
Impact of Foliar Application of Amino Acids on Essential Oil Content, Odor Profile, and Flavonoid Content of Different Mint Varieties in Field Conditions

 , and
, and 
Abstract
:1. Introduction
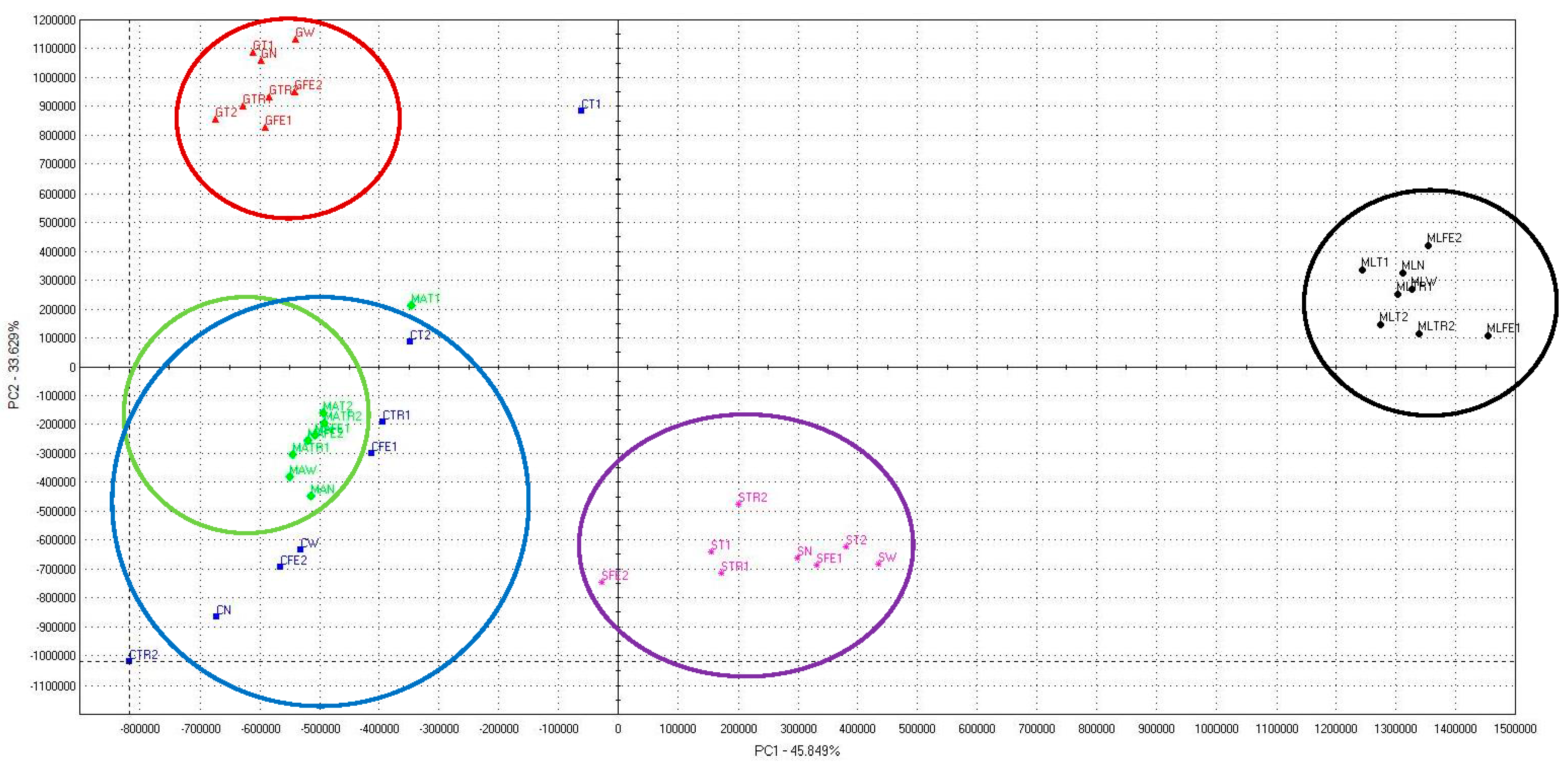
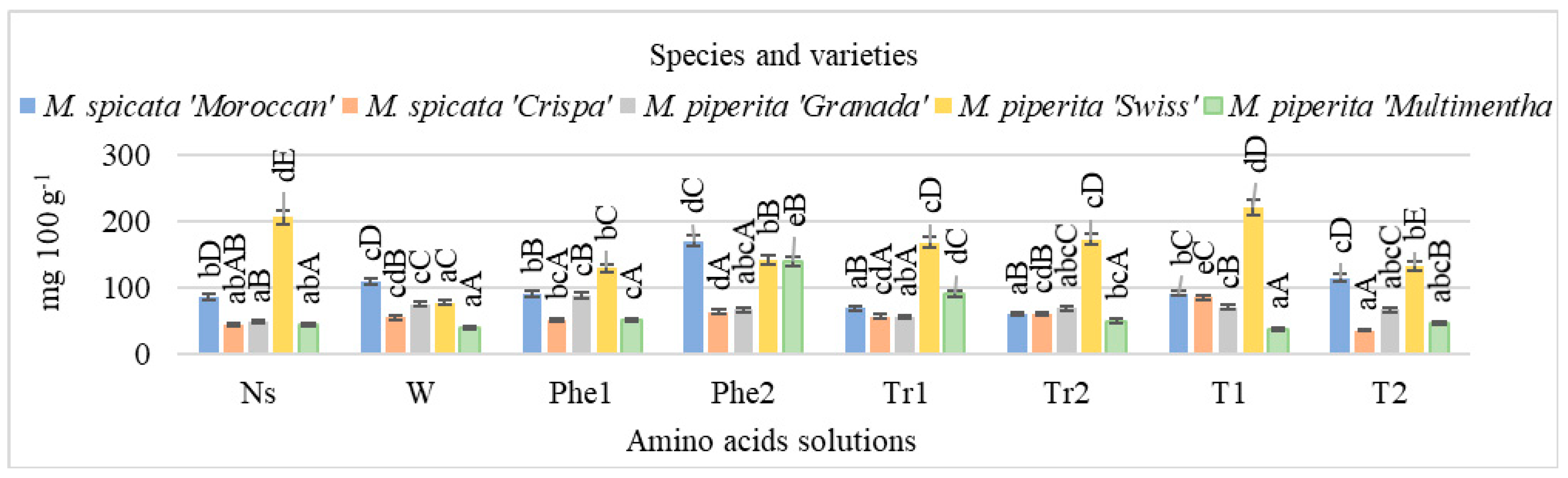
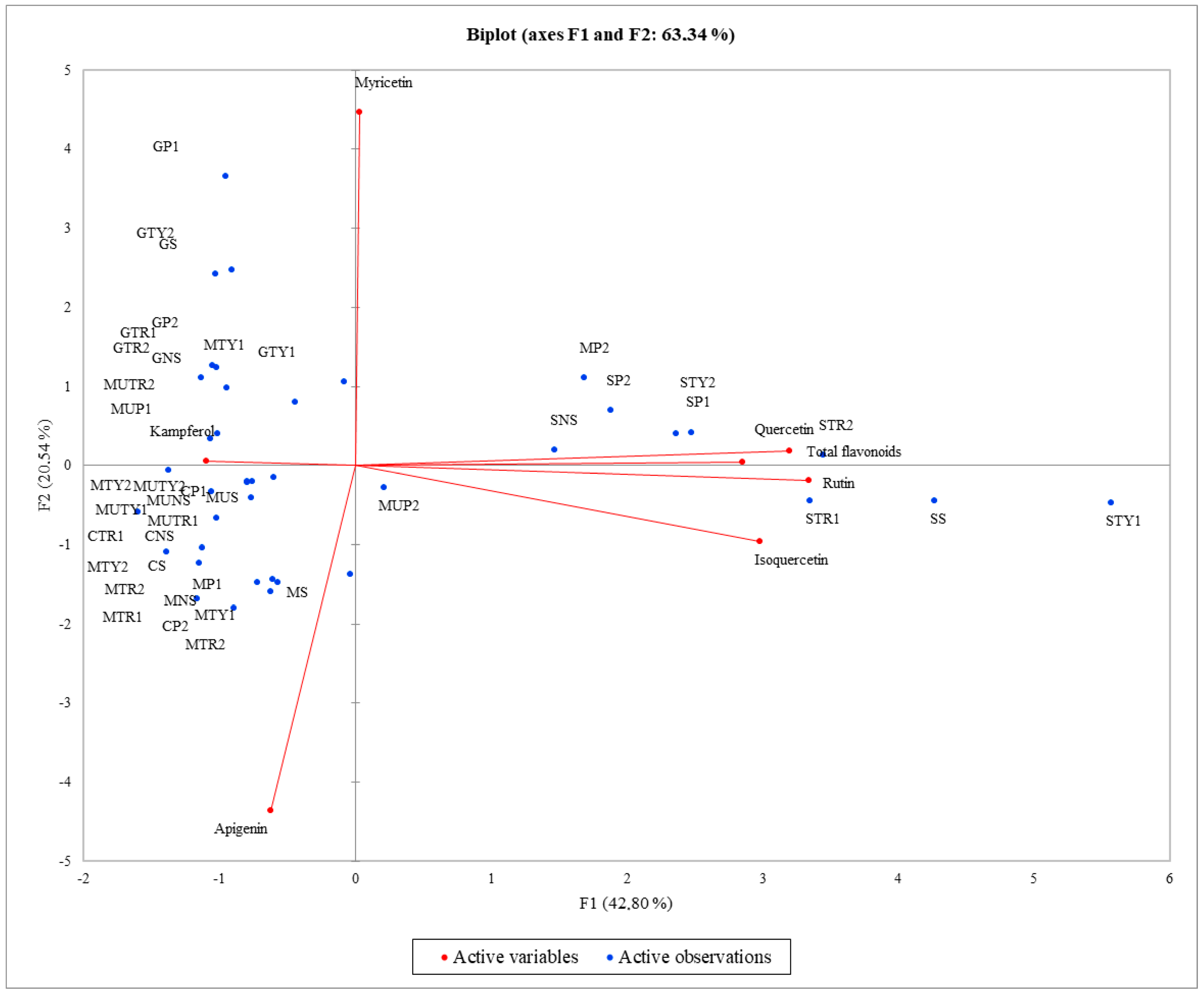
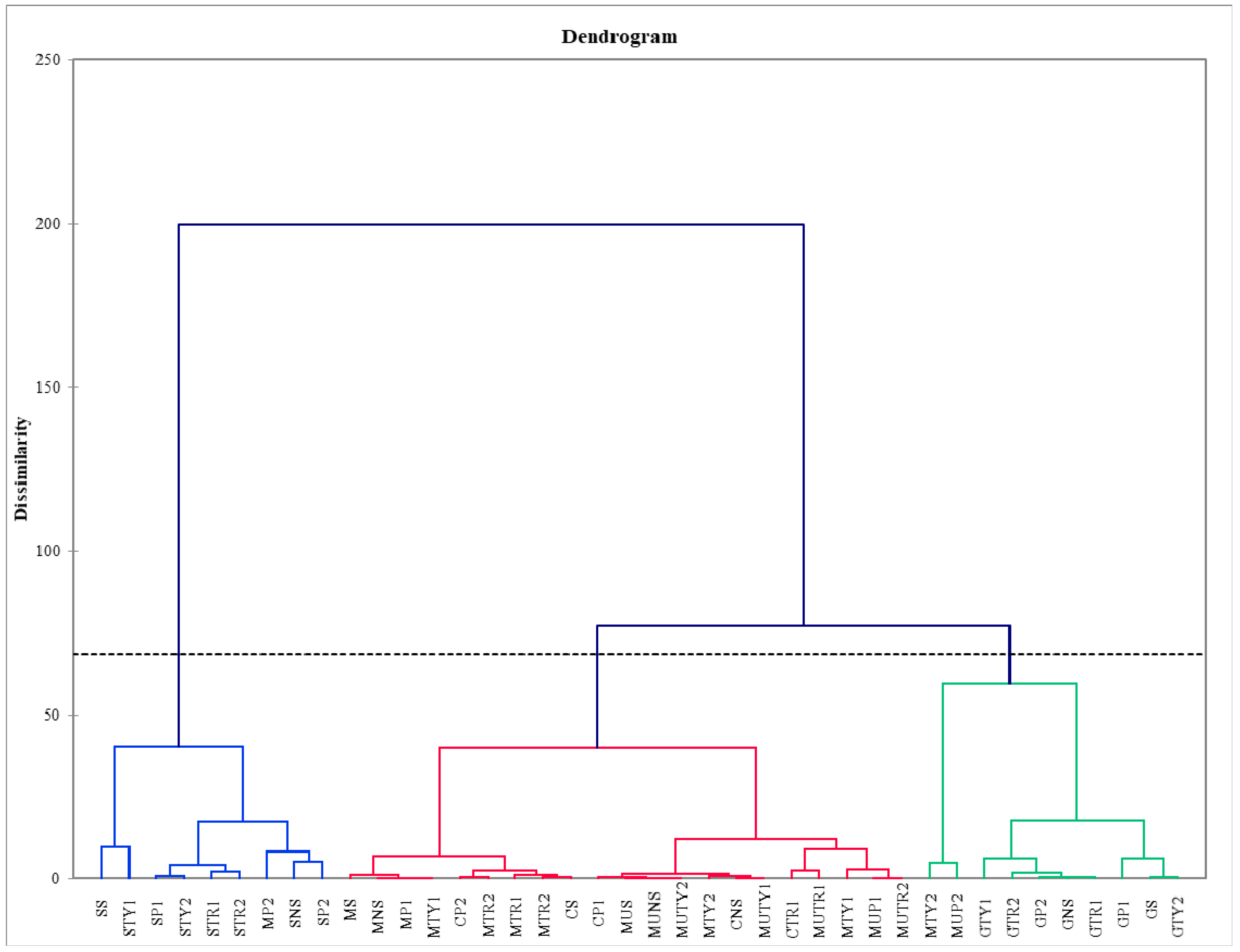
2. Results
3. Discussion
4. Material and Methods
4.1. Experimental Sites and Soil
4.2. Meteorological Conditions
4.3. Chemicals
4.4. Methods of Sample Preparation
4.5. Essential Oil Content
4.6. Flavonoid Content
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Teles, S.; Pereirab, J.A.; Santosa, C.H.B.; Menezesa, R.V.; Malheirob, R.; Lucchesed, A.M.; Silva, F. Effect of geographical origin on the essential oil content and composition of fresh and dried Mentha × villosa Hudson leaves. Ind. Crops Prod. 2013, 46, 1–7. [Google Scholar] [CrossRef]
- Telci, I.; Kacar, O.; Bayram, E.; Arabacı, O.; Demirtaş, İ.; Yılmaz, G.; Sönmez, Ç.; Göksu, E. The effect of ecological conditions on yield and quality traits of selected peppermint (Mentha piperita L.) clones. Ind. Crops Prod. 2011, 34, 1193–1197. [Google Scholar] [CrossRef]
- Lawrence, B.M. Mint: The Genus Mentha; CRC Press: Boca Raton, FL, USA, 2006; pp. 1–56. [Google Scholar]
- Verma, R.S.; Rahman, L.; Verma, R.K.; Chauhan, A.; Yadav, A.K.; Singh, A. Essential Oil Composition of Menthol Mint (Mentha arvensis) and Peppermint (Mentha piperita) Cultivars at Different Stages of Plant Growth from Kumaon Region of Western Himalaya. J. Med. Aromat. Plants 2010, 1, 13–18. [Google Scholar]
- Karousou, R.; Balta, M.; Hanlidou, E.; Kokkini, S. Mints, smells and traditional uses in Thessaloniki (Greece) and other Mediterranean countries. J. Ethnopharmacol. 2007, 109, 248–257. [Google Scholar] [CrossRef]
- Oksman-Caldentey, K.M.; Inze, D. Plant cell factories in the post-genomic era: New ways to produce designer secondary metabolites. Trends Plant Sci. 2004, 9, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Wen, K.S.; Ruan, X.; Zhao, Y.X.; Wei, F.; Wang, Q. Response of Plant Secondary Metabolites to Environmental Factors. Molecules 2018, 23, 762. [Google Scholar] [CrossRef] [Green Version]
- Wink, M. Evolution of Secondary Plant Metabolism. In eLS; John Wiley & Sons, Ltd.: Chichester, UK, 2016; pp. 1–11. ISBN 978-0-470-01590-2. [Google Scholar]
- Pereira, O.R.; Cardoso, S.M. Overview on Mentha and Thymus Polyphenols. Curr. Anal. Chem. 2013, 9, 382–396. [Google Scholar] [CrossRef] [Green Version]
- Salin, O.; Törmäkangas, L.; Leinonen, M.; Saario, E.; Hagström, M.; Ketola, R.A.; Saikku, P.; Vuorela, H.; Vuorela, P.M. Corn Mint (Mentha arvensis) Extract Diminishes Acute Chlamydia pneumoniae Infection In Vitro and In Vivo. J. Agric. Food Chem. 2011, 59, 12836–12842. [Google Scholar] [CrossRef]
- Fatiha, B.; Didier, H.; Naima, G.; Khodir, M.; Martin, K.; Léocadie, K.; Caroline, S.; Mohamed, C.; Pierre, D. Phenolic composition, in vitro antioxidant effects and tyrosinase inhibitory activity of three Algerian Mentha species: M. spicata (L.), M. pulegium (L.) and M. rotundifolia (L.) Huds (Lamiaceae). Ind. Crops Prod. 2015, 74, 722–730. [Google Scholar] [CrossRef]
- Benabdallah, A.; Rahmoune, C.; Boumendjel, M.; Aissi, O.; Messaoud, C. Total phenolic content and antioxidant activity of six wild Mentha species (Lamiaceae) from northeast of Algeria. Asian Pac. J. Trop. Biomed. 2016, 6, 760–766. [Google Scholar] [CrossRef] [Green Version]
- Horvath, P.; Koščova, J. In vitro Antibacterial Activity of Mentha Essential Oils against Staphylococcus aureus. Folia Vet. 2017, 61, 71–77. [Google Scholar] [CrossRef]
- Mahboubi, M. Mentha spicata as natural analgesia for treatment of pain in osteoarthritis patients. Complement. Ther. Clin. Pract. 2017, 26, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Bulgari, R.; Cocetta, G.; Trivellini, A.; Vernieri, P.; Ferrante, A. Biostimulants and crop responses: A review. Biol. Agric. Hortic. 2015, 31, 1–17. [Google Scholar] [CrossRef]
- Mehrabi, S.; Mehrafarin, A.; Badi, H.N. Clarifying the role of methanol and amino acids application on savory (Satureja hortensis L.). Ann. Biol. Res. 2013, 4, 190–195. [Google Scholar]
- Rafiee, H.; Mehrafarin, A.; Qaderi, A.; Jari, S.K.; Badi, H.N. Phytochemical, Agronomical and Morphological Responses of Pot Marigold (Calendula officinalis L.) to Foliar Application of Bio-stimulators (Bioactive Amino Acid Compounds). J. Med. Plants 2013, 12, 48–61. [Google Scholar]
- Hendawy, S.F.; Hussein, M.S.; El-Gohary, A.E.; Ibrahim, M.E. Effect of Foliar Organic Fertilization on the Growth, Yield and Oil Content of Mentha piperita var. citrata. Asian J. Agric. Res. 2015, 9, 237–248. [Google Scholar] [CrossRef] [Green Version]
- Wahba, H.E.; Motawe, H.M.; Ibrahim, A.Y. Growth and chemical composition of Urtica pilulifera L. plant as influenced by foliar application of some amino acids. J. Mater. Environ. Sci. 2015, 6, 499–506. [Google Scholar]
- Aghaye Noroozlo, Y.; Souri, M.K.; Delshad, M. Effects of foliar application of glycine and glutamine amino acids on growth and quality of sweet basil. Adv. Hortic. Sci. 2019, 33, 495–501. [Google Scholar]
- Ayyat, A.M.; Kenawy, A.G.M.; Aboel-Ainin, M.A.; Abdel-Mola, M.A.M. Improving Growth, Productivity and Oil Yield of Nigella sativa L. Plants by Foliar Spraying with some Stimulants. J. Plant Prod. 2021, 12, 339–344. [Google Scholar] [CrossRef]
- Shafie, F.; Bayat, H.; Aminifard, M.H.; Saeid Daghighi, S. Biostimulant Effects of Seaweed Extract and Amino Acids on Growth, Antioxidants, and Nutrient Content of Yarrow (Achillea millefolium L.) in the Field and Greenhouse Conditions. Commun. Soil Sci. Plant Anal. 2021, 52, 964–975. [Google Scholar] [CrossRef]
- Wafaa, H.A.A.A.; Rania, M.R.K.; El-Shafay, R.M.M. Effect of spraying with extracts of plants and amino acids on growth and productivity on Coriandrum sativum plants under Shalateen condition. Plant Arch. 2021, 21, 300–307. [Google Scholar]
- Elsayed, A.A.A.; El-Gohary, A.E.; Khalid, K.A.; Ahmed, A.M.A. Changes in Bitter Fennel Essential Oils Exposed to Foliar Spray with L-Phenylalanine. Egypt. J. Bot. 2022, 62, 241–253. [Google Scholar] [CrossRef]
- Pratelli, R.; Pilot, G. Regulation of amino acid metabolic enzymes and transporters in plants. J. Exp. Bot. 2014, 65, 5535–5556. [Google Scholar] [CrossRef] [PubMed]
- Maeda, H.; Dudareva, N. The Shikimate Pathway and Aromatic Amino Acid Biosynthesis in Plants. Annu. Rev. Plant Biol. 2012, 63, 73–105. [Google Scholar] [CrossRef]
- Tegeder, M.; Masclaux-Daubresse, C. Source and sink mechanisms of nitrogen transport and use. New Phytol. 2018, 217, 35–53. [Google Scholar] [CrossRef] [Green Version]
- Ma, H.L.; Cao, X.L.; Shi, S.D.; Li, S.; Gao, J.; Ma, Y.; Zhao, Q.; Chen, Q. Genome-wide survey and expression analysis of the amino acid transporter superfamily in potato (Solanum tuberosum L.). Plant Physiol. Biochem. 2016, 107, 164–177. [Google Scholar] [CrossRef]
- Barros, J.; Serrani-Yarce, J.C.; Chen, F.; Baxter, D.; Venables, B.J.; Dixon, R.A. Role of bifunctional ammonia-lyase in grass cell wall biosynthesis. Nat. Plants 2016, 2, 16050. [Google Scholar] [CrossRef]
- Barros, J.; Dixon, R.A. Plant Phenylalanine/Tyrosine Ammonia-lyases. Trends Plant Sci. 2020, 25, 66–79. [Google Scholar] [CrossRef]
- Croteau, R.; Kutchan, T.M.; Lewis, N.G. Natural Products (Secondary Metabolites). Biochem. Mol. Biol. Plants 2000, 24, 1250–1319. [Google Scholar]
- Musbah, H.M.; Ibrahim, K.M. Effects of feeding Tyrosine or Phenylalanine on the accumulation of polyphenols in Coleus blumei in vivo and in vitro. J. Biotechnol. Res. Cent. 2019, 13, 35–43. [Google Scholar] [CrossRef]
- Baharlou, M.J.; Pirbalouti, A.G.; Malekpoor, F. Effect of different concentrations of L-phenylalanine on chemical compositions and yield of essential oil of lemon balm (Melissa officinalis). J. Herb. Drugs 2019, 10, 175–183. [Google Scholar]
- Reham, M.S.; Khattab, M.E.; Ahmed, S.S.; Kandil, M.A.M. Influence of foliar spray with phenylalanine and nickel on growth, yield quality and chemical composition of genoveser basil plant. Afr. J. Agric. Res. 2016, 11, 1398–1410. [Google Scholar] [CrossRef] [Green Version]
- Noviyanti, R.; Sari, R.L.K.; Kristanti, A.N.; Yachya, A.; Manuhara, Y.S.W. Biomass and flavonoid production of gynura procumbens adventitious roots induced by sucrose, Phenylalanine and Tyrosine. Biosci. Res. 2017, 14, 934–941. [Google Scholar]
- Sachet, T.F.K.; Al-Mohammad, M.H.S.; Abbass, J.A. Effect of Biofertilizer and Spraying Phenylalanine on Mineral Content and Antioxidant Compounds of Dill. Indian J. Forensic Med. Toxicol. 2021, 15, 5540–5545. [Google Scholar]
- Poorghadir, M.; Torkashvand, A.M.; Mirjalili, S.A.; Moradi, P. Interactions of amino acids (proline and phenylalanine) and biostimulants (salicylic acid and chitosan) on the growth and essential oil components of savory (Satureja hortensis L.). Biocatal. Agric. Biotechnol. 2020, 30, 101815. [Google Scholar] [CrossRef]
- Samani, M.R.; Pirbalouti, A.G.; Moattar, F.; Golparvar, A.R. L-Phenylalanine and bio-fertilizers interaction effects on growth, yield and chemical compositions and content of essential oil from the sage (Salvia officinalis L.) leaves. Ind. Crops Prod. 2019, 137, 1–8. [Google Scholar] [CrossRef]
- Aghaei, K.; Pirbalouti, A.G.; Mousavi, G.A.; Badi, H.; Mehnatkesh, A. Effects of foliar spraying of l-phenylalanine and application of bio-fertilizers on growth, yield, and essential oil of hyssop [Hyssopus officinalis L. subsp. Angustifolius (Bieb.)]. Biocatal. Agric. Biotechnol. 2019, 21, 101318. [Google Scholar] [CrossRef]
- Tanaka, Y.; Sasaki, N.; Ohmiya, A. Biosynthesis of plant pigments: Anthocyanins, betalains and carotenoids. Plant J. 2008, 54, 733–749. [Google Scholar] [CrossRef]
- Pagare, S.; Bhatia, M.; Tripathi, N.; Pagare, S.; Bansal, Y.K. Secondary Metabolites of Plants and their Role: Overview. Curr. Trends Biotechnol. Pharm. 2015, 9, 293–304. [Google Scholar]
- EL-Zefzafy, M.M.; Shahhat, I.M.A.; Yousef, R.S.; Elsharkawy, E.R. Influence of foliar application with amino acids and citric acid on physiological and phytochemical responses of Artemisia abrotanum produced by in vitro culture. Biotechnol. Commun. Res. Commun. 2016, 9, 702–711. [Google Scholar] [CrossRef]
- Jacob, A.; Thomas, J. Flavonoids from cell suspension culture of Ocimum tenuiflorum and its enhancement using response surface methodology. Drug Invent. Today 2019, 11, 2188–2193. [Google Scholar]
- Kawka, B.; Kwiecień, I.; Ekiert, H. Production of Specific Flavonoids and Verbascoside in Shoot Cultures of Scutellaria baicalensis. In Plant Cell and Tissue Differentiation and Secondary Metabolites; Springer: Berlin/Heidelberg, Germany, 2019; pp. 1–24. [Google Scholar]
- Roy, D.; Mallick, B.; Samanta, D. Augmentation of antioxidative potential of in vitro propagated Mentha piperita L. Indian J. Exp. Biol. 2020, 58, 131–137. [Google Scholar]
- Talaat, I.M.; Khattab, H.I.; Ahmeda, A.M. Changes in growth, hormones levels and essential oil content of Ammi visnaga L. plants treated with some bioregulators. Saudi J. Biol. Sci. 2014, 21, 355–365. [Google Scholar] [CrossRef] [PubMed]
- Schwab, W.; Davidovich-Rikanati, R.; Lewinsohn, E. Biosynthesis of plant-derived flavor compounds. Plant J. 2008, 54, 712–732. [Google Scholar] [CrossRef]
- Gleadow, R.M.; Møller, B.L. Cyanogenic Glycosides: Synthesis, Physiology, and Phenotypic Plasticity. Annu. Rev. Plant Biol. 2014, 65, 155–185. [Google Scholar] [CrossRef]
- Jiao, Y.; Chen, Y.; Ma, C.; Qin, J.; Nguyen, T.H.N.; Liu, D. Phenylalanine as a nitrogen source induces root growth and nitrogen-use efficiency in Populus × canescens. Tree Physiol. 2017, 38, 66–82. [Google Scholar] [CrossRef] [Green Version]
- Khalid, A.K. Effect of NP and foliar spray on growth and chemical compositions of some medicinal Apiaceae plants grow in arid regions in Egypt. J. Soil Sci. Plant Nutr. 2012, 12, 617–632. [Google Scholar] [CrossRef]
- Dewick, P.M. The biosynthesis of C5–C25 terpenoid compounds. Nat. Prod. Rep. 1997, 14, 111–144. [Google Scholar] [CrossRef]
- Baranauskiene, R.; Venskutonis, P.R.; Viskelis, P.; Dambrauskiene, E. Influence of nitrogen fertilizers on the yield and composition of thyme (Thymus vulgaris). J. Agric. Food Chem. 2003, 51, 7751–7758. [Google Scholar] [CrossRef]
- Aroiee, H. Effect of Nitrogen Nutrition on Growth and Essential Oil Production of Peppermint (Mentha piperita). KMITL Sci. J. 2006, 6, 670–673. [Google Scholar]
- Ashraf, M.; Ali, Q.; Iqbal, Z. Effect of nitrogen application rate on the content and composition of oil, essential oil and minerals in black cumin (Nigella sativa L.) seeds. J. Sci. Food Agric. 2006, 86, 871–876. [Google Scholar] [CrossRef]
- Akbarinia, A.; Jahanfar, D.; Farzad, M.B. Effect of nitrogen fertilizer and plant density on seed yield, essential oil and oil content of Coriandrum sativum L. Iran. J. Med. Aromat. Plant 2007, 22, 410–419. [Google Scholar]
- Khalid, A.K. Nitrogen application and different plant densities effectiveness on the productivity of parsley crop. Asian J. Crop Sci. 2018, 10, 141–150. [Google Scholar] [CrossRef]
- Cheynier, V.; Comte, G.; Davies, K.M.; Lattanzio, V.; Martense, S. Plant phenolics: Recent advances on their biosynthesis, genetics, and ecophysiology. Plant Physiol. Biochem. 2013, 72, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Harborne, J.B. Classes and Functions of Secondary Products from Plants; Imperial College Press: London, UK, 1999; pp. 1–26. [Google Scholar]
- Koes, R.; Verweij, W.; Qwattrocchio, F. Flavonoids: A colorful model for the regulation and evolution of biochemical pathways. Trends Plant Sci. 2005, 10, 236–242. [Google Scholar] [CrossRef]
- Vogt, T. Phenylpropanoid biosynthesis. Mol. Plant. 2010, 3, 2–20. [Google Scholar] [CrossRef] [Green Version]
- Feduraev, P.; Skrypnik, L.; Riabova, A.; Pungin, A.; Tokupova, E.; Maslennikov, P.; Chupakhina, G. Phenylalanine and Tyrosine as Exogenous Precursors of Wheat (Triticum aestivum L.) Secondary Metabolism through PAL-Associated Pathways. Plants 2020, 9, 476. [Google Scholar] [CrossRef] [Green Version]
- Demirci, T.; Akcay, U.C.; Baydar, N.G. Efects of 24-epibrassinolide and l-phenylalanine on growth and cafeic acid derivative production in hairy root culture of Echinacea purpurea L. Moench. Acta Physiol. Plant. 2020, 42, 66. [Google Scholar] [CrossRef]
- Radwanski, E.R.; Last, R.L. Tryptophan Biosynthesis and Metabolism: Biochemical and Molecular Genetics. Plant Cell 1995, 7, 921–934. [Google Scholar]
- Bielach, A.; Hrtyan, M.; Tognetti, V.B. Plants under Stress: Involvement of Auxin and Cytokinin. Int. J. Mol. Sci. 2017, 18, 1427. [Google Scholar] [CrossRef] [Green Version]
- Korver, R.A.; Koevoets, I.T.; Testerink, C. Out of Shape During Stress: A Key Role for Auxin. Trends Plant Sci. 2018, 23, 9. [Google Scholar] [CrossRef] [PubMed]
- Bromke, M.A. Amino acid biosynthesis pathways in diatoms. Metabolism 2013, 3, 294–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verpoorte, R.; Van der Heijden, R.; ten Hoopen, H.J.G.; Memelink, J. Metabolic engineering of plant secondary metabolite pathways for the production of fine chemicals. Biotechnol. Lett. 1999, 21, 467–479. [Google Scholar] [CrossRef]
- Klimek-Szczykutowicz, M.; Dziurka, M.; Blaževic, I.; Dulovic, A.; Miazga-Karska, M.; Klimek, K.; Ekiert, H.; Szopa, A. Precursor-Boosted Production of Metabolites in Nasturtium officinale Microshoots Grown in Plantform Bioreactors, and Antioxidant and Antimicrobial Activities of Biomass Extracts. Molecules 2021, 26, 4660. [Google Scholar] [CrossRef]
- Ouyang, J.; Wang, X.D.; Zhao, B.; Wang, Y.C. Enhanced production of phenylethanoid glycosides by precursor feeding to cell culture of Cistanche deserticola. Process Biochem. 2005, 40, 3480–3484. [Google Scholar] [CrossRef]
- Ganeshpurkar, A.; Saluja, A.K. The Pharmacological Potential of Rutin. Saudi Pharm. J. 2017, 25, 149–164. [Google Scholar] [CrossRef]
- Wei, Y.; Xiaonan, X.; Yang, L.; Yingzi, W.; Ming, L.; Yong, W.; Xinhua, D.; Zhaohui, C. Rutin-mediated priming of plant resistance to three bacterial pathogens initiating the early SA signal pathway. PLoS ONE 2016, 11, e0146910. [Google Scholar] [CrossRef]
- El-Ashry, A.A.E.L.; Magdy Gabr, A.H.; Arafa, N.M.; El-Bahr, M.K. Rutin accumulation in gardenia callicultures as a response to phenylalanine and salicylic acid. Bull. Natl. Res. Cent. 2019, 43, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Emura, K.; Yokomoto, A.; Toyoshi, T.; Moriwak, M.N. Effect of enzymatically modified isoquercitrin in spontaneously hypertensive rats. J. Nutr. Sci. Vitam. 2007, 53, 68–74. [Google Scholar] [CrossRef] [Green Version]
- Panda, S.; Kar, A. Antidiabetic and antioxidative effects of Annona squamosa leaves are possibly mediated through quercetin-3-O-glucoside. Biofactors 2007, 31, 201–210. [Google Scholar] [CrossRef]
- Rogerio, A.P.; Kanashiro, A.; Fontanari, C.; da Silva, E.V.G.; Lucisano-Valim, Y.M.; Soares, E.G.; Faccioli, L.H. Anti-inflammatory activity of quercetin and isoquercitrin in experimental murine allergic asthma. Inflamm. Res. 2007, 56, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Valentova, K.; Vrba, J.; Bancirova, M.; Ulrichova, J.; Kren, V. Isoquercitrin: Pharmacology, toxicology, and metabolism. Food Chem. Toxicol. 2014, 68, 267–282. [Google Scholar] [CrossRef] [PubMed]
- Sheng, H.; Sun, X.; Yan, Y.; Yuan, Q.; Wang, J.; Shen, X. Metabolic Engineering of Microorganisms for the Production of Flavonoids. Front. Bioeng. Biotechnol. 2020, 8, 589069. [Google Scholar] [CrossRef] [PubMed]
- Marin, L.; Gutierrez-del-Rio, I.; Entrialgo-Cadierno, R.; Villar, C.J.; Lombo, F. De novo biosynthesis of myricetin, kaempferol and quercetin in Streptomyces albus and Streptomyces coelicolor. PLoS ONE 2018, 13, e0207278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Portu, J.; Lopez, R.; Santamaria, P.; Garde-Cerdan, T. Elicitation with methyl jasmonate supported by precursor feeding with phenylalanine: Effect on Garnacha grape phenolic content. Food Chem. 2017, 237, 416–422. [Google Scholar] [CrossRef]
- Singh, P.; Arif, Y.; Bajguz, A.; Hayat, S. The role of quercetin in plants. Plant Physiol. Biochem. 2021, 166, 10–19. [Google Scholar] [CrossRef]
- Lewis, D.R.; Ramirez, M.V.; Miller, N.D.; Vallabhaneni, P.; Ray, W.K.; Helm, R.F.; Winkel, B.S.J.; Muday, G.K. Auxin and Ethylene Induce Flavonol Accumulation through Distinct Transcriptional Networks. Plant Physiol. 2011, 156, 144–164. [Google Scholar] [CrossRef] [Green Version]
- Meena, M.C.; Meena, R.K.; Patni, V. Effect of elicitor on quersetin production in cell cultures of Citrullus colocynthis (Linn.) Schrad. Pharma Innov. 2014, 3, 2. [Google Scholar]
- Liu, W.; Feng, Y.; Yu, S.; Fan, Z.; Li, X.; Li, J.; Yin, H. The Flavonoid Biosynthesis Network in Plants. Int. J. Mol. Sci. 2021, 22, 12824. [Google Scholar] [CrossRef]
- Duan, L.; Ding, W.; Liu, X.; Cheng, X.; Cai, J.; Hua, E.; Jiang, H. Biosynthesis and engineering of kaempferol in Saccharomyces cerevisiae. Microb. Cell Factories 2017, 16, 165. [Google Scholar] [CrossRef] [Green Version]
- Manuhara, Y.S.W.; Noviyanti, R.; Hardjo, P.H.; Tanjung, M. Enhancement of flavonoid production of Gynura procumbens (Lour.) Merr adventitious roots in baloon-type bubble bioreactor influenced by phenylalanine and tyrosine. Ecol. Environ. Conserv. J. 2019, 25, S61–S66. [Google Scholar]
- Kim, M.S.; Lee, W.K.; Kim, H.Y.; Kim, C.; Ryu, Y.W. Effect of environmental factors on flavonol glycoside production and phenylalanine ammonia-lyase activity in cell suspension cultures of Ginkgo biloba. J. Microbiol. Biotechnol. 1998, 8, 237–244. [Google Scholar]
- Nurisa, A.; Kristanti, A.N.; Manuhara, Y.S.W. Effect of Sucrose, Erythrose-4-Phosphate and Phenylalanine on Biomassa and Flavonoid Content of Callus Culture from Leaves of Gynura procumbens Merr. AIP Conf. Proc. 2017, 1868, 090013. [Google Scholar] [CrossRef] [Green Version]
- Hack, H.; Bleiholder, H.; Buhr, L.; Meier, U.; Schnock-Fricke, U.; Weber, E.; Witzenberger, A. Einheitliche Codierung der Phanologischen Entwicklungsstadien Mono und Dikotyler Pflanzen–Entweirterte BBCH-Skala. Allg. Nachr. Deut Pflanz. 1992, 44, 265–270. [Google Scholar]
- Bac, S.; Koźmiński, C.; Rojek, M. Agrometeorologia; PWN: Warszawa, Poland, 1993; pp. 32–33. [Google Scholar]
- Skowera, B.; Puła, J. Skrajne warunki pluwio termiczne w okresie wiosennym na obszarze Polski w latach 1971–2000. Acta Agroph 2004, 3, 171–177. [Google Scholar]
- Rubinskienė, M.; Viškelis, P.; Dambrauskienė, E.; Viškelis, J.; Karklelienė, R. Effect of drying methods on the chemical composition and colour of peppermint (Mentha × piperita L.) leaves. Zemdirb.-Agric. 2015, 102, 223–228. [Google Scholar] [CrossRef] [Green Version]
- Hallmann, E.; Sabała, P. Organic and Conventional Herbs Quality Reflected by Their Antioxidant Compounds Concentration. Appl. Sci. 2020, 10, 3468. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Species/Varieties | ||||
|---|---|---|---|---|---|
| M. spicata ‘Moroccan’ | M. spicata ‘Crispa’ | M. piperita ‘Granada’ | M. piperita ‘Swiss’ | M. piperita ‘Multimentha’ | |
| Unsprayed | 2.79 ± 0.05 abBC | 1.59 ± 0.01cA | 1.41 ± 0.30 aA | 3.24 ± 0.71 bC | 2.12 ± 0.11 aAB |
| Water | 2.26 ± 0.15 aC | 1.58 ± 0.01 cA | 1.71 ± 0.03 abA | 2.05 ± 0.03 aB | 1.62 ± 0.03 aA |
| Phenylalanine 100 mg L−1 | 2.83 ± 0.03 abB | 1.51 ± 0.05 cA | 1.94 ± 0.15 bAB | 2.85 ± 0.51abB | 1.92 ± 0.67 aAB |
| Phenylalanine 200 mg L−1 | 3.09 ± 0.02 bD | 1.20 ± 0.02 abA | 1.50 ± 0.15 abAB | 2.20 ± 0.24 abC | 1.54 ± 0.03 aB |
| Tryptopan 100 mg L−1 | 2.75 ± 0.79 abC | 1.25 ± 0.07 abA | 1.44 ± 0.07 aAB | 2.40 ± 0.64 abABC | 2.54 ± 0.36 aBC |
| Tryptopan 200 mg L−1 | 3.09 ± 0.23 bC | 1.23 ± 0.03 abA | 1.68 ± 0.35 abAB | 2.34 ± 0.13 abB | 2.09 ± 0.44 aB |
| Tyrosine 100 mg L−1 | 2.71 ± 0.01abC | 1.33 ± 0.10 bA | 1.36 ± 0.05 aA | 2.55 ± 0.20 abBC | 2.06 ± 0.45 aB |
| Tyrosine 200 mg L−1 | 3.04 ± 0.16 abC | 1.19 ± 0.01 aA | 1.37 ± 0.03 aA | 2.65 ± 0.20 abC | 2.11 ± 0.30 aB |
| Treatment | Species/Varieties | ||||
|---|---|---|---|---|---|
| M. spicata ‘Moroccan’ | M. spicata ‘Crispa’ | M. piperita ‘Granada’ | M. piperita ‘Swiss’ | M. piperita ‘Multimentha’ | |
| Unsprayed | 7.01 ± 0.52 aA | 8.73 ± 1.73 bcAB | 10.50 ± 0.75 cdABC | 12.30 ± 3.11 aBC | 13.04 ± 1.06 cC |
| Water | 21.33 ± 3.13 bB | 8.40 ± 0.52 bcA | 9.08 ± 1.03 bcA | 81.15 ± 7.49 dC | 15.95 ± 1.12 cAB |
| Phenylalanine 100 mg L−1 | 7.79 ± 0.57 aA | 10.32 ± 0.94 cA | 5.77 ± 0.90 abA | 35.86 ± 11.62 bB | 12.28 ± 0.90 cA |
| Phenylalanine 200 mg L−1 | 8.46 ± 0.57 aAB | 8.08 ± 0.59 bcAB | 5.45 ± 0.51 aA | 34.27 ± 3.17 bC | 11.80 ± 1.47 cB |
| Tryptopan 100 mg L−1 | 8.50 ± 0.71 aBC | 7.46 ± 0.69 bAB | 12.20 ± 0.75 cdeC | 54.28 ± 3.74cD | 4.01 ± 0.32 aA |
| Tryptopan 200 mg L−1 | 6.25 ± 0.47 aA | 8.72 ± 0.88bcAB | 11.67 ± 0.77 cdB | 41.09 ± 2.84 bcC | 12.14 ± 3.61 cB |
| Tyrosine 100 mg L−1 | 7.79 ± 0.50 aAB | 4.89 ± 0.30 aA | 15.32 ± 2.99 eB | 82.80 ± 6.53dC | 7.72 ± 0.65 abAB |
| Tyrosine 200 mg L−1 | 6.91 ± 0.51 aA | 4.25 ± 0.51 aA | 13.41 ± 0.95 deB | 47.57 ± 3.28 bcC | 13.27 ± 0.89 cB |
| Treatment | Species/Varieties | ||||
|---|---|---|---|---|---|
| M. spicata ‘Moroccan’ | M. spicata ‘Crispa’ | M. piperita ‘Granada’ | M. piperita ‘Swiss’ | M. piperita ‘Multimentha’ | |
| Unsprayed | 3.03 ± 0.12 aA | 3.48 ± 0.11 cdB | 2.84 ± 0.16 abA | 3.62 ± 0.13 aB | 3.57 ± 0.09 cB |
| Water | 2.86 ± 0.11 aA | 3.41 ± 0.09 cB | 2.96 ± 0.12 bA | 7.30 ± 0.07 cD | 3.94 ± 0.17 dC |
| Phenylalanine 100 mg L−1 | 3.05 ± 0.12 aAB | 3.72 ± 0.09 dB | 2.43 ± 0.16 aA | 5.34 ± 0.57 bC | 2.48 ± 0.13 aA |
| Phenylalanine 200 mg L−1 | 3.13 ± 0.10 aA | 4.02 ± 0.10 eC | 2.88 ± 0.11 bA | 3.03 ± 0.13 aA | 3.64 ± 0.17 cdB |
| Tryptopan 100 mg L−1 | 3.16 ± 0.12 aB | 2.45 ± 0.11 aA | 3.02 ± 0.12 bB | 5.41 ± 0.29 bC | 3.18 ± 0.16 bB |
| Tryptopan 200 mg L−1 | 2.95 ± 0.12 aB | 4.60 ± 0.16 fC | 3.03 ± 0.09 bB | 5.14 ± 0.29 bD | 2.33 ± 0.13 aA |
| Tyrosine 100 mg L−1 | 3.08 ± 0.13 aB | 2.30 ± 0.13 aA | 5.09 ± 0.30 cC | 6.78 ± 0.25 cD | 3.51 ± 0.11 bcB |
| Tyrosine 200 mg L−1 | 2.99 ± 0.12 aA | 2.94 ± 0.09 bA | 2.95 ± 0.12 bA | 4.88 ± 0.10 bC | 3.61 ± 0.11 cdB |
| Treatment | Species/Varieties | ||||
|---|---|---|---|---|---|
| M. spicata ‘Moroccan’ | M. spicata ‘Crispa’ | M. piperita ‘Granada’ | M. piperita ‘Swiss’ | M. piperita ‘Multimentha’ | |
| Unsprayed | 5.85 ± 0.58 aA | 6.38 ± 0.57 aA | 13.31 ± 0.77 aB | 5.81 ± 0.59 aA | 5.69 ± 0.51 aA |
| Water | 5.89 ± 0.60 aA | 6.06 ± 0.58 aA | 25.16 ± 0.86 cC | 10.40 ± 0.70 cB | 5.77 ± 0.58 aA |
| Phenylalanine 100 mg L−1 | 5.95 ± 0.58 aA | 5.88 ± 0.57 aA | 39.11 ± 1.43 dC | 9.07 ± 0.67 bcB | 5.91 ± 0.59 aA |
| Phenylalanine 200 mg L−1 | 17.52 ± 2.22 bB | 6.25 ± 0.59 aA | 15.46 ± 0.83 abB | 8.28 ± 0.65 bA | 7.12 ± 0.61 aA |
| Tryptopan 100 mg L−1 | 6.17 ± 0.57 aA | 6.13 ± 0.61 aA | 16.14 ± 0.67 abC | 9.46 ± 0.67 bcB | 6.11 ± 0.59 aA |
| Tryptopan 200 mg L−1 | 5.91 ± 0.55 aA | 6.19 ± 0.59 aA | 15.33 ± 0.83 abC | 8.11 ± 0.55 bB | 5.53 ± 0.57 aA |
| Tyrosine 100 mg L−1 | 6.07 ± 0.56 aA | 8.92 ± 0.66 bB | 16.96 ± 0.91 bD | 12.59 ± 0.60 dC | 6.18 ± 0.59 aA |
| Tyrosine 200 mg L−1 | 6.04 ± 0.57 aA | 5.82 ± 0.60 aA | 25.83 ± 1.83 cC | 9.20 ± 0.58 bcB | 6.13 ± 0.63aA |
| Treatment | Species/Varieties | ||||
|---|---|---|---|---|---|
| M. spicata ‘Moroccan’ | M. spicata ‘Crispa’ | M. piperita ‘Granada’ | M. piperita ‘Swiss’ | M. piperita ‘Multimentha’ | |
| Unsprayed | 29.58 ± 0.28 cB | 4.15 ± 0.09 aA | 2.76 ± 0.10 aA | 45.53 ± 2.31 aC | 2.98 ± 0.09 aA |
| Water | 38.44 ± 1.51 dC | 3.59 ± 0.09 aA | 16.25 ± 0.24 dB | 89.32 ± 1.97 eD | 3.10 ± 0.13 aA |
| Phenylalanine 100 mg L−1 | 32.68 ± 0.86 cdD | 9.11 ± 2.65 bA | 17.52 ± 0.43 eC | 69.29 ± 0.43 cE | 14.24 ± 0.16 bB |
| Phenylalanine 200 mg L−1 | 115.91 ± 5.84 eE | 3.60 ± 0.10 aA | 17.63 ± 0.14 eB | 87.67 ± 0.56 eD | 55.50 ± 0.99 eC |
| Tryptopan 100 mg L−1 | 6.03 ± 0.13 aB | 3.71 ± 0.10 aA | 2.85 ± 0.13 aA | 76.84 ± 0.45 dD | 31.98 ± 1.67 dC |
| Tryptopan 200 mg L−1 | 13.32 ± 0.20 bB | 3.47 ± 0.10 aA | 3.44 ± 0.13 bcA | 104.83 ± 1.52 gD | 18.21 ± 0.34 cC |
| Tyrosine 100 mg L−1 | 33.55 ± 0.62 cdB | 52.35 ± 0.32 cC | 2.92 ± 0.11 abA | 97.37 ± 4.35 fD | 3.04 ± 0.13 aA |
| Tyrosine 200 mg L−1 | 14.03 ± 0.14 bC | 5.83 ± 0.41 aB | 3.98 ± 0.09 cAB | 56.09 ± 1.65 bD | 3.81 ± 0.17 aA |
| Treatment | Species/Varieties | ||||
|---|---|---|---|---|---|
| M. spicata ‘Moroccan’ | M. spicata ‘Crispa’ | M. piperita ‘Granada’ | M. piperita ‘Swiss’ | M. piperita ‘Multimentha’ | |
| Unsprayed | 26.58 ± 1.42 bcD | 16.42 ± 1.36 bcC | 6.35 ± 0.80 bA | 5.81 ± 1.00 aA | 10.03 ± 0.92 cdB |
| Water | 25.53 ± 1.40 bcE | 20.62 ± 1.57 cdD | 3.59 ± 0.72 aA | 12.24 ± 1.01 bC | 8.62 ± 0.87 cB |
| Phenylalanine 100 mg L−1 | 26.26 ± 1.34 bcC | 9.96 ± 2.19 abB | 6.70 ± 0.79 bA | 4.09 ± 0.72 aA | 5.63 ± 0.76 abA |
| Phenylalanine 200 mg L−1 | 10.79 ± 0.88 aB | 28.99 ± 1.50 eC | 6.17 ± 0.76 bA | 3.85 ± 0.72 aA | 13.52 ± 2.84 deB |
| Tryptopan 100 mg L−1 | 28.87 ± 1.87 cC | 17.05 ± 5.89 cB | 6.04 ± 0.79 bA | 14.60 ± 3.57 bB | 14.19 ± 1.00 eB |
| Tryptopan 200 mg L−1 | 23.56 ± 1.54 bB | 25.50 ± 1.53 deB | 7.10 ± 0.82 bA | 7.11 ± 1.04 aA | 4.61 ± 0.75 aA |
| Tyrosine 100 mg L−1 | 26.84 ± 1.95 bcD | 4.61 ± 0.75 aA | 6.21 ± 0.80 bA | 16.29 ± 1.68 bC | 12.19 ± 1.00 deB |
| Tyrosine 200 mg L−1 | 22.58 ± 2.11 bC | 9.50 ± 0.89 aB | 3.65 ± 0.72 aA | 4.95 ± 0.74 aA | 10.31 ± 0.92 cdB |
| Treatment | Species/Varieties | ||||
|---|---|---|---|---|---|
| M. spicata ‘Moroccan’ | M. spicata ‘Crispa’ | M. piperita ‘Granada’ | M. piperita ‘Swiss’ | M. piperita ‘Multimentha’ | |
| Unsprayed | 14.19 ± 0.80 abB | 5.81 ± 0.37 aA | 12.74 ± 0.73 aB | 4.62 ± 0.54 aA | 4.45 ± 0.56 aA |
| Water | 15.58 ± 1.43 abB | 13.10 ± 1.03 abB | 18.22 ± 1.37 bC | 6.77 ± 0.22 cA | 6.06 ± 0.35 abA |
| Phenylalanine 100 mg L−1 | 15.39 ± 1.33 abCD | 12.97 ± 1.00 abBC | 17.44 ± 1.27 bD | 5.77 ± 0.38 bA | 11.33 ± 0.40 bB |
| Phenylalanine 200 mg L−1 | 14.83 ± 0.84 abBC | 12.55 ± 0.57 abAB | 19.50 ± 1.55 bcC | 5.76 ± 0.32 bA | 49.14 ± 5.74 dD |
| Tryptopan 100 mg L−1 | 17.04 ± 0.98 abAB | 20.37 ± 14.37 bAB | 16.36 ± 0.86 abAB | 8.07 ± 0.13 dA | 30.66 ± 3.01 cB |
| Tryptopan 200 mg L−1 | 8.80 ± 0.08 aA | 12.37 ± 0.49 abB | 28.56 ± 2.87 dC | 7.12 ± 0.36 cA | 7.39 ± 0.15 abA |
| Tyrosine 100 mg L−1 | 14.79 ± 0.81abB | 12.43 ± 0.56 abB | 23.56 ± 2.59 cC | 5.31 ± 0.45 abA | 5.28 ± 0.45 abA |
| Tyrosine 200 mg L−1 | 62.50 ± 7.09 cC | 7.64 ± 0.29 abA | 17.51 ± 1.29 bB | 10.10 ± 0.23 eAB | 8.53 ± 0.11 abA |
| Year | Month | Hydrotermal Sielianinov Coefficient | Classification by the Month, by Skowera, Pula (2004) |
|---|---|---|---|
| 2017 | April | 15.1 | Extremely wet |
| May | 0.3 | Extremely dry | |
| June | 1.6 | Quite wet | |
| July | 1.5 | Optimal | |
| August | 1.0 | Quite dry | |
| 2018 | April | 2.9 | Very wet |
| May | 0.4 | Extremely dry | |
| June | 1.1 | Quite dry | |
| July | 2.2 | Wet | |
| August | 1.1 | Quite dry | |
| 2019 | April | 0.0 | Extremely dry |
| May | 1.1 | Quite dry | |
| June | 0.8 | Dry | |
| July | 1.1 | Quite dry | |
| August | 1.2 | Quite dry |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Velička, A.; Tarasevičienė, Ž.; Hallmann, E.; Kieltyka-Dadasiewicz, A. Impact of Foliar Application of Amino Acids on Essential Oil Content, Odor Profile, and Flavonoid Content of Different Mint Varieties in Field Conditions. Plants 2022, 11, 2938. https://doi.org/10.3390/plants11212938
Velička A, Tarasevičienė Ž, Hallmann E, Kieltyka-Dadasiewicz A. Impact of Foliar Application of Amino Acids on Essential Oil Content, Odor Profile, and Flavonoid Content of Different Mint Varieties in Field Conditions. Plants. 2022; 11(21):2938. https://doi.org/10.3390/plants11212938
Chicago/Turabian StyleVelička, Aloyzas, Živilė Tarasevičienė, Ewelina Hallmann, and Anna Kieltyka-Dadasiewicz. 2022. "Impact of Foliar Application of Amino Acids on Essential Oil Content, Odor Profile, and Flavonoid Content of Different Mint Varieties in Field Conditions" Plants 11, no. 21: 2938. https://doi.org/10.3390/plants11212938