
The 5-Aminolevulinic Acid (5-ALA) Supplement Enhances PSII Photochemical Activity and Antioxidant Activity in the Late Growth Promotion of Pseudostellaria heterophylla
Abstract
:1. Introduction
2. Results
2.1. Effects of 5-ALA on the Growth of Tuberous Roots
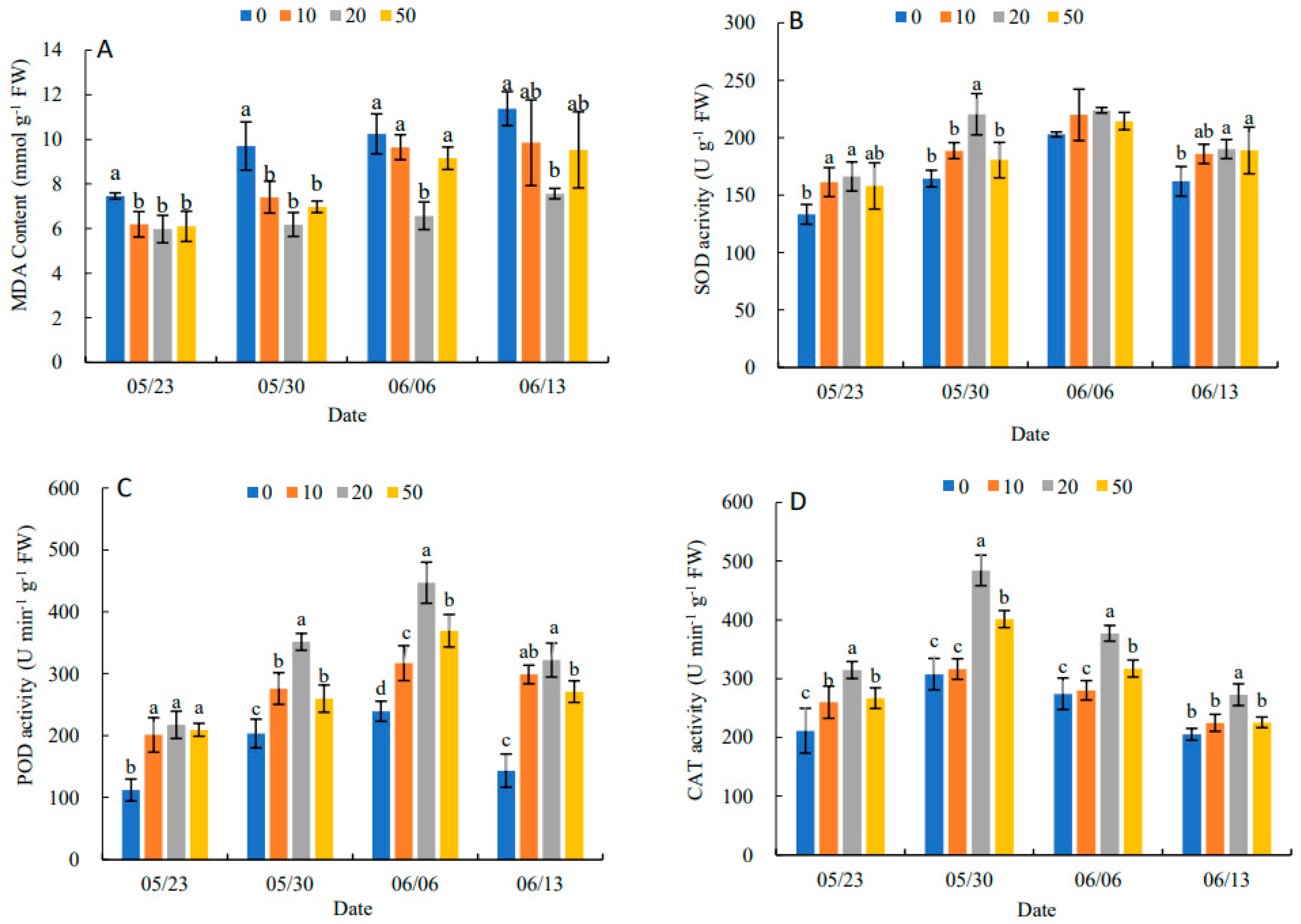
2.2. Effects of 5-ALA on Antioxidant Enzyme Activity
2.3. Effects on Photosynthetic Pigments in the Leaves
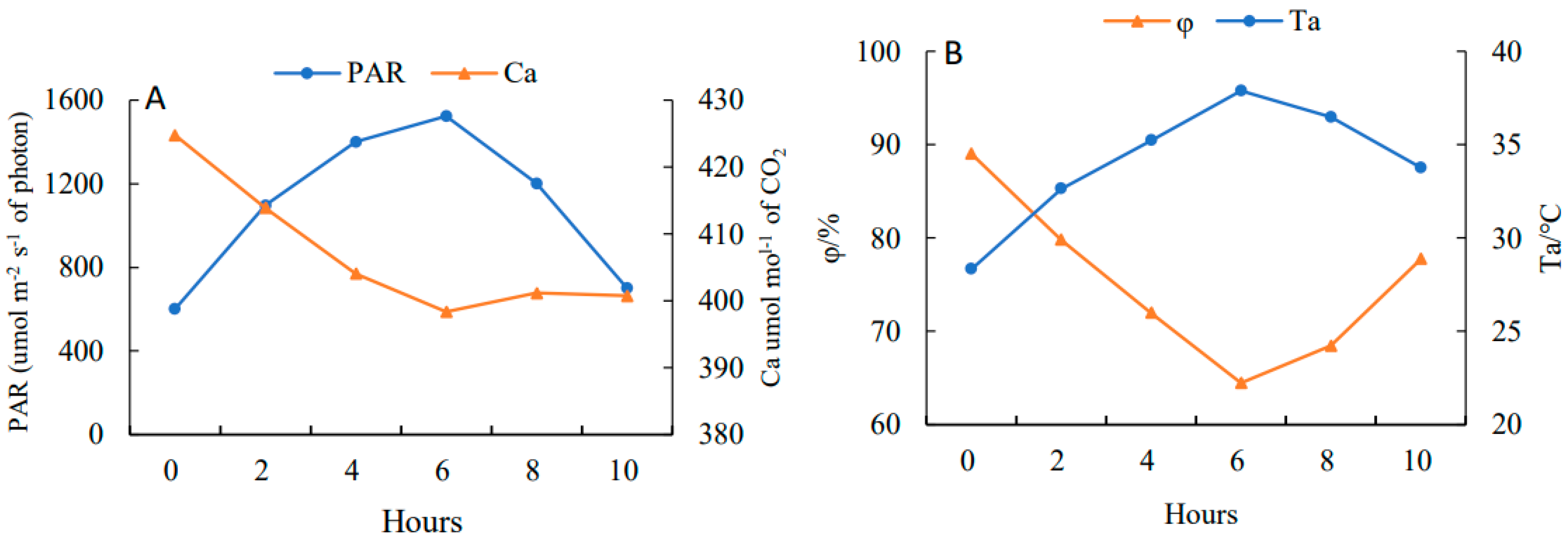
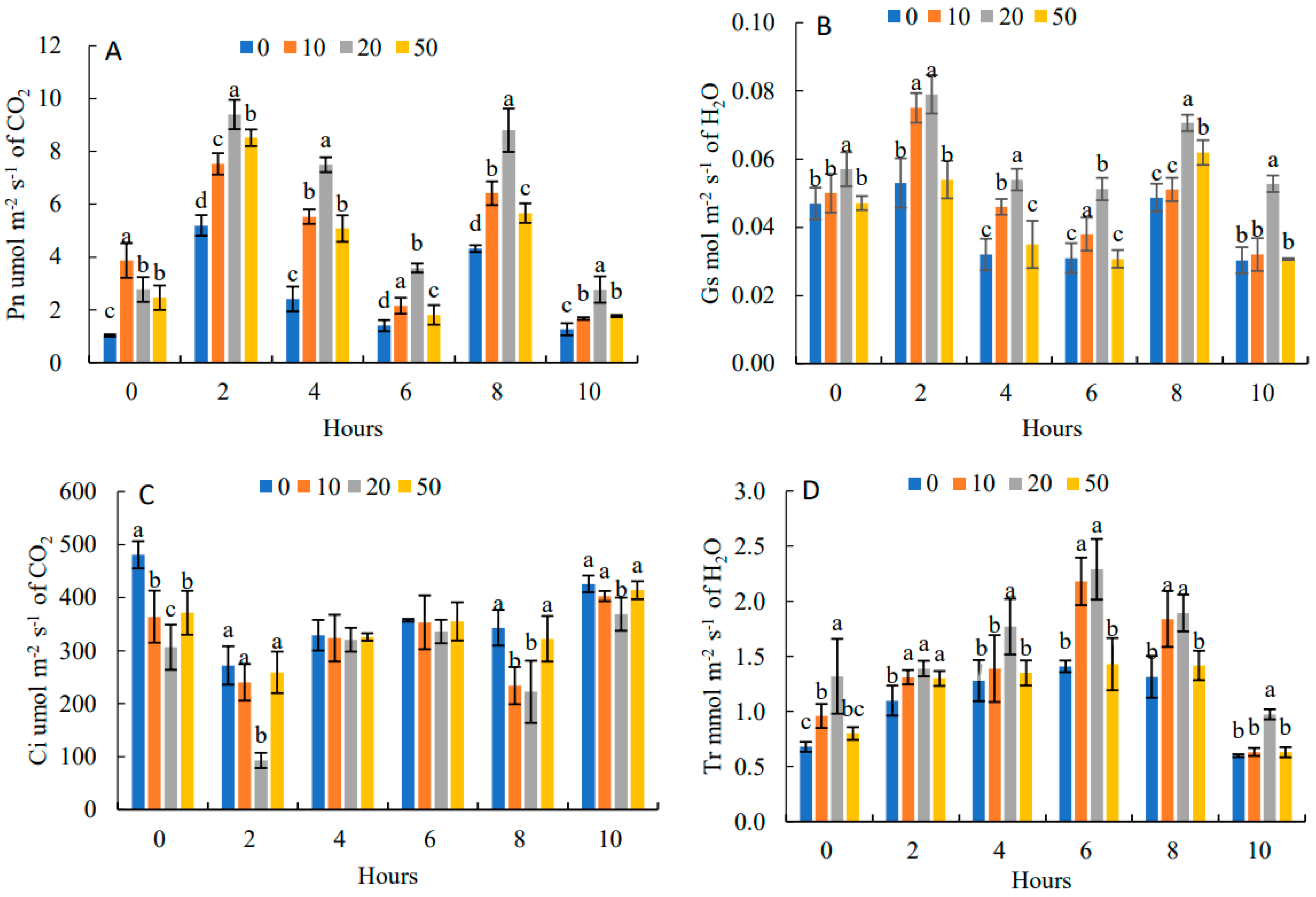
2.4. Effects on the Dynamic Parameters of Daily Photosynthetic Changes
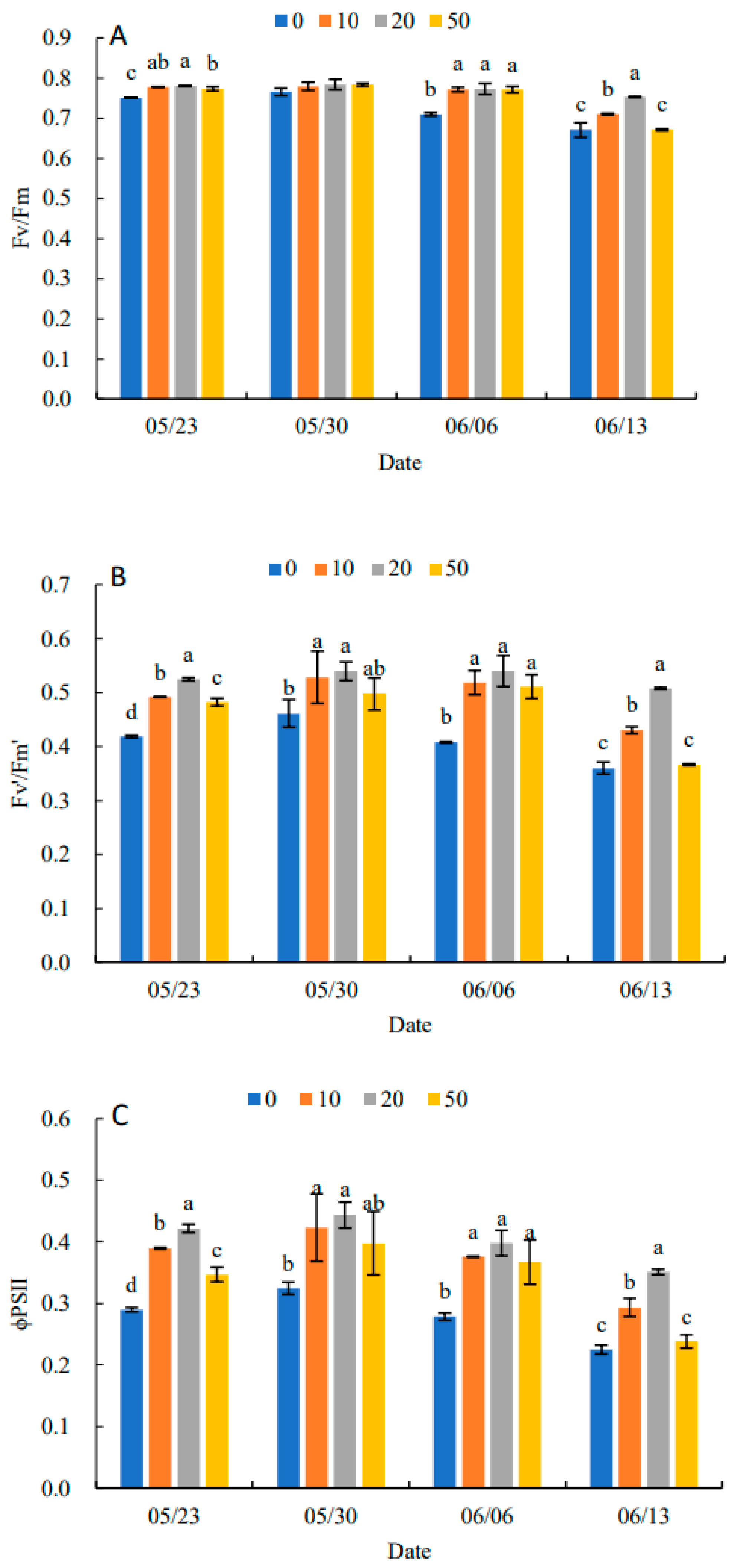
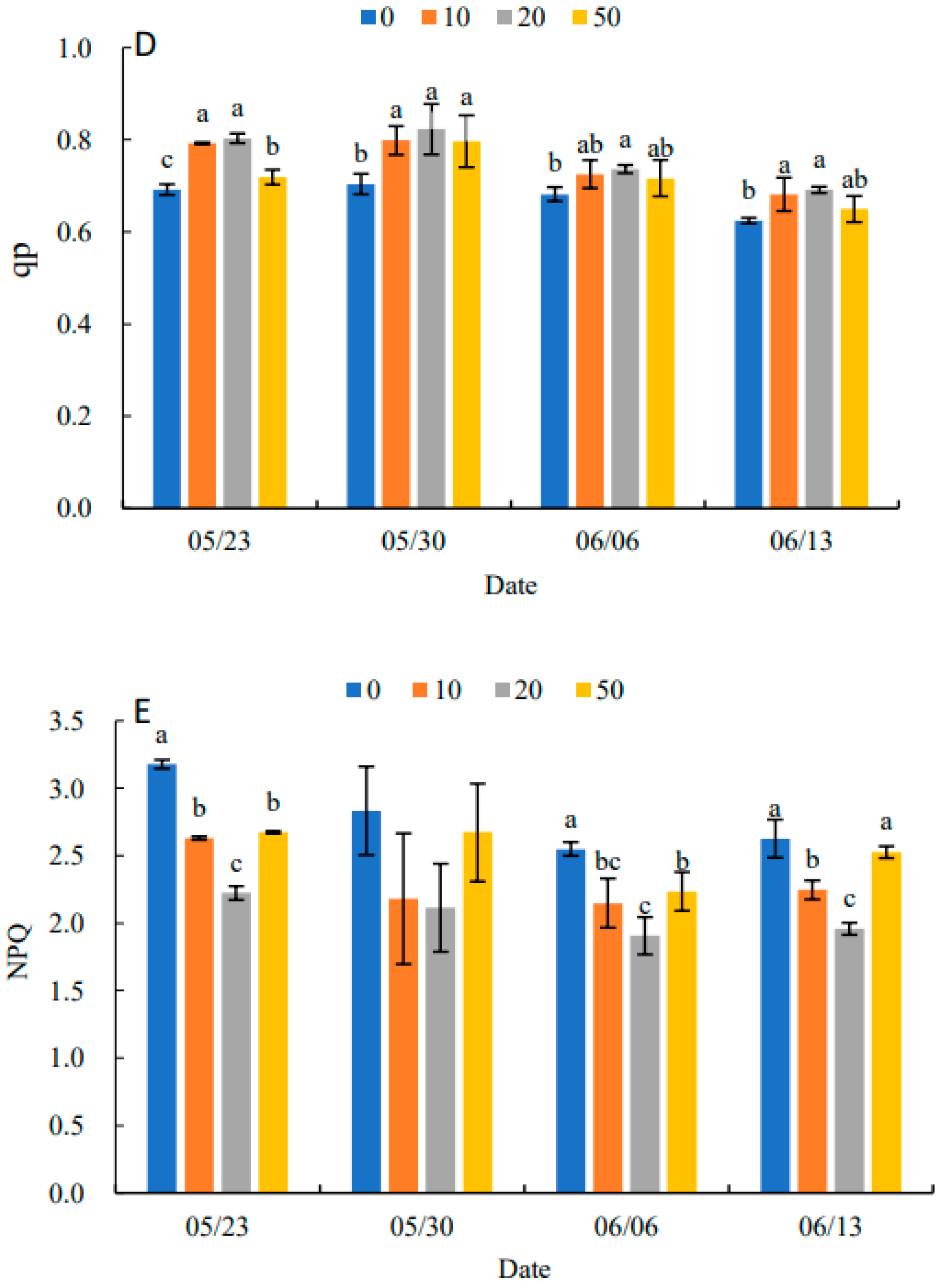
2.5. Effects on Dynamic Changes of Chlorophyll Fluorescence Parameters in Leaves
2.6. Effects on PSII Energy Partitioning and Specific Activity Parameters
2.7. Correlation Analysis
3. Discussion
4. Method and Materials
4.1. Sites and Materials
4.2. Experimental Design and Treatment
4.3. Measurement of Physiological Indicate
4.3.1. Growth Index
4.3.2. Antioxidant Enzyme Activity
4.3.3. Photosynthetic Pigment and Diurnal Variation Parameters of Leaf Photosynthesis
4.3.4. Measurement of Dynamic Changes in ChlorophyII Fluorescence Parameters
4.3.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Hua, Y.J.; Wang, C.C.; Wang, S.N.; Liu, Z.X.; Liu, X.H.; Zou, L.S.; Gu, W.; Luo, Y.Y.; Liu, J.X. iTRAQ-Based Quantitative Proteomic Analysis of Pseudostellaria heterophylla from Geo-Authentic Habitat and Cultivated Bases. Curr. Proteom. 2019, 16, 231–245. [Google Scholar] [CrossRef]
- Zhang, L.Y.; Guo, Z.W.; Hufang Gao, H.F.; Peng, X.Q.; Lin, W.X. Interaction of Pseudostellaria heterophylla with Quorum Sensing and Quorum Quenching Bacteria Mediated by Root Exudates in a Consecutive Monoculture System. J. Microbiol. Biotechnol. 2016, 26, 2159–2170. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.L.; Wang, X.R.; Zou, H.C.; Wang, T.; Chen, Y.; Xie, Y.F. Advances in chemical constituents in Pseudostellaria heterophylla. J. Anhui Agric. Univ. 2016, 43, 827–833. (In Chinese) [Google Scholar] [CrossRef]
- Lin, S.; Huangpu, J.J.; Chen, T.; Wu, L.K.; Lin, W.X. Analysis of soil microbial community structure and enzyme activities associated with negative effects of Pseudostellaria heterophylla consecutive monoculture on yield. Pak. J. Bot. 2015, 47, 761–769. [Google Scholar]
- Tan, S.Y.; Cao, J.; Xia, X.L.; Li, Z.H. Advances in 5-Aminolevulinic Acid Priming to Enhance Plant Tolerance to Abiotic Stress. Int. J. Mol. Sci. 2022, 23, 702. [Google Scholar] [CrossRef]
- Wu, Y.; Liao, W.B.; Dawuda, M.M.; Hu, L.; Yu, J. 5-Aminolevulinic acid (ALA) biosynthetic and metabolic pathways and its role in higher plants: A review. Plant Growth Regul. 2019, 87, 327. [Google Scholar] [CrossRef] [Green Version]
- Yang, N.; Li, Y.M.; Li, J.W.; Teng, R.M.; Chen, Y.; Wang, Y.H.; Zhuang, J. Effects of Exogenous 5-Aminolevulinic acid on the Chlorophyll Synthesis and Fluorescence Characteristics and Gene Expression of Key Enzymes in Tea Plants under Drought Stress. J. Tea Sci. 2022, 42, 187–199. (In Chinese) [Google Scholar] [CrossRef]
- Rhaman, M.S.; Imran, S.; Karim, M.; Chakrobortty, J.; Mahamud, A.; Sarker, P.; Ul-Arif, T.; Robin, A.H.K.; Ye, W.; Murata, Y.; et al. 5-aminolevulinic acid-mediated plant adaptive responses to abiotic stress. Plant Cell Rep. 2021, 40, 1451–1469. [Google Scholar] [CrossRef]
- Wei, Z.Y.; Zhang, Z.P.; Lee, M.R.; Sun, Y.P.; Wang, L.Y. Effect of 5-Aminolevulinic acid on leaf senescence and nitrogen metabolism of pakchoi under different nitrate levels. J. Plant Nutr. 2012, 35, 49–63. [Google Scholar] [CrossRef]
- Sunman, M.S.E.; Elradi, S.B.M.; Nimir, N.E.A.; Zhou, G.S.; Zhu, G.L.; Ibrahim, M.E.H.; Ali, A.Y.A. Foliar application of 5-aminolevulinic acid alleviated high temperature and drought stresses on wheat plants at seedling stage. Chil. J. Agric. Res. 2021, 81, 291–299. [Google Scholar] [CrossRef]
- Zhang, Z.J.; Li, H.Z.; Zhou, W.J.; Takeuchi, Y.; Yoneyama, K. Effect of 5-Aminolevulinic Acid on Development and Salt Tolerance of Potato (Solanum tuberosum L.) Microtubers in vitro. Plant Growth Regul. 2006, 49, 27–34. [Google Scholar] [CrossRef]
- Liang, H.Z.; Zhu, F.; Wang, R.J.; Huang, X.H.; Chu, J.J. Photosystem II of Ligustrum lucidum in response to different levels of manganese exposure. Sci. Rep. 2019, 9, 12568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Wang, D.Q.; Yao, Y.; Jin, X.F. Observation on biological characteristics of wild Pseudostellaria heterophylla. J. Chin. Med. Mater. 2011, 34, 1323–1328. [Google Scholar] [CrossRef]
- Kang, C.Z.; Zhou, T.; Jiang, W.K.; Guo, L.P.; Zhang, X.B. Research on quality regionalization of cultivated Pseudostellaria heterophylla based on climate factors. China J. Chin. Mater. Med. 2016, 41, 2386–2390. [Google Scholar] [CrossRef]
- Ji, H.J.; Jia, H.X.; Zhang, X.L.; Hu, J.J. Effect of drought stress on photosynthetic diurnal course and growth of Salix purpurea. J. Nanjing For. Univ. (Nat. Sci. Ed. ) 2016, 40, 41–46. [Google Scholar]
- Saeed, T.; Hassan, I.; Abbasi, N.A.; Jilani, G. Antioxidative activities and qualitative changes in gladiolus cut flowers in response to salicylic acid application. Sci. Hortic. 2016, 210, 236–241. [Google Scholar] [CrossRef]
- Yang, J.H.; Gao, Y.; Li, Y.M.; Qi, X.H.; Zhang, M.F. Salicylic acid-induced enhancement of cold tolerance through activation of antioxidative capacity in watermelon. Sci. Hortic. 2008, 118, 200–205. [Google Scholar]
- Heyneck, E.; Fernie, A.R. Metabolic regulation of photosynthesis. Biochem. Soc. Trans. 2018, 46, 321–328. [Google Scholar] [CrossRef]
- Wang, P.; Liu, Z.C.; Luo, S.L.; Li, J.; Zhang, J.; Li, L.; Xie, J. 5-Aminolevulinic acid and hydrogen sulphide alleviate chilling stress in pepper (Capsicum annuum L.) seedlings by enhancing chlorophyII synthesis pathway. Plant Physiol. Biochem. 2021, 167, 567–576. [Google Scholar] [CrossRef]
- Wang, C.K.; Guo, M. Effect of 5-Aminolevulinic Acid on Growth and Antioxidant Enzyme Activities of Avena nuda under Normal and High Temperature Conditions. Henan Agric. Sci. 2017, 46, 30–34. (In Chinese) [Google Scholar] [CrossRef]
- Christen, D.; Schönmann, S.; Jermini, M.; Strasser, R.J.; Défago, G. Characterization and early detection of grapevine (Vitis vinifera) stress responses to esca disease by in situ chlorophyll fluorescence and comparison with drought stress. Environ. Exp. Bot. 2007, 60, 504–514. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Sharkey, T.D. Stomatal conductance and photosynthesis. Annu. Rev. Plant Physiol. 1982, 33, 317–345. [Google Scholar] [CrossRef]
- Chen, J.H.; Wang, S.Q.; Chen, B.; Li, Y.; Amir, M.; Ma, L.; Zhu, K.; Yang, F.T.; Wang, X.B.; Liu, Y.Y.; et al. Comparative Analysis on the Estimation of Diurnal Solar-Induced Chlorophyll Fluorescence Dynamics for a Subtropical Evergreen Coniferous Forest. Remote Sens. 2021, 13, 1343. [Google Scholar] [CrossRef]
- Xu, Y.F.; Chu, C.C.; Yao, S.G. The impact of high-temperature stress on rice: Challenges and solutions. Crop J. 2021, 9, 963–976. [Google Scholar] [CrossRef]
- Li, Y.T.; Xu, W.W.; Ren, B.Z.; Zhao, B.; Wang, J.W.; Liu, P.; Zhang, Z.S. High temperature reduces photosynthesis in maize leaves by damaging chloroplast ultrastructure and photosystem II. J. Agron. Crop Sci. 2020, 206, 548–564. [Google Scholar] [CrossRef]
- Baker, N.R. ChlorophyII fluorescence: A probe of photosynthesis in vivo. Annu. Rev. Plant Biol. 2008, 59, 89–113. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Wang, Z.B.; Nie, Y.T.; Wang, R.H.; Guo, Y.P.; Zhao, Z.Y.; Mei, L.X. Alleviation of Photoinhibition by Methyl Jasmonate and Brassionlide under Strong Light in Apple Leaves. Acta Agric. Boreali-Occident. Sin. 2017, 26, 906–915. [Google Scholar]
- Zavafer, A.; Mancilla, C. Concepts of photochemical damage of Photosystem II and the role of excessive excitation. J. Photochem. Photobiol. C-Photochem. Rev. 2021, 47, 100421. [Google Scholar] [CrossRef]
- Zhang, J.L.; Chen, H.P.; Cheng, D.; Li, Y.L. Significance of Photoinhibition Response Mechanism of Sabina vulgaris Ant. in the Mu US Sandland. Arid. Zone Res. 2018, 35, 882–890. [Google Scholar] [CrossRef]
- Strasser, R.J.; Tsimilli-Michael, M.; Srivastava, A. Analysis of the Chlorophyll a Fluorescence Transient: ChlorophyII a Fluorescence; Springer: Berlin/Heidelberg, Germany, 2004; Volume 19, pp. 321–336. [Google Scholar]
- Hu, W.H.; Yan, X.H.; Li, X.H.; Cao, Z.G. Effect of 24-Epibrassionolide on the ChlorophyII Fluorescence Triansient in leaves of Pepper under drought stress. Bull. Bot. Res. 2021, 41, 53–59. [Google Scholar]
- Appenroth, K.J.; Stöckel, J.; Srivastava, A.; Strasser, R.J. Multiple effects of chromate on the photosynthetic apparatus of Spirodela polyrhiza as probed by OJIP chlorophyII a fluorescence measurement. Environ. Pollut. 2001, 115, 49–64. [Google Scholar] [CrossRef]
- Ma, Y.L.; Gu, H.; Zhang, Y.F.; Dai, L.; Li, J.; Jiang, F.L.; Xia, D.J.; Xie, Y.F. Effect of cerium nitrate on growth and photosynthetic characteristics of Pseudostellaria heterophylla. Nanjing For. Univ. (Nat. Sci.) 2017, 41, 71–79. (In Chinese) [Google Scholar]
- Han, W.X.; Jiang, H.; Bian, J.; Yun, J.H.; Sun, Y.Y.; Zhang, W.X.; Peng, Y. Leaf color change and its correlation with pigment content in 10 ornamental crabapple varieties in spring. J. Zhejiang Univ. (Agric. Life Sci.) 2020, 46, 562–570. [Google Scholar]
- Dai, Y.; Yuan, L.Y.; Zhang, S.J.; Wang, J.; Xie, S.L.; Zhao, M.R.; Chen, G.H.; Sun, R.F.; Wang, C.G. Comprehensive Evaluation for Cold Tolerance in Wucai (Brassica campestris L.) by the Performance Index on an Absorption Basis (PIabs). Agronomy 2019, 9, 61. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 5-ALA Concentration (mg·L−1) | Tuberous Root | Fresh Root | Aerial Part Weight (g) | Underground Part Weight (g) | Biomass Allocation (g) | Root-Shoot Ratio | |
|---|---|---|---|---|---|---|---|
| Root Length (cm) | Root Diameter (mm) | Fresh Root Yield (gm−2) | |||||
| 0 | 3.45 ± 0.47 c | 4.78 ± 0.37 c | 302.97 ± 7.34 c | 1.99 ± 0.99 b | 1.52 ± 0.05 c | 3.51 ± 0.76 c | 0.76 ± 0.06 b |
| 10 | 3.89 ± 0.30 b | 5.01 ± 0.39 b | 357.04 ± 10.34 b | 2.16 ± 0.08 a | 1.80 ± 0.12 b | 3.96 ± 0.83 b | 0.83 ± 0.06 b |
| 20 | 4.48 ± 0.33 a | 6.35 ± 0.73 a | 418.47 ± 15.66 a | 2.19 ± 0.09 a | 2.20 ± 0.10 a | 4.39 ± 1.01 a | 1.01 ± 0.07 a |
| 50 | 3.82 ± 0.32 b | 5.91 ± 0.41 b | 368.04 ± 11.69 b | 2.19 ± 0.11 a | 1.86 ± 0.11 b | 4.05 ± 085 b | 0.85 ± 0.08 b |
| φPo | Ψo | φEo | φDo | ABS/RC | TRo/RC | ETo/RC | |
|---|---|---|---|---|---|---|---|
| 0 | 0.60 ± 0.04 a | 0.38 ± 0.03 a | 0.23 ± 0.01 a | 0.41 ± 0.04 a | 4.18 ± 0.68 b | 2.47 ± 0.27 a | 0.95 ± 0.16 a |
| 20 | 0.73 ± 0.03 a | 0.44 ± 0.04 a | 0.32 ± 0.02 a | 0.27 ± 0.03 b | 3.21 ± 0.31 a | 2.45 ± 0.31 a | 1.04 ± 0.11 a |
| DIo/RC | ABS/CSm | TRo/CSm | ETo/CSm | DIo/CSo | RC/CSo | PIabs | |
| 0 | 1.71 ± 0.41 a | 1200.67 ± 376.97 a | 705.33 ± 189.41 a | 271.67 ± 84.89 a | 495.33 ± 188.50 a | 100.69 ± 2.67 a | 0.23 ± 0.05 b |
| 20 | 0.85 ± 0.02 b | 1384.33 ± 179.27 a | 1016 ± 153.91 a | 450.33 ± 81.19 a | 368.33 ± 42.06 a | 95.31 ± 1.74 b | 0.69 ± 0.08 a |
| Yield | Biomass Allocation | Pn | Gs | Ci | Tr | MDA | SOD | POD | CAT | Fv/Fm | Fv′/Fm′ | φPSII | qP | NPQ | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Yield | 1 | ||||||||||||||
| Biomass allocation | 0.998 ** | 1 | |||||||||||||
| Pn | 0.947 * | 0.944 * | 1 | ||||||||||||
| Gs | 0.802 | 0.775 | 0.913 | 1 | |||||||||||
| Ci | −0.927 | −0.914 | −0.998 * | −0.962 * | 1 | ||||||||||
| Tr | 0.684 | 0.666 | 0.871 | 0.962 * | −0.907 | 1 | |||||||||
| MDA | −0.995 ** | −0.986 * | −0.942 | −0.835 | 0.938 | 0.705 | 1 | ||||||||
| SOD | 0.880 | 0.909 | 0.842 | 0.548 | −0.753 | 0.512 | −0.827 | 1 | |||||||
| POD | 0.906 | 0.925 | 0.940 | 0.727 | −0.876 | 0.719 | −0.867 | 0.964 * | 1 | ||||||
| CAT | 0.951 * | 0.929 | 0.918 | 0.898 | −0.946 | 0.758 | −0.977 * | 0.695 | 0.773 | 1 | |||||
| Fv/Fm | 0.808 | 0.781 | 0.912 | 1.000 ** | −0.962 * | 0.955 * | −0.842 | 0.548 | 0.724 | 0.906 | 1 | ||||
| Fv′/Fm′ | 0.830 | 0.804 | 0.925 | 0.999 ** | −0.971 * | 0.948 | −0.862 | 0.575 | 0.743 | 0.920 | 0.999 ** | 1 | |||
| φPSII | 0.847 | 0.826 | 0.947 * | 0.996 ** | −0.983 * | 0.957 * | −0.872 | 0.625 | 0.788 | 0.915 | 0.995 ** | 0.997 ** | 1 | ||
| qP | 0.850 | 0.852 | 0.972 * | 0.900 | −0.954 * | 0.923 | −0.838 | 0.801 | 0.931 | 0.812 | 0.895 | 0.901 | 0.932 | 1 | |
| NPQ | −0.863 | −0.845 | −0.961 * | −0.990 ** | 0.989 * | −0.954 * | 0.884 | −0.659 | −0.816 | −0.917 | −0.989 * | −0.993 ** | −0.999 ** | 0.964 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, J.; Sun, M.; Qiu, L.; Xie, Y.; Ma, Y.; Liang, W. The 5-Aminolevulinic Acid (5-ALA) Supplement Enhances PSII Photochemical Activity and Antioxidant Activity in the Late Growth Promotion of Pseudostellaria heterophylla. Plants 2022, 11, 3035. https://doi.org/10.3390/plants11223035
Ma J, Sun M, Qiu L, Xie Y, Ma Y, Liang W. The 5-Aminolevulinic Acid (5-ALA) Supplement Enhances PSII Photochemical Activity and Antioxidant Activity in the Late Growth Promotion of Pseudostellaria heterophylla. Plants. 2022; 11(22):3035. https://doi.org/10.3390/plants11223035
Chicago/Turabian StyleMa, Julin, Meng Sun, Lingling Qiu, Yinfeng Xie, Yingli Ma, and Wenchao Liang. 2022. "The 5-Aminolevulinic Acid (5-ALA) Supplement Enhances PSII Photochemical Activity and Antioxidant Activity in the Late Growth Promotion of Pseudostellaria heterophylla" Plants 11, no. 22: 3035. https://doi.org/10.3390/plants11223035