyris, Laura Valverde, Ana Montserrat Martín-Hernández, Jordi Garcia-Mas, and Marta Pujol. 2022. "Fruit Morphology and Ripening-Related QTLs in a Newly Developed Introgression Line Collection of the Elite Varieties ‘Védrantais’ and ‘Piel de Sapo’" Plants 11, no. 22: 3120. https://doi.org/10.3390/plants11223120