
Effect of Mycotoxin Cytochalasin A on Photosystem II in Ageratina adenophora

Abstract
:1. Introduction
2. Results and Discussion
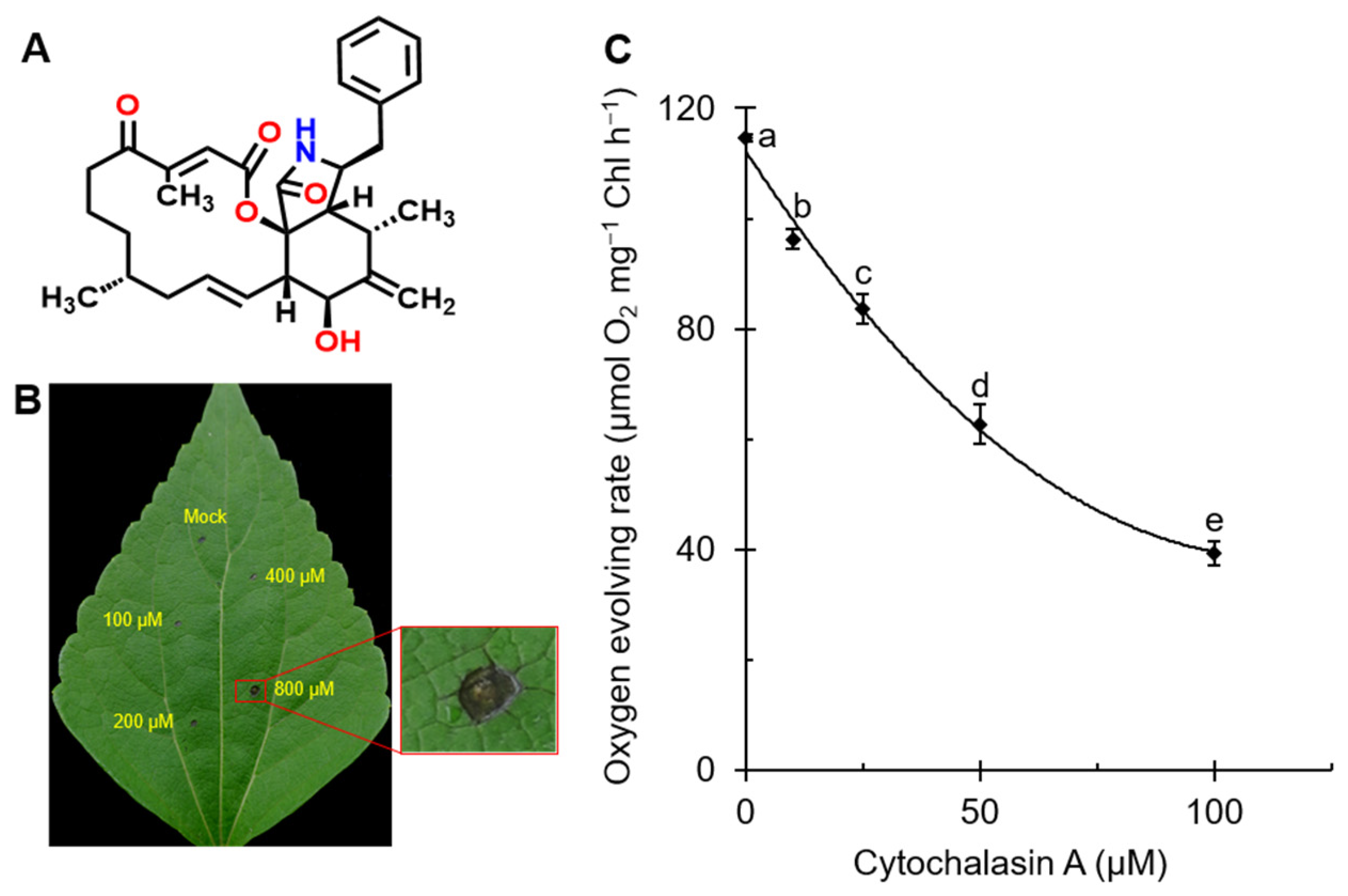
2.1. CA Caused Leaf Lesions in A. adenophora and Reduced the Rate of Oxygen Evolution in C. reinhardtii
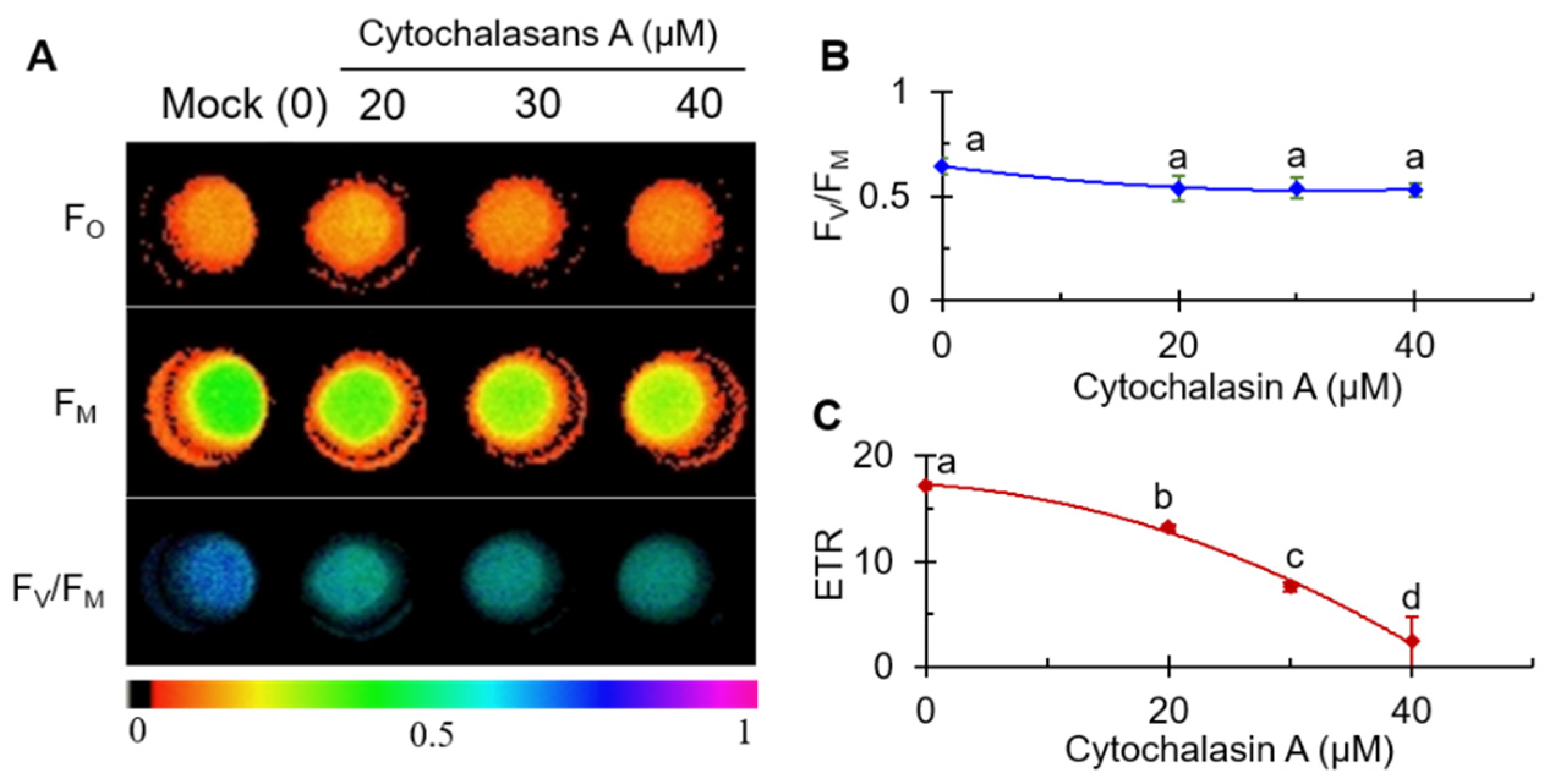
2.2. CA Inhibited Photosynthesis Activity of C. reinhardtii
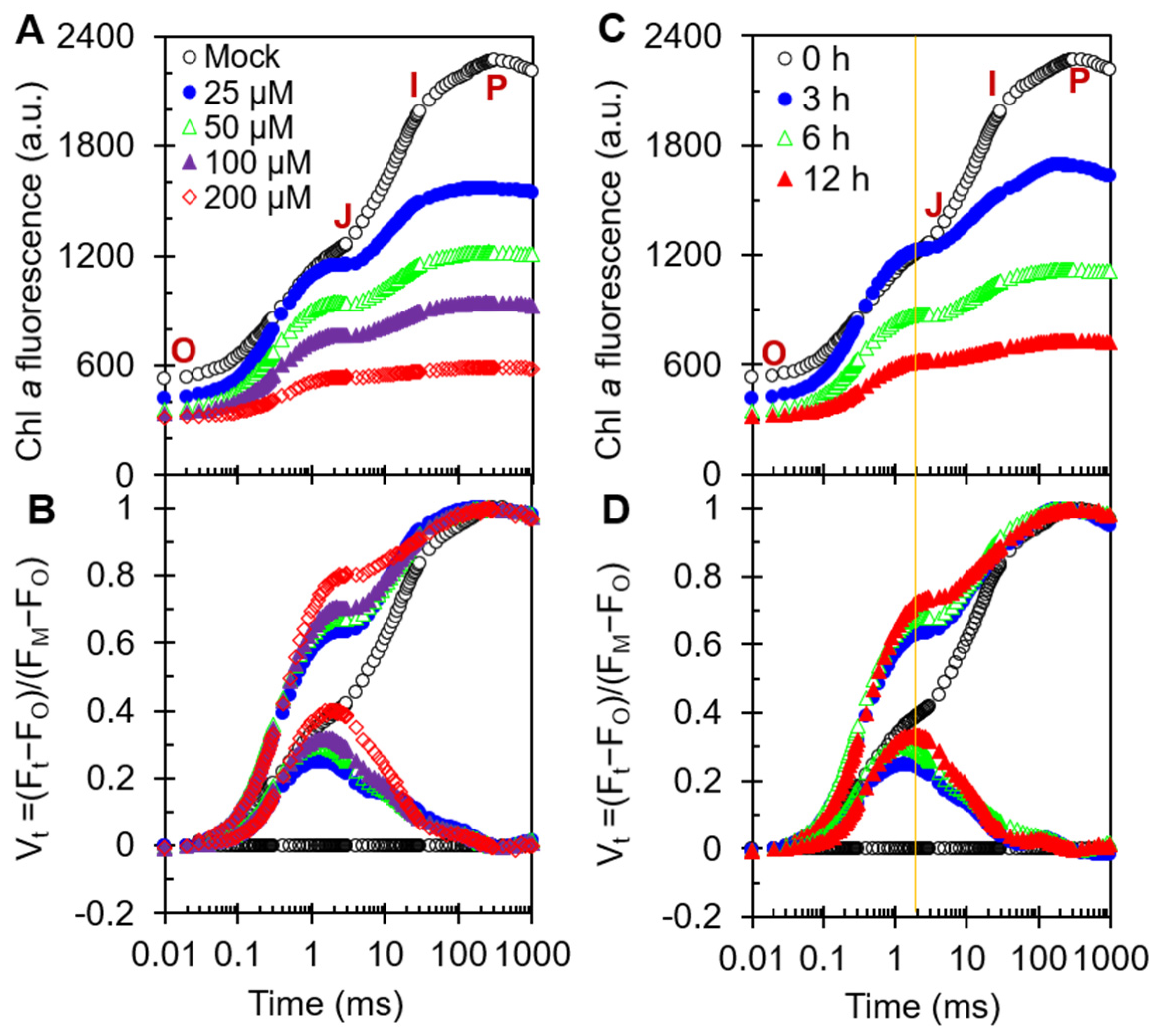
2.3. Fast Chl a Fluorescence Rise Kinetics OJIP of CA-Treated A. adenophora
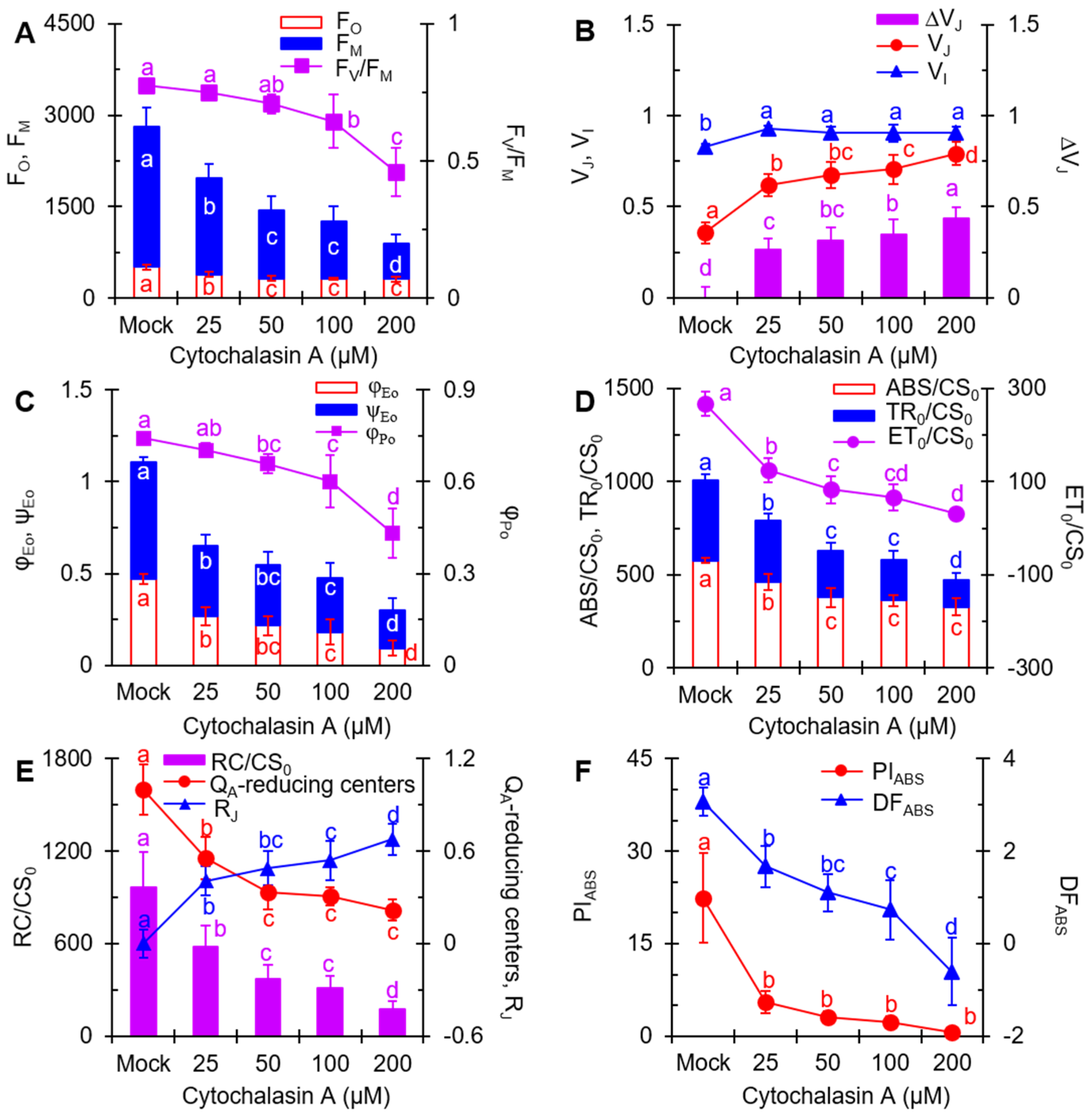
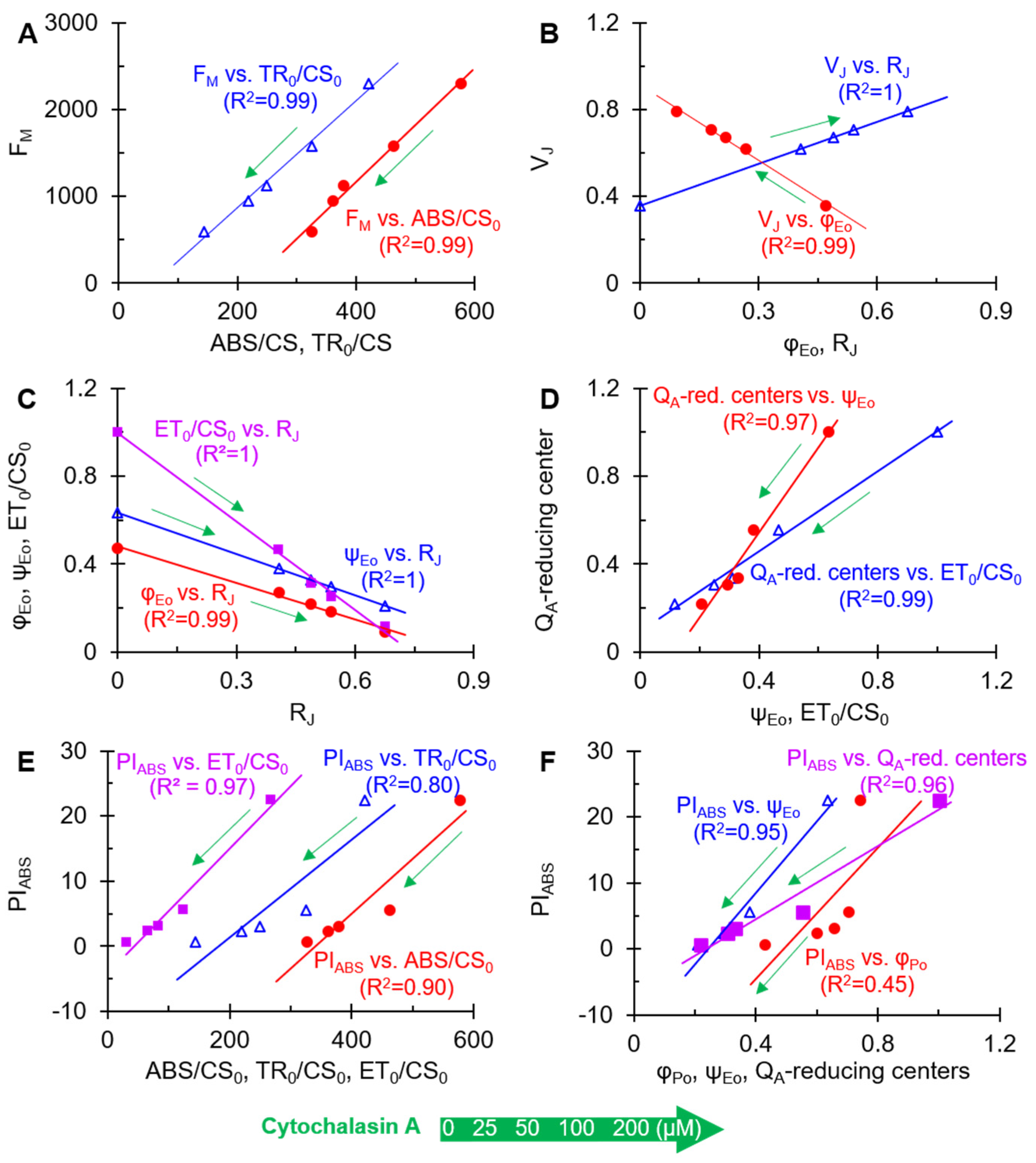
2.4. The JIP-Test Analysis
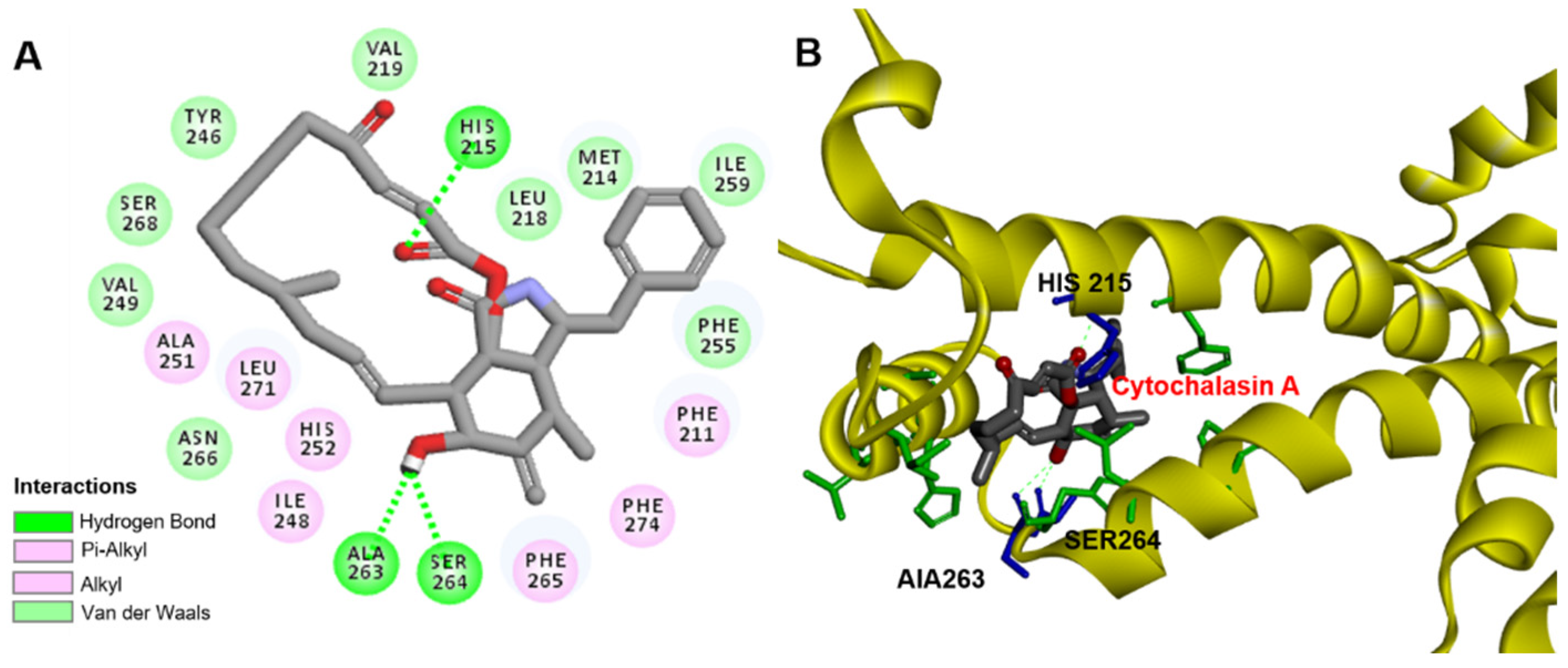
2.5. Modeling of CA Binding to D1 Protein
3. Materials and Methods
3.1. Plants and Chemicals
3.2. Phytotoxicity Assessment
3.3. Measurement of PSII Oxygen-Evolution Rate of C. reinhardtii
3.4. Chl a Fluorescence Imaging
3.5. Chl a Fluorescence Kinetics OJIP Curves and JIP-Test
3.6. Modeling of CA in D1 Protein of A. adenophora
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Green, J.M.; Owen, M.D. Herbicide-resistant crops: Utilities and limitations for herbicide-resistant weed management. J. Agric. Food Chem. 2011, 59, 5819–5829. [Google Scholar] [CrossRef] [PubMed]
- Kaur, S.; Singh, H.P.; Mittal, S.; Batish, D.R.; Kohli, R.K. Phytotoxic effects of volatile oil from Artemisia scoparia against weeds and its possible use as a bioherbicide. Ind. Crops Prod. 2010, 32, 54–61. [Google Scholar] [CrossRef]
- Huang, H.; Morgan, C.M.; Asolkar, R.N.; Koivunen, M.E.; Marrone, P.G. Phytotoxicity of sarmentine isolated from long pepper (Piper longum) fruit. J. Agric. Food Chem. 2010, 58, 9994–10000. [Google Scholar] [CrossRef] [PubMed]
- Harding, D.P.; Raizada, M.N. Controlling weeds with fungi, bacteria and viruses: A review. Front. Plant Sci. 2015, 6, 659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scherlach, K.; Boettger, D.; Remme, N.; Hertweck, C. The chemistry and biology of cytochalasans. Nat. Prod. Rep. 2010, 27, 869–886. [Google Scholar] [CrossRef] [PubMed]
- Skellam, E. The biosynthesis of cytochalasans. Nat. Prod. Rep. 2017, 34, 1252–1263. [Google Scholar] [CrossRef]
- Himes, R.H.; Houston, L.L. The action of cytochalasin A on the in vitro polymerization of brain tubulin and muscle G-actin. J. Supramol. Struct. 1976, 5, 81–90. [Google Scholar] [CrossRef]
- Peterson, J.R.; Mitchison, T.J. Small Molecules, big Impact: A history of chemical Inhibitors and the cytoskeleton. Chem. Biol. 2002, 9, 1275–1285. [Google Scholar] [CrossRef] [Green Version]
- MacLean-Fletcher, S.; Pollard, T.D. Mechanism of action of cytochalasin B on actin. Cell 1980, 20, 329–341. [Google Scholar] [CrossRef]
- Brown, S.S.; Spudich, J.A. Mechanism of action of cytochalasin: Evidence that it binds to actin filament ends. J. Cell Biol. 1981, 88, 487–491. [Google Scholar] [CrossRef]
- Cooper, J.A. Effects of cytochalasin and phalloidin on actin. J. Cell Biol. 1987, 105, 1473–1478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shepro, D.; Belamarich, F.A.; Robblee, L.; Chao, F.C. Antimotility effect of cytochalasin B observed in mammalian clot retraction. J. Cell Biol. 1970, 47, 544–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rampal, A.L.; Pinkofsky, H.B.; Jung, C.Y. Structure of cytochalasins and cytochalasin B binding sites in human erythrocyte membranes. Biochemistry 1980, 19, 679–683. [Google Scholar] [CrossRef]
- Nemeth, E.F.; Douglas, W.W. Effects of microfilament-active drugs, phalloidin and the cytochalasins A and B, on exocytosis in mast cells evoked by 48/80 or A23187. Naunyn-Schmiedeberg’s Arch. Pharmacol. 1978, 302, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Lin, D.C.; Spudich, J.A.; Kun, E. Inhibition of mitochondrial contraction by cytochalasin B. FEBS Lett. 1973, 37, 241–243. [Google Scholar]
- Gandalovicova, A.; Rosel, D.; Fernandes, M.; Vesely, P.; Heneberg, P.; Cermak, V.; Petruzelka, L.; Kumar, S.; Sanz-Moreno, V.; Brabek, J. Migrastatics-anti-metastatic and anti-invasion drugs: Promises and challenges. Trends Cancer 2017, 3, 391–406. [Google Scholar] [CrossRef] [Green Version]
- Trendowski, M.; Mitchell, J.M.; Corsette, C.M.; Acquafondata, C.; Fondy, T.P. Chemotherapy with cytochalasin congeners in vitro and in vivo against murine models. Investig. New Drug 2015, 33, 290–299. [Google Scholar] [CrossRef] [Green Version]
- Wagner, G.; Haupt, W.; Laux, A. Reversible inhibition of chloroplast movement by cytochalasin B in the green alga mougeofia. Science 1972, 176, 808–809. [Google Scholar] [CrossRef]
- Izutani, Y.; Takagi, S.; Nag, R. Orientation movements of chloroplasts in Vallisneriu epidermal cells: Different effects of light at low-and high-fluence rate. Photochem. Photobiol. 1990, 51, 105–111. [Google Scholar] [CrossRef]
- Gestel, K.V.; Kohler, R.H.; Verbelen, J.P. Plant mitochondria move on F-actin, but their positioning in the cortical cytoplasm depends on both F-actin and microtubules. J. Exp. Bot. 2002, 53, 659–667. [Google Scholar] [CrossRef]
- Quader, H.; Hofmann, A.; Schnepf, E. Reorganization of the endoplasmic reticulum in epidermal cells of onion bulb scales after cold stress: Involvement of cytoskeletal elements. Planta 1989, 177, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Boevink, P.; Oparka, K.; Cruz, S.S.; Martin, B.; Betteridge, A.; Hawes, C. Stacks on tracks: The plant Golgi apparatus traffics on an actin/ER network. Plant J. 1998, 15, 441–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nebenfuhr, A.; Gallagher, L.A.; Dunahay, T.G.; Frohlick, J.A.; Mazurkiewicz, A.M.; Meehl, J.B.; Staehelin, L.A. Stop-and-go movements of plant Golgi stacks are mediated by the acto-myosin system. Plant Physiol. 1999, 121, 1127–1141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mano, S.; Nakamori, C.; Hayashi, M.; Kato, A.; Kondo, M.; Nishimura, M. Distribution and characterization of peroxisomes in Arabidopsis by visualization with GFP: Dynamic morphology and actin-dependent movement. Plant Cell Physiol. 2002, 43, 331–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalachova, T.; Leontovycova, H.; Iakovenko, O.; Pospichalova, R.; Marsik, P.; Kloucek, P.; Janda, M.; Valentova, O.; Kocourkova, D.; Martinec, J.; et al. Interplay between phosphoinositides and actin cytoskeleton in the regulation of immunity related responses in Arabidopsis thaliana seedlings. Environ. Exp. Bot. 2019, 167, 103867. [Google Scholar] [CrossRef]
- Leontovycova, H.; Kalachova, T.; Trda, L.; Pospichalova, R.; Lamparova, L.; Dobrev, P.I.; Malinska, K.; Burketova, L.; Valentova, O.; Janda, M. Actin depolymerization is able to increase plant resistance against pathogens via activation of salicylic acid signalling pathway. Sci. Rep. 2019, 9, 10397. [Google Scholar] [CrossRef] [Green Version]
- Evidente, A.; Andolfi, A.; Vurro, M.; Zonno, M.C.; Motta, A. Cytochalasins Z1, Z2 and Z3, three 24-oxal[14]cytochalasans produced by Pyrenophora semeniperda. Phytochemistry 2002, 60, 45–53. [Google Scholar] [CrossRef]
- Berestetskiy, A.; Dmitriev, A.; Mitina, G.; Lisker, I.; Andolfi, A.; Evidente, A. Nonenolides and cytochalasins with phytotoxic activity against Cirsium arvense and Sonchus arvensis: A structure-activity relationships study. Phytochemistry 2008, 69, 953–960. [Google Scholar] [CrossRef]
- Konig, G.M.; Wright, A.D.; Aust, H.J.; Draeger, S.; Schulz, B. Geniculol, a new biologically active diterpene from the endophytic fungus Geniculosporium sp. J. Nat. Prod. 1999, 62, 155–157. [Google Scholar] [CrossRef]
- Kshirsagar, A.; Reid, A.J.; McColl, S.M.; Saunders, V.A.; Whalley, A.J.S.; Evans, E.H. The effect of fungal metabolites on leaves as detected by chlorophyll fluorescence. New Phytol. 2001, 151, 451–457. [Google Scholar] [CrossRef]
- Shen, J.R. The structure of photosystem II and the mechanism of water oxidation in photosynthesis. Annu. Rev. Plant Biol. 2015, 66, 23–48. [Google Scholar] [CrossRef] [PubMed]
- Oettmeier, W. Herbicide resistance and supersensitivity in photosystem II. Cell. Mol. Life Sci. 1999, 55, 1255–1277. [Google Scholar] [CrossRef] [PubMed]
- Trebst, A. The Mode of Action of Triazine Herbicides in Plants; Elsevier: San Diego, CA, USA, 2008; pp. 101–110. [Google Scholar]
- Battaglino, B.; Grinzato, A.; Pagliano, C. Binding properties of photosynthetic herbicides with the QB site of the D1 protein in plant photosystem II: A combined functional and molecular docking study. Plants 2021, 10, 1501. [Google Scholar] [CrossRef] [PubMed]
- Vurro, M.; Ellis, B.E. Effect of fungal toxins on induction of phenylalanine ammonia-lyase activity in elicited cultures of hybrid poplar. Plant Sci. 1997, 126, 29–38. [Google Scholar] [CrossRef]
- Guo, Y.; Cheng, J.; Lu, Y.; Wang, H.; Gao, Y.; Shi, J.; Yin, C.; Wang, X.; Chen, S.; Strasser, R.J.; et al. Novel action targets of natural product gliotoxin in photosynthetic apparatus. Front. Plant Sci. 2020, 10, 1688. [Google Scholar] [CrossRef] [Green Version]
- Xiao, W.; Wang, H.; Liu, W.; Wang, X.; Guo, Y.; Strasser, R.J.; Qiang, S.; Chen, S.; Hu, Z. Action of alamethicin in photosystem II probed by the fast chlorophyll fluorescence rise kinetics and the JIP-test. Photosynthetica 2020, 58, 358–368. [Google Scholar] [CrossRef]
- Guo, Y.; Liu, W.; Wang, H.; Wang, X.; Qiang, S.; Kalaji, H.M.; Strasser, R.J.; Chen, S. Action mode of the mycotoxin patulin as a novel natural photosystem II Inhibitor. J. Agric. Food Chem. 2021, 69, 7313–7323. [Google Scholar] [CrossRef]
- Chen, S.; Xu, X.; Dai, X.; Yang, C.; Qiang, S. Identification of tenuazonic acid as a novel type of natural photosystem II inhibitor binding in QB-site of Chlamydomonas reinhardtii. Biochim. Biophys. Acta 2007, 1767, 306–318. [Google Scholar] [CrossRef] [Green Version]
- Nimbal, C.I.; Yerkes, C.N.; Weston, L.A.; Weller, S.C. Herbicidal activity and site of action of the natural product sorgoleone. Pestic. Biochem. Phys. 1996, 54, 73–83. [Google Scholar] [CrossRef]
- Strasser, R.J.; Tsimilli-Michael, M.; Srivastava, A. Analysis of the chlorophyll a fluorescence transient. In Chlorophyll a Fluorescence: A Signature of Photosynthesis; Papageorgiou, G.C., Ed.; Springer: Dordrecht, The Netherlands, 2004; Volume 19, pp. 321–362. [Google Scholar]
- Krause, G.H.; Weis, E. Chlorophyll fluorescence and photosynthesis: The basics. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1991, 42, 313–349. [Google Scholar] [CrossRef]
- Toth, S.Z.; Schansker, G.; Strasser, R.J. In intact leaves, the maximum fluorescence level FM is independent of the redox state of the plastoquinone pool: A DCMU-inhibition study. Biochim. Biophys. Acta 2005, 1708, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Liu, W.; Wang, X.; Yang, L.; Han, S.; Chen, S.; Strasser, R.J.; Valverde, B.E.; Qiang, S. Comparative phytotoxicity of usnic acid, salicylic acid, cinnamic acid and benzoic acid on photosynthetic apparatus of Chlamydomonas reinhardtii. Plant Physiol. Biochem. 2018, 128, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Strasser, R.J.; Qiang, S. In vivo assessment of effect of phytotoxin tenuazonic acid on PSII reaction centers. Plant Physiol. Biochem. 2014, 84, 10–21. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Yang, J.; Zhang, M.; Strasser, R.J.; Qiang, S. Classification and characteristics of heat tolerance in Ageratina adenophora populations using fast chlorophyll a fluorescence rise O-J-I-P. Environ. Exp. Bot. 2016, 122, 126–140. [Google Scholar] [CrossRef]
- Kyle, D.J. The 32000 dalton QB protein of photosystem II. Photochem. Photobiol. 1985, 41, 107–116. [Google Scholar] [CrossRef]
- Xiong, J.; Subramaniam, S.; Govindjee. Modeling of the D1/D2 proteins and cofactors of the photosystem II reaction center: Implications for herbicide and bicarbonate binding. Protein Sci. 1996, 5, 2054–2073. [Google Scholar] [CrossRef]
- Sun, H. Homology modeling and ligand-based molecule design. In A Practical Guide to Rational Drug Design; Sun, H., Ed.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 109–160. [Google Scholar]
- Wang, H.; Yao, Q.; Guo, Y.; Zhang, Q.; Wang, Z.; Strasser, R.J.; Valverde, B.E.; Chen, S.; Qiang, S.; Kalaji, H.M. Structure-based ligand design and discovery of novel tenuazonic acid derivatives with high herbicidal activity. J. Adv. Res. 2021, 40, 29–44. [Google Scholar] [CrossRef]
- Takahashi, R.; Hasegawa, K.; Takano, A.; Noguchi, T. Structures and binding sites of phenolic herbicides in the QB pocket of photosystem II. Biochemistry 2010, 49, 5445–5454. [Google Scholar] [CrossRef] [PubMed]
- Strasser, R.J.; Tsimilli-Michael, M.; Qiang, S.; Goltsev, V. Simultaneous in vivo recording of prompt and delayed fluorescence and 820-nm reflection changes during drying and after rehydration of the resurrection plant Haberlea rhodopensis. Biochim. Biophys. Acta Bioenerg. 2010, 1797, 1313–1326. [Google Scholar] [CrossRef] [Green Version]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Technical Fluorescence Parameters | |
|---|---|
| Ft | fluorescence at time t after onset of actinic illumination |
| FO ≅ F20μs | minimal fluorescence, when all PSII RCs are open |
| FL ≡ F150μs | fluorescence intensity at the L-step (150 μs) of OJIP |
| FK ≡ F300μs | fluorescence intensity at the K-step (300 μs) of OJIP |
| FJ ≡ F2ms | fluorescence intensity at the J-step (2 ms) of OJIP |
| FI ≡ F30ms | fluorescence intensity at the I-step (30 ms) of OJIP |
| FP (= M) | maximal fluorescence, at the peak P of OJIP |
| Fv ≡ Ft − FO | variable fluorescence at time t |
| Vt ≡ (Ft − FO)/(FM − FO) | relative variable fluorescence at time t |
| VJ = (FJ − FO)/(FM − FO) | relative variable fluorescence at the J-step |
| VI = (FI − FO)/(FM − FO) | relative variable fluorescence at the I-step |
| Quantum efficiencies or flux ratios | |
| φPo = PHI(P0) = TR0/ABS = 1 − FO/FM | maximum quantum yield for primary photochemistry |
| ψΕo = PSI0 = ET0/TR0 = (1 − VJ) | probability that an electron moves further than QA− |
| φEo = PHI(E0) = ET0/ABS = (1 − FO/FM) (1 − VJ) | quantum yield for electron transport (ET) |
| Phenomenological energy fluxes (per excited leaf cross-section-CS) | |
| ABS/CS = Chl/CS | absorption flux per CS |
| TR0/CS = φPo·(ABS/CS) | trapped energy flux per CS |
| ET0/CS = φPo·ψΕo·(ABS/CS) | electron transport flux per CS |
| Density of reaction center (QA-reducing PSII reaction center–RC) | |
| RC/CS = φPo·(VJ/M0)·(ABS/CS) | density of QA-reducing PSII RCs per CS |
| QA-reducing centers = (RC/RCreference)·(ABS/ABSreference) = [(RC/CS)treatment/(RC/CS)control]·[(ABS/CS)treatment/(ABS/CS)control] | fraction of QA-reducing PSII RCs |
| RJ = (ψΕo(control) − ψΕo(treatment))/ψΕo(control) = (VJ(treatment) − VJ(control))/(1 − VJ(control)) | number of PSII RCs with QB-site filled by PSII inhibitor |
| Performance indexes | |
| ≡ | performance index for energy conservation from photons absorbed by PSII to the reduction of intersystem electron acceptors |
| Phenomenological energy fluxes (per excited leaf cross-section-CS) | |
| driving force on absorption basis | |
| CA Chemical Structure | Donor | Acceptor | Interactions | Bound Distance (Å) |
|---|---|---|---|---|
 | D1-Phe 211 | CAC16-CH3 | Pi Hydrophobic | 3.97 |
| D1-His 215 NH | CA O3 | Hydrogen Bond | 2.48 | |
| D1-Ile 248 | CA C10-CH3 | Alkyl Hydrophobic | 3.72 | |
| D1-Leu 251 | CA C11 | Alkyl Hydrophobic | 3.77 | |
| D1-His 252 | CA C10-CH3 | Alkyl Hydrophobic | 3.81 | |
| CA C18-OH | D1-Ala 263 CO | Hydrogen Bond | 2.44 | |
| CA C18-OH | D1-Ser 264 CO | Hydrogen Bond | 2.35 | |
| D1-Phe 265 | CA C17=CH2 | Pi Hydrophobic | 3.83 | |
| D1-Leu 271 | CA C11 | Alkyl Hydrophobic | 3.91 | |
| D1-Phe 274 | CA C16-CH3 | Pi Hydrophobic | 3.86 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, M.; Yang, Q.; Wang, H.; Luo, Z.; Guo, Y.; Shi, J.; Wang, X.; Qiang, S.; Strasser, R.J.; Chen, S. Effect of Mycotoxin Cytochalasin A on Photosystem II in Ageratina adenophora. Plants 2022, 11, 2797. https://doi.org/10.3390/plants11202797
Jiang M, Yang Q, Wang H, Luo Z, Guo Y, Shi J, Wang X, Qiang S, Strasser RJ, Chen S. Effect of Mycotoxin Cytochalasin A on Photosystem II in Ageratina adenophora. Plants. 2022; 11(20):2797. https://doi.org/10.3390/plants11202797
Chicago/Turabian StyleJiang, Mengyun, Qian Yang, He Wang, Zhi Luo, Yanjing Guo, Jiale Shi, Xiaoxiong Wang, Sheng Qiang, Reto Jörg Strasser, and Shiguo Chen. 2022. "Effect of Mycotoxin Cytochalasin A on Photosystem II in Ageratina adenophora" Plants 11, no. 20: 2797. https://doi.org/10.3390/plants11202797