
Effects of Phenolic Compounds on Walnut Bacterial Blight in the Green Husk of Hungarian-Bred Cultivars

 , ,
, ,
Abstract
:1. Introduction
2. Results
Phenolic Compounds
3. Discussion
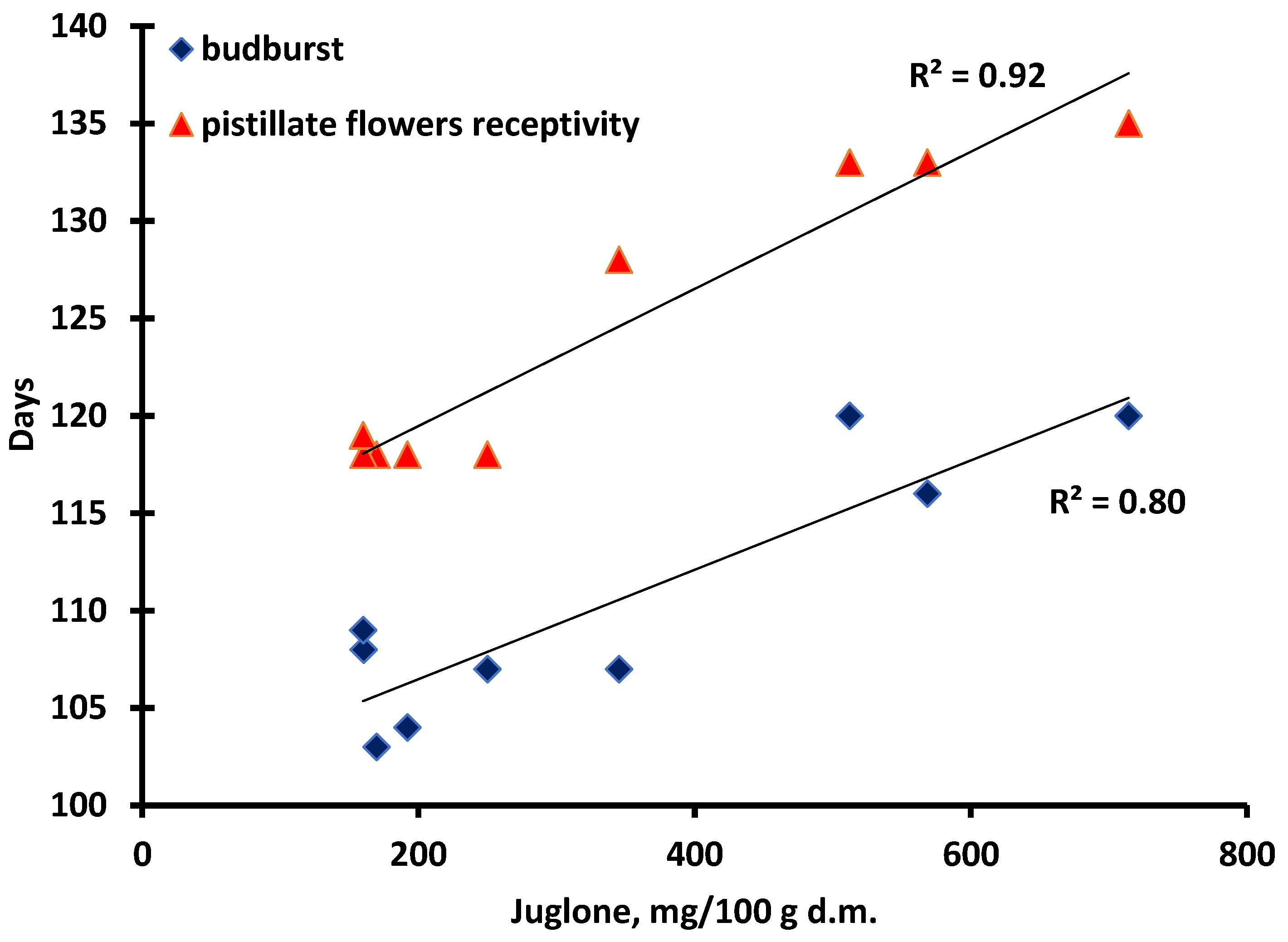
3.1. Phenolic Compounds and Early Spring Phenology
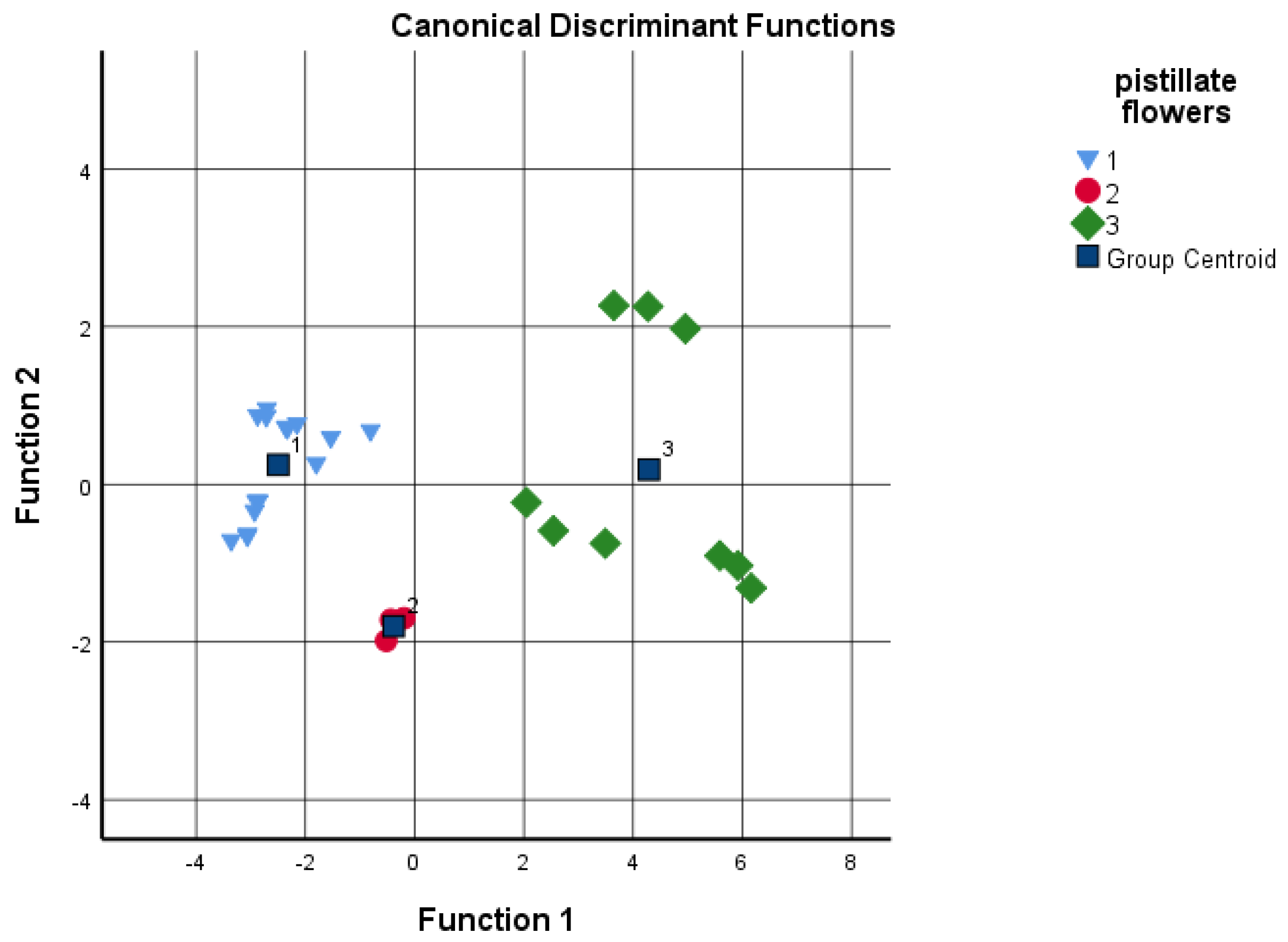
3.2. Tolerance/Susceptivity to Xanthomonas Arboricola pv. Juglandis
4. Materials and Methods
4.1. Plant Material
4.2. Phenological Stages
4.3. Sample Collection and Preparation
4.4. Chemicals
4.5. Total Phenolic Content
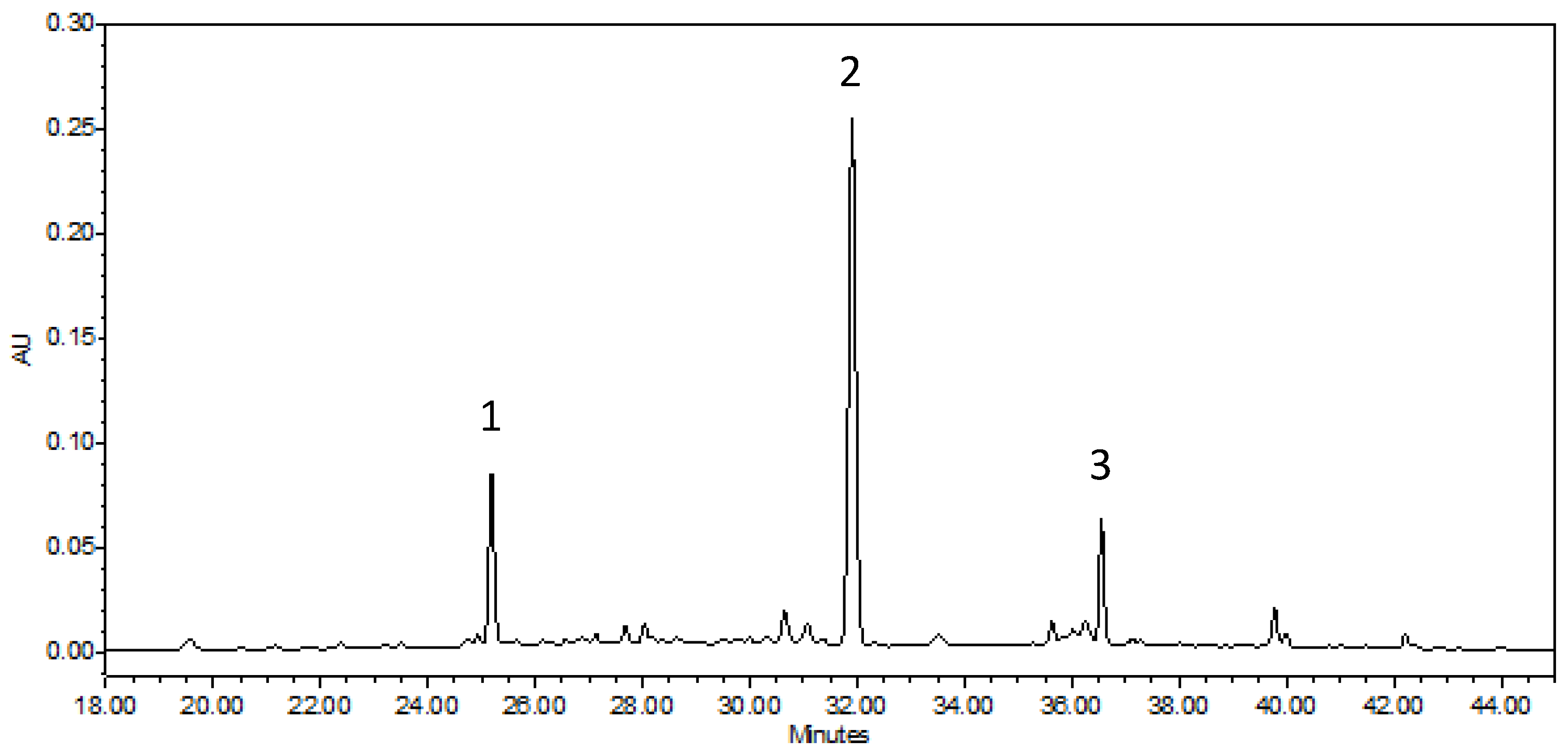
4.6. Phenolic Compounds’ Determination by HPLC-ESI-DAD
4.7. Walnut Blight Immunity Test
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rahimipanah, M.; Hamedi, M.; Mirzapour, M. Antioxidant activity and phenolic contents of Persian walnut (Juglans regia L.) green husk extract. Afr. J. Food Sci. Technol. 2010, 4, 105–111. [Google Scholar]
- Al-Nadaf, A.H.; Bastoni, H.M.; Hamdan, D.F. Microwave-assisted efficient extraction of phenolics from Juglans regia L.: Pellicle; kernel unripe fruits; and leaves in different solvents. Int. J. Green Pharm. 2018, 3, 182–190. [Google Scholar] [CrossRef]
- Marius, R.E.; Gheldiu, A.-M.; Moldovan, A.M.C.; Popa, D.-S.; Tomuta, I.; Vlase, L. Process Optimization for Improved Phenolic Compounds Recovery from Walnut (Juglans regia L.) Septum: Phytochemical Profile and Biological Activities. Molecules 2018, 11, 2814. [Google Scholar] [CrossRef] [Green Version]
- Acquaviva, R.; D’Angeli, F.; Malfa, G.A.; Ronsisvalle, S.; Garozzo, A.; Stivala, A.; Ragusa, S.; Nicolosi, D.; Salmeri, M.; Genovese, C. Antibacterial and anti-biofilm activities of walnut pellicle extract (Juglans regia L.) against coagulase-negative staphylococci. Nat. Prod. Res. 2021, 12, 2076–2081. [Google Scholar] [CrossRef]
- La Torre, C.; Caputo, P.; Plastina, P.; Cione, E.; Fazio, A. Green Husk of Walnuts (Juglans regia L.) from Southern Italy as a Valuable Source for the Recovery of Glucans and Pectins. Fermentation 2021, 7, 305. [Google Scholar] [CrossRef]
- Croitoru, A.; Ficai, D.; Craciun, L.; Ficai, A.; Andronescu, E. Evaluation and Exploitation of Bioactive Compounds of Walnut, Juglans regia. Curr. Pharm. Des. 2019, 2, 119–131. [Google Scholar] [CrossRef]
- Soto-Madrid, D.; Gutiérrez-Cutiño, M.; Pozo-Martínez, J.; Zúñiga-López, M.C.; Olea-Azar, C.; Matiacevich, S. Dependence of the Ripeness Stage on the Antioxidant and Antimicrobial Properties of Walnut (Juglans regia L.) Green Husk Extracts from Industrial By-Products. Molecules 2021, 10, 2878. [Google Scholar] [CrossRef]
- Oliveira, I.; Sousa, A.; Ferreira, I.C.; Bento, A.; Estevinho, L.; Pereira, J.A. Total phenols, antioxidant potential and antimicrobial activity of walnut (Juglans regia L.) green husks. Food Chem. Toxicol. 2008, 7, 2326–2331. [Google Scholar] [CrossRef]
- Carvalho, M.; Ferreira, P.J.; Mendes, V.S.; Silva, R.; Pereira, J.A.; Jerónimo, C.; Silva, B.M. Human cancer cell antiproliferative and antioxidant activities of Juglans regia L. Food Chem. Toxicol. 2010, 1, 441–447. [Google Scholar] [CrossRef]
- Cosmulesscu, S.N.; Trandafir, I.; Achim, G.; Botu, M.; Baciu, A.; Gruia, M. Phenolics of Green Husk in Mature Walnut Fruits. Not. Bot. Horti Agrobot. 2010, 1, 53–56. [Google Scholar] [CrossRef]
- Popovici, C.; Gîtin, L.; Alexe, P. Characterization of walnut (Juglans regia L.) green husk extract obtained by supercritical carbon dioxide fluid extraction. J. Food Packag. Sci. Tech. Technol. 2012, 1, 5–9. [Google Scholar]
- Ding, C.-B.; Wu, S.-Z.; Li, G.-Q.; Jia, C.-H.; Chang, L.-X. Anti-fungal activity of walnut green husk extracts. Mod. Food Sci. Technol. 2013, 29, 722–724, 755. [Google Scholar]
- Zhang, W.X.; He, K.Z.; Pu, Q. Antimicrobial and antioxidant activities of extracts from walnut green husks. Ying Yong Yu Huan Jing Sheng Wu Xue Bao 2014, 1, 87–92. [Google Scholar] [CrossRef]
- Salejda, A.; Janiewicz, U.; Korzeniowska, M.; Kolniak-Ostek, J.; Grazyna, K. Effect of walnut green husk addition on some quality properties of cooked sausages. LWT Food Sci. Technol. 2015, 61, 751–757. [Google Scholar] [CrossRef]
- Panth, N.; Paudel, K.R.; Karki, R. Phytochemical profile and biological activity of Juglans regia. J. Integr. Med. 2016, 5, 359–373. [Google Scholar] [CrossRef]
- Wianowska, D.; Garbaczewska, S.; Cieniecka-Roslonkiewicz, A.; Dawidowicz, A.L.; Jankowska, A. Comparison of antifungal activity of extracts from different Juglans regia cultivars and juglone. Microb. Pathog. 2016, 100, 263–267. [Google Scholar] [CrossRef]
- Sadeghi-Kiakhani, M.; Tehrani-Bagha, A.; Gharanjig, K.; Hashemi, E. Use of Pomegranate peels and Walnut Green husks as the green antimicrobial agents to reduce the consumption of inorganic nanoparticles on wool yarns. J. Clean. Prod. 2019, 231, 1463–1473. [Google Scholar] [CrossRef]
- Wianowska, D.; Garbaczewska, S.; Cieniecka-Roslonkiewicz, A.; Dawidowicz, A.; Typek, R.; Kielczewska, A. Influence of the Extraction Conditions on the Antifungal Properties of Walnut Green Husk Isolates. Anal. Lett. 2020, 53, 1970–1981. [Google Scholar] [CrossRef]
- Dehghani, S.; Nouri, M.; Baghi, M. The effect of adding walnut green husk extract on antioxidant and antimicrobial properties of ketchup. J. Food Bioprocess Eng. 2020, 2, 93–100. [Google Scholar]
- Medic, A.; Jakopič, J.; Hudina, M.; Solar, A.; Veberic, R. Identification and quantification of major phenolic constituents in Juglans regia L. leaves: Healthy vs. infected leaves with Xanthomonas campestris pv. juglandis using HPLC-MS/MS. J. King Saud Univ.-Sci. 2022, 34, 101890. [Google Scholar] [CrossRef]
- Colarič, M.; Veberic, R.; Solar, A.; Hudina, M.; Stampar, F. Phenolic Acids, Syringaldehyde, and Juglone in Fruits of Different Cultivars of Juglans regia L. J. Agric. Food Chem. 2005, 53, 6390–6396. [Google Scholar] [CrossRef] [PubMed]
- Hama, J.; Anwar, R.; Rashid, R.; Mohammed, N.; Thoss, V. The Diversity of Phenolic Compounds along Defatted Kernel, Green Husk and Leaves of Walnut (Juglans regia L.). Anal. Chem. Lett. 2016, 1, 35–46. [Google Scholar] [CrossRef]
- Kamran, G.; Yousef, G.; Abdollah, E.; Seyed, M.N.; Seyed, F.N.; Mohammad, A.E.; Fereshteh, P. Influence of environmental factors on antioxidant activity, phenol and flavonoids contents of walnut (Juglans regia L.) green husks. J. Med. Plants Res. 2011, 5, 1128–1133. [Google Scholar]
- Cosmulescu, S.; Trandafir, I.; Achim, G.; Baciu, A. Juglone Content in Leaf and Green Husk of Five Walnut (Juglans regia L.) Cultivars. Not. Bot. Horti Agrobot. 2011, 39, 237–240. [Google Scholar] [CrossRef] [Green Version]
- Akbari, V.; Jamei, R.; Heidari, R.; Esfahlan, A.J. Antiradical activity of different parts of Walnut (Juglans regia L.) fruit as a function of genotype. Food Chem. 2012, 135, 2404–2410. [Google Scholar] [CrossRef]
- Rahmani, F.; Dehganiasl, M.; Heidari, R.; Rezaee, R.; Darvishzadeh, R. Genotype impact on antioxidant potential of hull and kernel in Persian walnut (Juglans regia L.). Int. Food Res. J. 2018, 25, 35–42. [Google Scholar]
- Jahanban-Esfahlan, A.; Ostadrahimi, A.; Tabibiazar, M.; Amarowicz, R. A Comprehensive Review on the Chemical Constituents and Functional Uses of Walnut (Juglans spp.) Husk. Int. J. Mol. Sci. 2019, 20, 3920. [Google Scholar] [CrossRef] [Green Version]
- Jakopic, J.; Solar, A.; Colaric, M.; Hudina, M.; Veberic, R.; Stampar, F. The influence of ethanol concentration on content of total and individual phenolics in walnut alcoholic drink. Acta Aliment. 2008, 37, 233–239. [Google Scholar] [CrossRef]
- Jakopic, J.; Veberic, R. Extraction of phenolic compounds from green walnut fruits in different solvents. Acta Agric. Slov. 2009, 93, 11–15. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Agulló, A.; Pereira, E.; Freire, M.S.; Valentão, P.; Andrade, P.B.; González-Álvarez, J.; Pereira, J.A. Influence of solvent on the antioxidant and antimicrobial properties of walnut (Juglans regia L.) green husk extracts. Ind. Crop. Prod. 2013, 42, 126–132. [Google Scholar] [CrossRef]
- Oluwaseun, R.A.; Nour, H.A.; Chinonso, I.U. Extraction of phenolic compounds: A review. Curr. Res. Food Sci. 2021, 4, 200–214. [Google Scholar] [CrossRef]
- Jakopic, J.; Colaric, M.; Veberic, R.; Hudina, M.; Solar, A.; Stampar, F. How much do cultivar and preparation time influence on phenolics content in walnut liqueur? Food Chem. 2007, 104, 100–105. [Google Scholar] [CrossRef]
- Cosmulescu, S.; Trandafir, I.; Nour, V.; Ionica, M.; Tutulescu, F. Phenolics content, antioxidant activity and color of green walnut extracts for preparing walnut liquor. Not. Bot. Horti Agrobot. 2014, 42, 551–555. [Google Scholar] [CrossRef] [Green Version]
- Uğurlu, S.; Okumuş, E.; Bakkalbaşı, E. Reduction of Bitterness in Green Walnuts By Conventional And Ultrasound-Assisted Maceration. Ultrason. Sonochem. 2020, 66, 105094. [Google Scholar] [CrossRef] [PubMed]
- Binici, H.; Şat, İ.; Aoudeh, E. Nutritional Composition and Health Benefits of Walnut and its Products. Atatürk Üniversitesi Ziraat Fakültesi Derg. 2021, 52, 224–230. [Google Scholar] [CrossRef]
- Cindrić, I.; Zeiner, M.; Hlebec, D. Mineral Composition of Elements in Walnuts and Walnut Oils. Int. J. Environ. Res. Public Health 2018, 15, 2674. [Google Scholar] [CrossRef] [PubMed]
- Grosso, A.; Asensio, C.; Nepote, V.; Grosso, N. Antioxidant Activity Displayed by Phenolic Compounds Obtained from Walnut Oil Cake used for Walnut Oil Preservation. J. Am. Oil Chem. Soc. 2018, 95, 1409–1419. [Google Scholar] [CrossRef]
- Rabrenović, B.B.; Natić, M.; Dabić Zagorac, D.; Meland, M.; Fotirić Akšić, M. Bioactive Phytochemicals from Walnut (Juglans spp.) Oil Processing By-Products. In Bioactive Phytochemicals from Vegetable Oil and Oilseed Processing By-Products; Ramadan Hassanien, M.F., Ed.; Reference Series in Phytochemistry; Springer: Cham, Switzerland, 2021; pp. 189–197. [Google Scholar] [CrossRef]
- Beiki, T.; Najafpour, G.D.; Hosseini, M. Evaluation of antimicrobial and dyeing properties of walnut (Juglans regia L.) green husk extract for cosmetics. Color. Technol. 2018, 134, 71–81. [Google Scholar] [CrossRef]
- Tsiantos, J.; Vagelas, I.K.; Rumbos, C.I.; Chatzaki, A.; Rouskas, D.; Gravanis, F.T. Evaluation of resistance of cultivated walnut varieties and selections to Xanthomonas arboricola pv. juglandis in Greece. In Proceedings of the COST873 Management Committee Meeting, Athens, Greece, 15 May 2008; Available online: http://www.cost873.ch/uploads/files/GRXajSheetWalnut.pdf (accessed on 26 April 2020).
- Han, M.M.; Shi, S.Y.; Wang, F.; Pang, M.X.; Qu, C.; Qi, J.H. Extraction and Dyeing Propertie of Juglone from Walnut Green Husk. IOP Conf. Ser. Earth Environ. Sci. 2018, 199, 042008. [Google Scholar] [CrossRef]
- Soto-Maldonado, C.; Vergara-Castro, M.; Jara-Quezada, J.; Caballero-Valdés, E.; Müller-Pavez, A.; Zúñiga-Hansen, M.E.; Altamirano, C. Polyphenolic extracts of walnut (Juglans regia) green husk containing juglone inhibit the growth of HL-60 cells and induce apoptosis. Electronic. J. Biotechnol. 2019, 39, 1–7. [Google Scholar] [CrossRef]
- Frutos, D.; Ortega, G. Search for Juglans regia genotypes resistant/tolerant to Xanthomonas arboricola pv. Juglandis in the framework of COST Action 873. J. Plant Pathol. 2012, 94, 37–46. [Google Scholar] [CrossRef]
- Pereira, J.A.; Oliveira, I.; Sousa, A.; Valentão, P.; Andrade, P.B.; Ferreira, I.C.F.R.; Ferreres, F.; Bento, A.; Seabra, R.; Estevinho, L. Walnut (Juglans regia L.) leaves: Phenolic com-pounds, antibacterial activity and antioxidant potential of different cultivars. Food Chem. Toxicol. 2007, 45, 2287–2295. [Google Scholar] [CrossRef] [PubMed]
- Matias, J.; Jakopic, J.; Solar, A.; Rovira, M.; Aletà, N. Walnut varietal patterns of polyphenol contents. In Proceedings of the Cost 873, WG3/WG4 Joint Meeting, Murcia, Spain, 23–25 October 2007; Available online: http://www.cost873.ch/_uploads/_files/m_Matias_2007_10.pdf (accessed on 10 May 2022).
- Matias, J.; Aletà, N.; Rovira, C.; Moragrega, C. Phenoliccomposition in Juglans regia Commercial Cultivars. Relationship with Blight Susceptibility. In Proceedings of the Annual Cost 873 Meeting, Cetara, Italy, 26–29 October 2009; Available online: http://www.cost873.ch/_uploads/_files/JMatias_Walnut-Phenolics_Italy_1.pdf (accessed on 10 May 2022).
- Solar, A.; Jakopic, J.; Veberic, R.; Stampar, F. Phenolic compounds as a potential marker of walnut resistance against Xanthomonas arboricola pv. juglandis. In Proceedings of the Cost 873, WG3/WG4 Joint Meeting, Murcia, Spain, 23–25 October 2007; Available online: https://www.cost873.ch/_up_loads/_files/m_Solar_murcia (accessed on 10 May 2022).
- Dirlewanger, E. Genetic mapping and MAS of resistancegenes in stone fruit and nut trees at INRA. In Proceedings of the Cost 873 STF Meeting, Barcelona, Spain, 2010; Available online: https://www.cost873.ch/_uploads/_files/EDirlewanger_GeneticMappingPeach (accessed on 10 May 2022).
- Solar, A.; Jakopic, J.; Mikulic-Petkovsěk, M.; Veberic, R.; Dreo, T.; Zadravec, P.; Stampar, F. Validation of polyphenol contents as biochemical markers for walnut blight (Xanthomonas arboricola pv. juglandis resistance. In Proceedings of the Cost 873, WG and Management Committee Meeting, Athens, Greece, 20–23 October 2008; Available online: http://www.cost873.ch/_uploads/_files/m_Athens_Abstracts_FinalBook.pdf (accessed on 10 May 2022).
- Yang, H. Resistance evaluation of walnut (Juglans spp.) against Xanthomonas arboricola and the correlation between leaf structure and resistance. For. Pathol. 2021, 51, e12659. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1999, 16, 144–158. [Google Scholar]
- Scalbert, A.; Monties, B.; Janin, G. Tannins in Wood: Comparison of Different Estimation Methods. J. Agric. Food Chem. 1989, 5, 1324–1329. [Google Scholar] [CrossRef]
- Stampar, F.; Solar, A.; Hudina, M.; Veberic, R.; Colaric, M. Traditional walnut liqueur—Cocktail of phenolics. Food Chem. 2006, 95, 627–631. [Google Scholar] [CrossRef]
- Solar, A.; Colarič, M.; Usenik, V.; Stampar, F. Seasonal variations of selected flavonoids, phenolic acids, and quinones in annual shoots of common walnut (Juglans regia L.). Plant Sci. 2006, 170, 453–461. [Google Scholar] [CrossRef]
- Szentiványi, P.; Kállay, T. (Eds.) Dió [Walnut]. In Diófajták Leírása [Description of Walnut Cultivars]; Mezőgazda Kiadó: Budapest, Hungary, 2006; pp. 62–65. [Google Scholar]
- Sagawa, H.D.C.; de Assis, A.B.R.; Zaini, P.A.; Wilmarth, P.A.; Phinney, B.S.; Moreira, L.M.; Dandekar, A.M. Proteome Analysis of Walnut Bacterial Blight Disease. Int. J. Mol. Sci. 2020, 21, 7453. [Google Scholar] [CrossRef]
- Bujdosó, G.; Fodor, A.; Végh, A. BD6 walnut. Hortscience 2020, 8, 1393–1394. [Google Scholar] [CrossRef]
- Dobos, E.; Bialko, T.; Micheli, E.; Kobra, J. Legacy soil data harmonization and database development. In Digital Soil Mapping; Boettinger, J.L., Howell, D.W., Moore, A.C., Hartemink, A.E., Kienast-Brown, S., Eds.; Progress in Soil Science; Springer: Dordrecht, Germany, 2010; Volume 2, pp. 215–223. [Google Scholar]
- Germain, E.; Prunet, J.-P.; Garcin, A. Le Noyer [The walnut]. In Biologie Florale [Flower Biology]; Centr’Imprim: Issoudun, France, 1999; pp. 43–56. [Google Scholar]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventos, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteu reagent. Method Enzymol. 1999, 299, 152–178. [Google Scholar]
- Nour, V.; Trandafir, I.; Cosmulescu, S. HPLC Determination of Phenolic Acids, Flavonoids and Juglone in Walnut Leaves. J. Chromatogr. Sci. 2012, 51, 883–890. [Google Scholar] [CrossRef] [PubMed]
- Huo, J.-H.; Du, X.-W.; Sun, G.-D.; Dong, W.-T.; Wang, W.-M. Identification and characterization of major constituents in Juglans mandshurica using ultra performance liquid chromatography coupled with time-of-flight mass spectrometry (UPLC-ESI-Q-TOF/MS). Chin. J. Nat. Med. 2018, 7, 525–545. [Google Scholar] [CrossRef]
- Phenol-Explorer. Available online: https://phenol-explorer.eu/ (accessed on 10 May 2022).
- MassBank of North America. Available online: https://mona.fiehnlab.ucdavis.edu/ (accessed on 10 May 2022).
- PubChem. Available online: https://pubchem.ncbi.nlm.nih.gov/ (accessed on 10 May 2022).
- Ozaktan, H.; Erdal, M.; Akkopru, A.; Aslan, E. Evaluation of susceptibility of some walnut cultivars to Xanthomonas arboricola pv. juglandis by immature nut test. In Proceedings of the COST873 Management Committee Meeting, Athens, Greece, 20–23 October 2008; Available online: https://www.cost873.ch/uploads/files/GRXapFactSheetWalnut.pdf (accessed on 26 April 2020).
- Solar, A.; Jakopič, J.; Veberic, R.; Stampar, F. Correlations between Xanthomonas arboricola pv. juglandis severity and endogenous juglone and phenolic acids in walnut. J. Plant Pathol. 2012, 94, 229–235. [Google Scholar] [CrossRef]
- Botu, M.; Tudor, M.; Papachatzis, A. Evaluation of some walnut cultivars with different bearing habits in the ecological conditions of Oltenia—Romania. Acta Hortic. 2010, 861, 119–126. [Google Scholar] [CrossRef]
- Bujdosó, G.; Izsépi, F.; Szügyiné Bartha, K.; Varjas, V.; Szentiványi, P. Persian walnut breeding program at Naric Fruticulture Research Institute in Hungary. Acta Hortic. 2020, 1280, 89–94. [Google Scholar] [CrossRef]
- Solar, A.; Stampar, F. Evaluation of some perspective walnut genotypes in Slovenia. Acta Hortic. 2005, 705, 131–136. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Budburst | Blossom Time | |
|---|---|---|
| hydroxybenzoic acids | 0.27 | 0.27 |
| hydroxycinnamic acids | 0.15 | 0.14 |
| flavonoids | 0.37 | 0.31 |
| naphthoquinones | 0.70 | 0.78 |
| total phenolic acids | 0.40 | 0.35 |
| budburst | 0.79 |
| Parameters | Value |
|---|---|
| average yearly temperature | 11.4 °C |
| average yearly temperature during the growing season (March–September) | 16.1 °C |
| average yearly luminous flux | 1015 L/m2/day |
| average yearly precipitation | 434.1 mm |
| annual average of sunshine hours | 2065 |
| Cultivars | Nut Size (Diameter) (mm) | Dried Kernel Weight (g/Nut) | Kernel Color |
|---|---|---|---|
| Alsószentiváni 117 | 33–36 | 4.4–6.2 | medium brown |
| Milotai 10 | 33–35 | 5.2–7.5 | light yellow |
| Tiszacsécsi 83 | 32–34 | 4.4–5.3 | medium brown |
| Köpcös | 32–34 | 4.6–4.8 | medium brown |
| Chandler | 30–32 | 4.2–4.5 | light yellow |
| Milotai intenzív | 32–34 | 4.4–5.3 | light yellow |
| Bonifác | 32–34 | 4.8–6.2 | light brown |
| Milotai kései | 32–34 | 5.3–6.6 | light brown |
| Alsószentiváni kései | 32–35 | 4.4–6.2 | light brown |
| Budburst (Days) 1 | Pistillate Flowers’ Receptivity (Days) 2 | Days after Receptivity before Sampling 3 | |
|---|---|---|---|
| Alsószentiváni 117 | 104 a | 118 a | 63 a |
| Milotai 10 | 107 ab | 118 a | 63 a |
| Tiszacsécsi 83 | 103 a | 118 a | 63 a |
| Köpcös | 108 ab | 118 a | 63 a |
| Chandler | 109 ab | 119 a | 62 a |
| Milotai intenzív | 107 ab | 128 b | 53 b |
| Bonifác | 116 b | 133 c | 48 c |
| Milotai kései | 120 c | 135 c | 45 c |
| Alsószentiváni kései | 120 c | 133 c | 48 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bujdosó, G.; Lengyel-Kónya, É.; Berki, M.; Végh, A.; Fodor, A.; Adányi, N. Effects of Phenolic Compounds on Walnut Bacterial Blight in the Green Husk of Hungarian-Bred Cultivars. Plants 2022, 11, 2996. https://doi.org/10.3390/plants11212996
Bujdosó G, Lengyel-Kónya É, Berki M, Végh A, Fodor A, Adányi N. Effects of Phenolic Compounds on Walnut Bacterial Blight in the Green Husk of Hungarian-Bred Cultivars. Plants. 2022; 11(21):2996. https://doi.org/10.3390/plants11212996
Chicago/Turabian StyleBujdosó, Géza, Éva Lengyel-Kónya, Mária Berki, Anita Végh, Attila Fodor, and Nóra Adányi. 2022. "Effects of Phenolic Compounds on Walnut Bacterial Blight in the Green Husk of Hungarian-Bred Cultivars" Plants 11, no. 21: 2996. https://doi.org/10.3390/plants11212996