
The Interaction between Hydromulching and Arbuscular Mycorrhiza Improves Escarole Growth and Productivity by Regulating Nutrient Uptake and Hormonal Balance

 , , , ,
, , , ,  and
and 
Abstract
:1. Introduction
2. Results
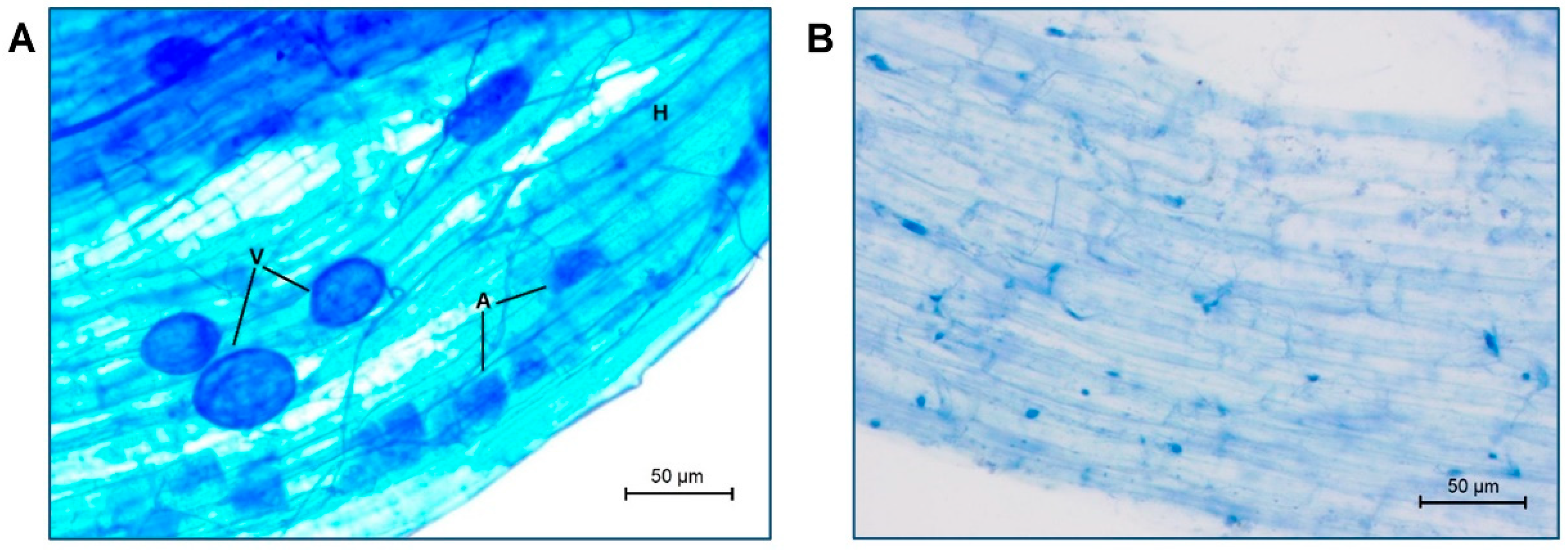
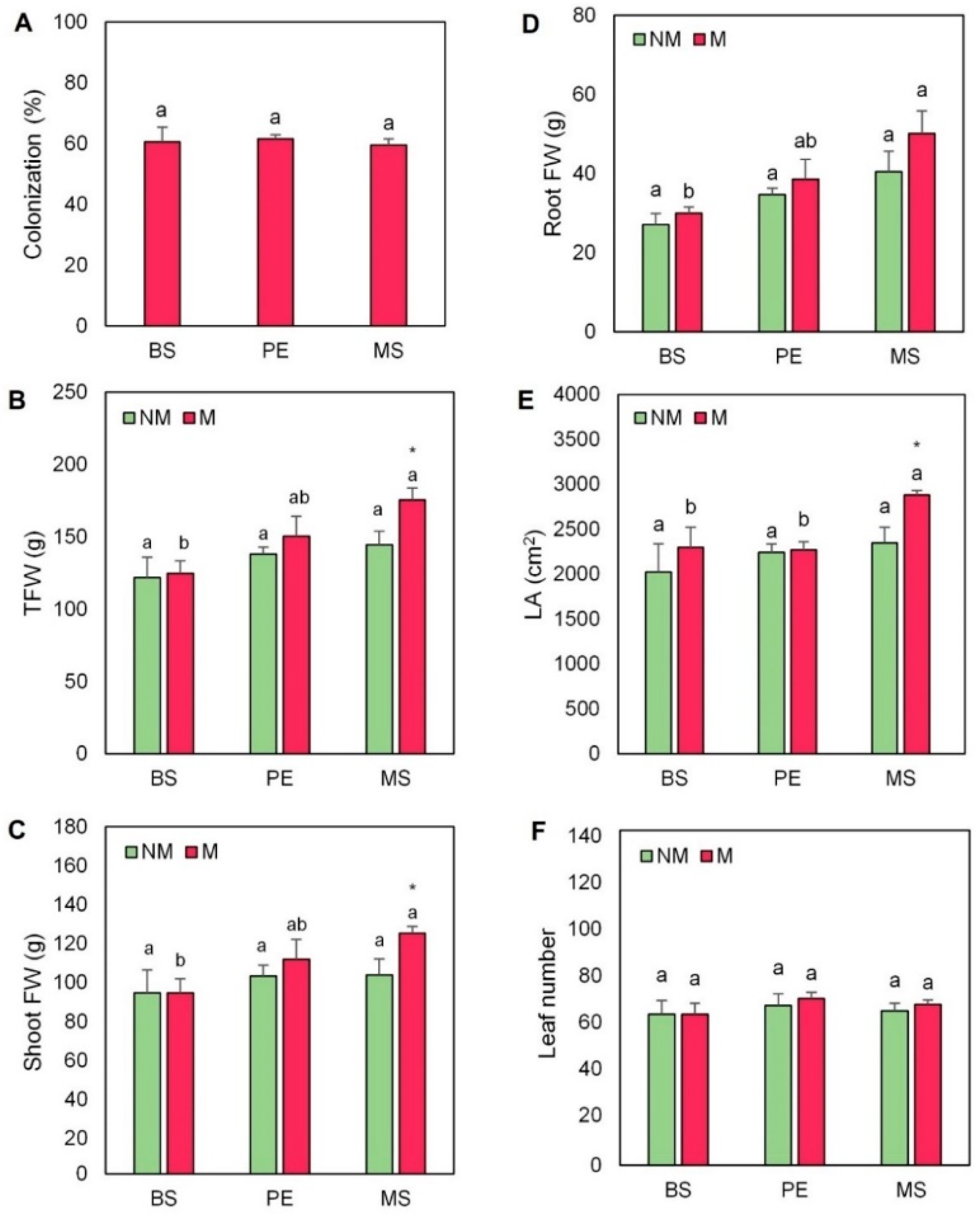
2.1. Mycorrhizal Colonization and Growth-Related Determinations
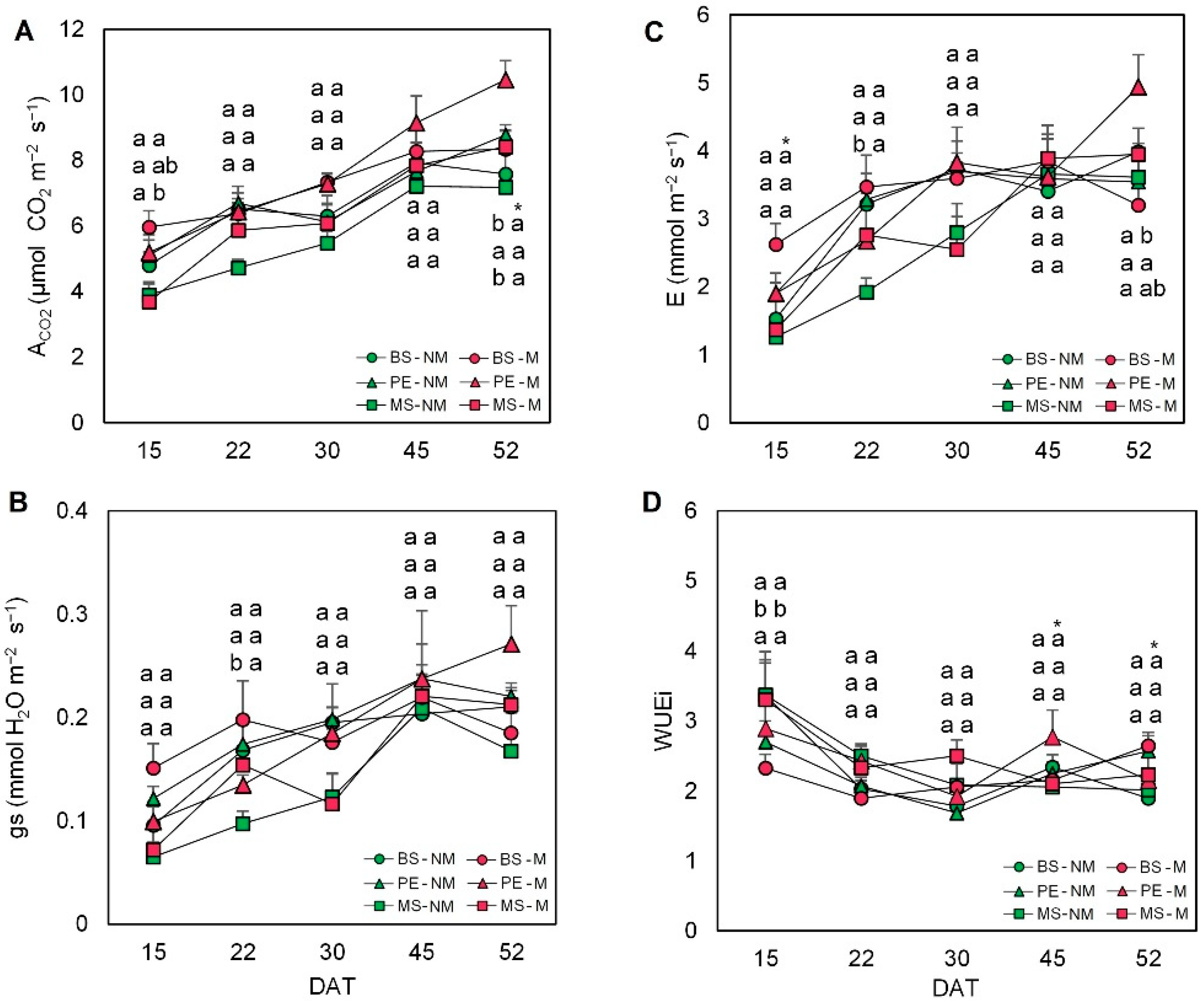
2.2. Gas Exchange Measurements
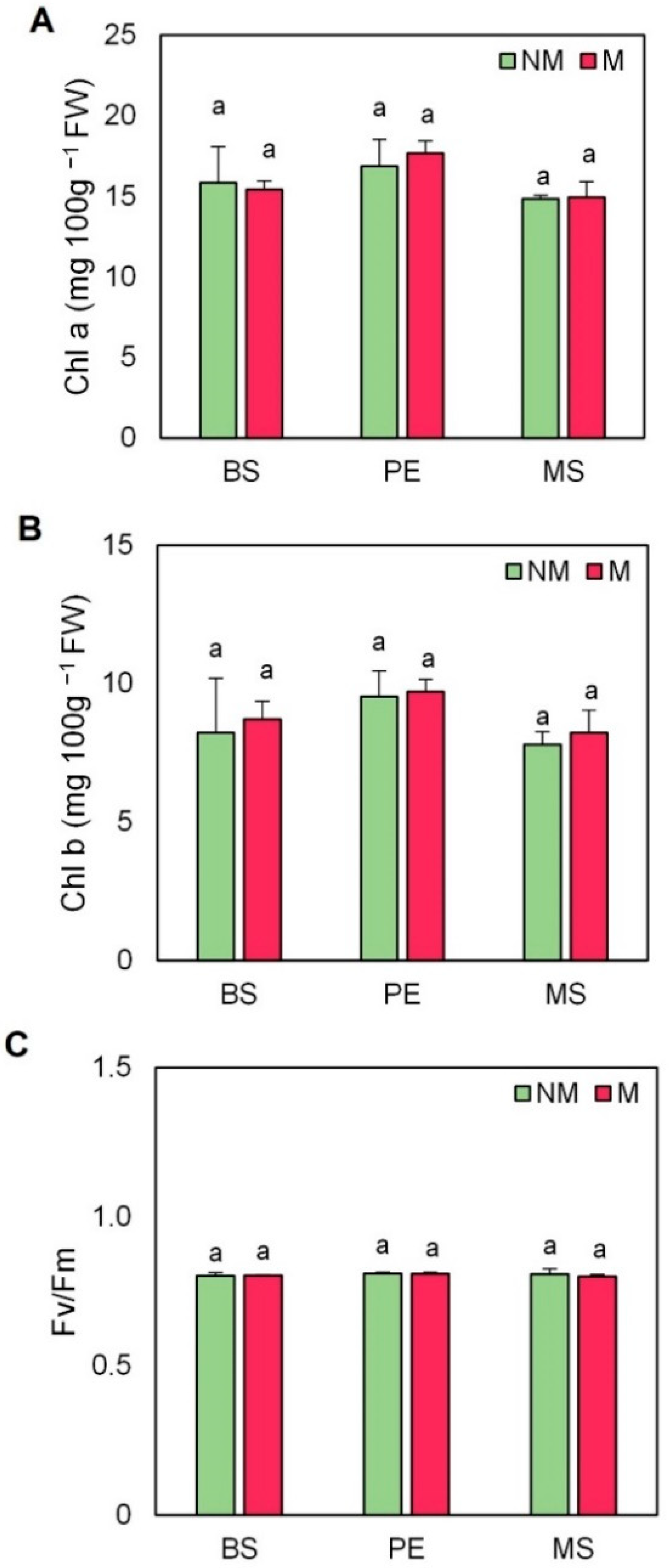
2.3. Chlorophyll Content and Fluorescence
2.4. Plant Mineral Content and Nutrient Use Efficiency
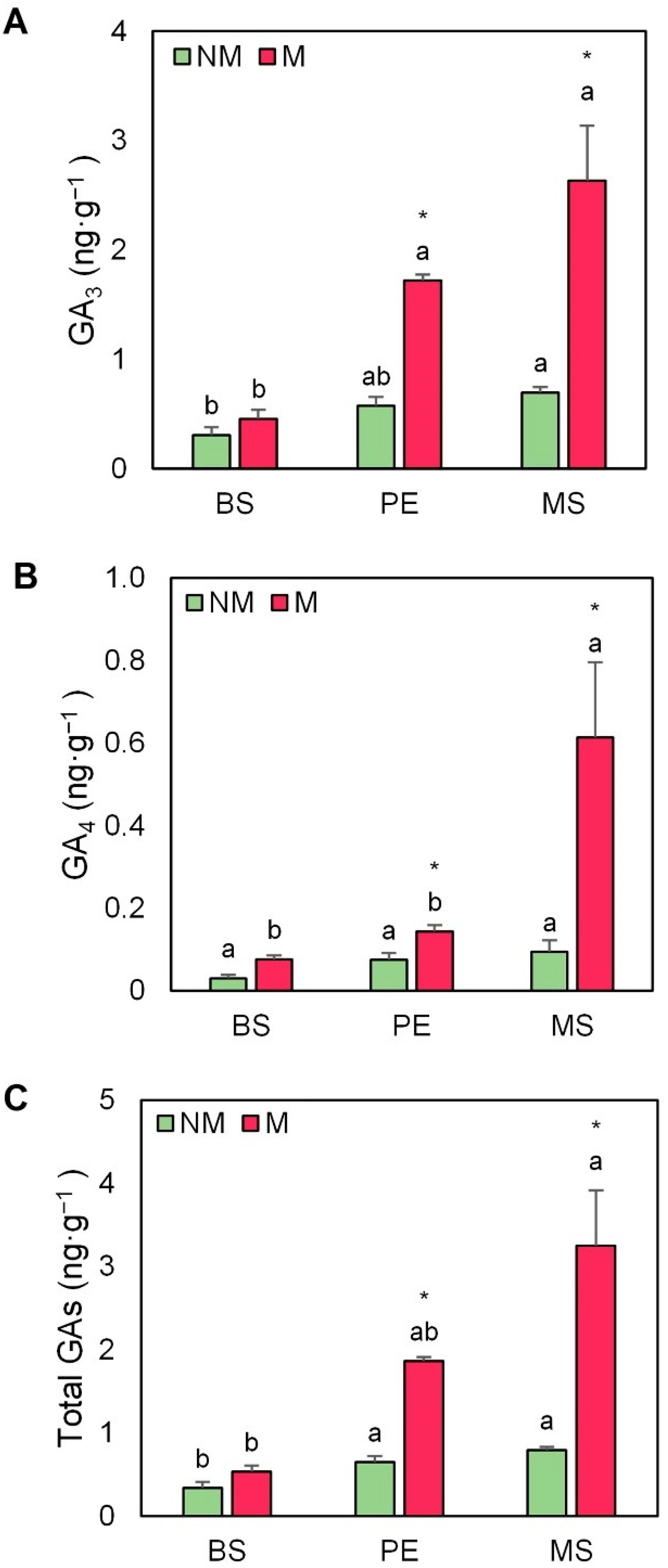
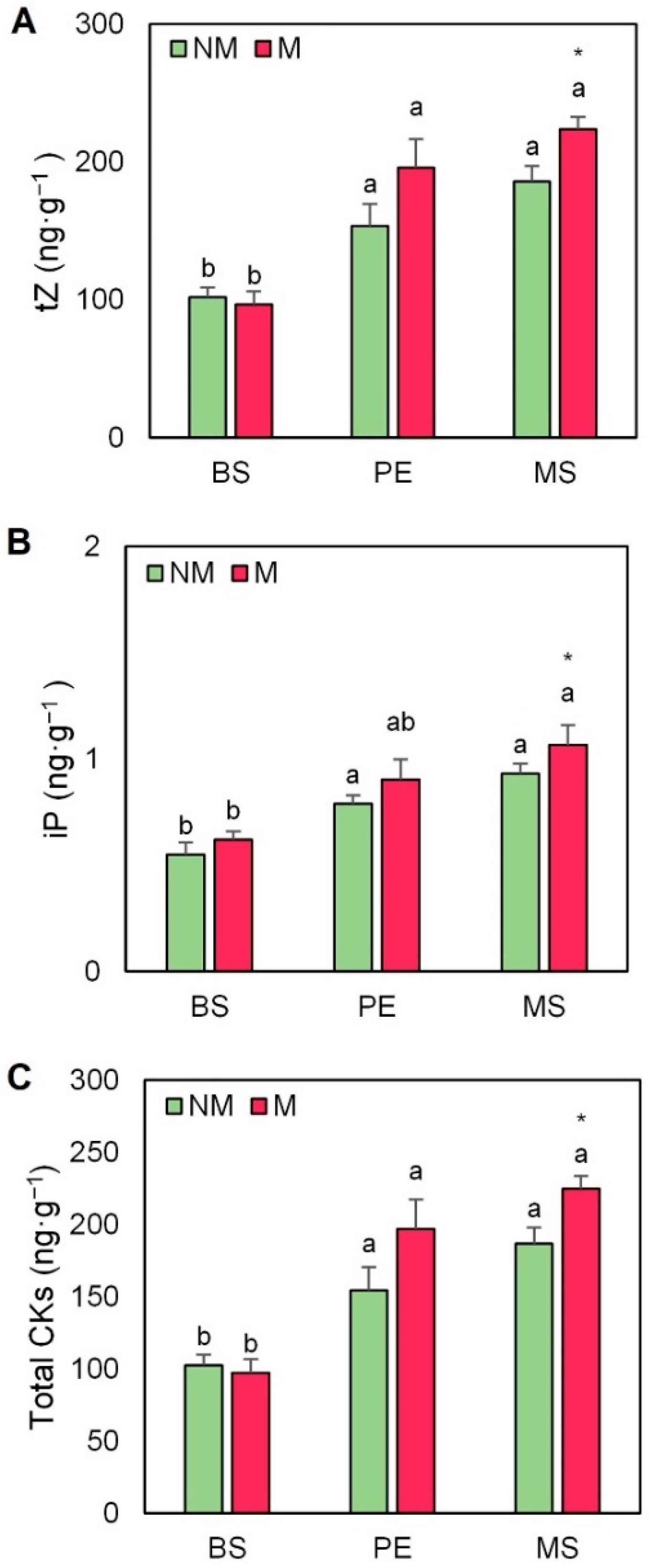
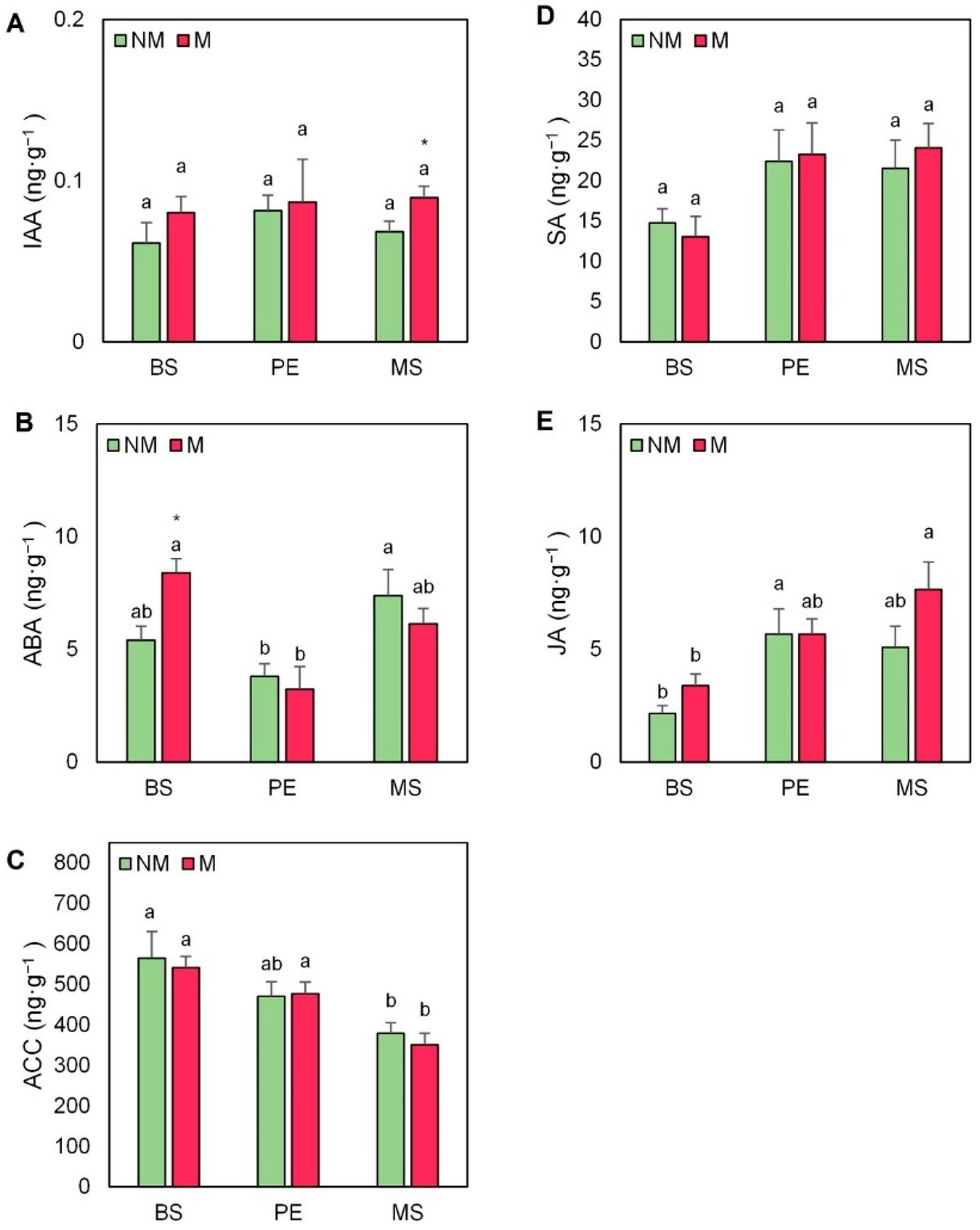
2.5. Hormonal Profile
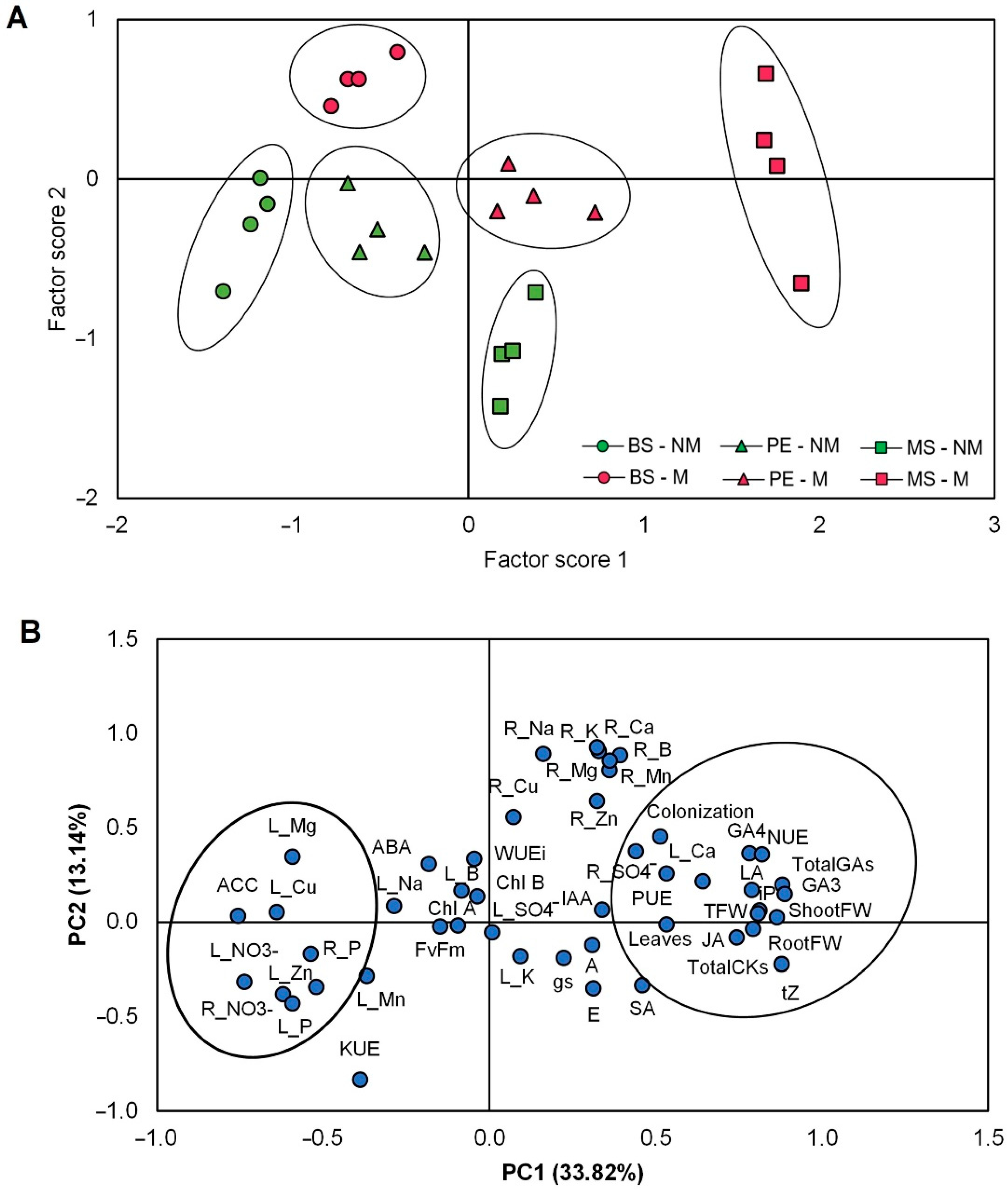
2.6. Principal Component Analysis
3. Discussion
4. Materials and Methods
4.1. Biological Material and Experimental Design
4.2. Plant Growth-Related Determinations
4.3. AM Fungal Root Colonization
4.4. Gas Exchange Measurements
4.5. Chlorophyll Content
4.6. Maximum Potential Quantum Efficiency of PSII
4.7. Plant Mineral Content
4.8. Hormone Extraction and Analysis
4.9. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Filippo D’Antuono, L.; Ferioli, F.; Manco, M.A. The Impact of Sesquiterpene Lactones and Phenolics on Sensory Attributes: An Investigation of a Curly Endive and Escarole Germplasm Collection. Food Chem. 2016, 199, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Romero-Gámez, M.; Audsley, E.; Suárez-Rey, E.M. Life Cycle Assessment of Cultivating Lettuce and Escarole in Spain. J. Clean. Prod. 2014, 73, 193–203. [Google Scholar] [CrossRef]
- FAOSTAT—Statistics Division of FAO (Food and Agriculture Organization of the United Nations). Production: Crops and Livestock Products. 2022. Available online: https://www.fao.org/faostat/es/#data/QCL/visualize (accessed on 12 October 2022).
- Noya, I.; González-García, S.; Bacenetti, J.; Fiala, M.; Moreira, M.T. Environmental Impacts of the Cultivation-Phase Associated with Agricultural Crops for Feed Production. J. Clean. Prod. 2018, 172, 3721–3733. [Google Scholar] [CrossRef]
- Poveda, J.; Hermosa, R.; Monte, E.; Nicolás, C. Trichoderma Harzianum Favours the Access of Arbuscular Mycorrhizal Fungi to Non-Host Brassicaceae Roots and Increases Plant Productivity. Sci. Rep. 2019, 9, 11650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Heijden, M.G.A.; Streitwolf-Engel, R.; Riedl, R.; Siegrist, S.; Neudecker, A.; Ineichen, K.; Boller, T.; Wiemken, A.; Sanders, I.R. The Mycorrhizal Contribution to Plant Productivity, Plant Nutrition and Soil Structure in Experimental Grassland. New Phytol. 2006, 172, 739–752. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Hardan, A.N. Importance of Mycorrhizae in Crop Productivity. In Mitigating Environmental Stresses for Agricultural Sustainability in Egypt; Springer: Berlin/Heidelberg, Germany, 2021; pp. 471–484. [Google Scholar]
- Li, X.; Zeng, R.; Liao, H. Improving Crop Nutrient Efficiency through Root Architecture Modifications. J. Integr. Plant Biol. 2016, 58, 193–202. [Google Scholar] [CrossRef] [Green Version]
- Mitra, D.; Uniyal, N.; Panneerselvam, P.; Senapati, A.; Ganeshamurthy, A.N. Role of Mycorrhiza and Its Associated Bacteria on Plant Growth Promotion and Nutrient Management in Sustainable Agriculture. IJLSAS 2020, 1, 1. [Google Scholar]
- Balliu, A.; Sallaku, G.; Rewald, B. AMF Inoculation Enhances Growth and Improves the Nutrient Uptake Rates of Transplanted, Salt-Stressed Tomato Seedlings. Sustainability 2015, 7, 15967–15981. [Google Scholar] [CrossRef] [Green Version]
- Begum, N.; Qin, C.; Ahanger, M.A.; Raza, S.; Khan, M.I.; Ashraf, M.; Ahmed, N.; Zhang, L. Role of Arbuscular Mycorrhizal Fungi in Plant Growth Regulation: Implications in Abiotic Stress Tolerance. Front. Plant. Sci. 2019, 10, 1068. [Google Scholar] [CrossRef] [Green Version]
- Bonfante, P.; Genre, A. Mechanisms Underlying Beneficial Plant—Fungus Interactions in Mycorrhizal Symbiosis. Nat. Commun. 2010, 1, 48. [Google Scholar] [CrossRef] [Green Version]
- Hashem, A.; Alqarawi, A.A.; Radhakrishnan, R.; Al-Arjani, A.B.F.; Aldehaish, H.A.; Egamberdieva, D.; Abd Allah, E.F. Arbuscular Mycorrhizal Fungi Regulate the Oxidative System, Hormones and Ionic Equilibrium to Trigger Salt Stress Tolerance in Cucumis Sativus L. Saudi J. Biol. Sci. 2018, 25, 1102–1114. [Google Scholar] [CrossRef] [PubMed]
- Kaur, S.; Suseela, V. Unraveling Arbuscular Mycorrhiza-Induced Changes in Plant Primary and Secondary Metabolome. Metabolites 2020, 10, 335. [Google Scholar] [CrossRef] [PubMed]
- Liao, D.; Wang, S.; Cui, M.; Liu, J.; Chen, A.; Xu, G. Phytohormones Regulate the Development of Arbuscular Mycorrhizal Symbiosis. Int. J. Mol. Sci. 2018, 19, 3146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pozo, M.J.; López-Ráez, J.A.; Azcón-Aguilar, C.; García-Garrido, J.M. Phytohormones as Integrators of Environmental Signals in the Regulation of Mycorrhizal Symbioses. New Phytol. 2015, 205, 1431–1436. [Google Scholar] [CrossRef]
- Gutjahr, C. Phytohormone Signaling in Arbuscular Mycorhiza Development. Curr. Opin. Plant Biol. 2014, 20, 26–34. [Google Scholar] [CrossRef]
- Foo, E.; Ross, J.J.; Jones, W.T.; Reid, J.B. Plant Hormones in Arbuscular Mycorrhizal Symbioses: An Emerging Role for Gibberellins. Ann. Bot. 2013, 111, 769–779. [Google Scholar] [CrossRef]
- Khalloufi, M.; Martínez-Andújar, C.; Lachaâl, M.; Karray-Bouraoui, N.; Pérez-Alfocea, F.; Albacete, A. The Interaction between Foliar GA3 Application and Arbuscular Mycorrhizal Fungi Inoculation Improves Growth in Salinized Tomato (Solanum Lycopersicum L.) Plants by Modifying the Hormonal Balance. J. Plant Physiol. 2017, 214, 134–144. [Google Scholar] [CrossRef]
- Romero-Muñoz, M.; Albacete, A.; Gálvez, A.; Piñero, M.C.; del Amor, F.M.; López-Marín, J. The Use of Ecological Hydromulching Improves Growth in Escarole (Cichorium Endivia L.) Plants Subjected to Drought Stress by Fine-Tuning Cytokinins and Abscisic Acid Balance. Agronomy 2022, 12, 459. [Google Scholar] [CrossRef]
- Gálvez, A.; Albacete, A.; del Amor, F.M.; López-Marín, J. The Use of Red Shade Nets Improves Growth in Salinized Pepper (Capsicum Annuum L.) Plants by Regulating Their Ion Homeostasis and Hormone Balance. Agronomy 2020, 10, 1766. [Google Scholar] [CrossRef]
- Shaul-Keinan, O.; Gadkar, V.; Ginzberg, I.; Grünzweig, J.M.; Chet, I.; Elad, Y.; Wininger, S.; Belausov, E.; Eshed, Y.; Atzmon, N.; et al. Hormone Concentrations in Tobacco Roots Change during Arbuscular Mycorrhizal Colonization with Glomus Intraradices. New Phytol. 2002, 154, 501–507. [Google Scholar] [CrossRef]
- Barker, S.J.; Tagu, D. The Roles of Auxins and Cytokinins in Mycorrhizal Symbioses. J. Plant Growth Regul. 2000, 19, 144–154. [Google Scholar] [CrossRef]
- Cosme, M.; Ramireddy, E.; Franken, P.; Schmülling, T.; Wurst, S. Shoot- and Root-Borne Cytokinin Influences Arbuscular Mycorrhizal Symbiosis. Mycorrhiza 2016, 26, 709–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Medina, A.; Roldán, A.; Albacete, A.; Pascual, J.A. The Interaction with Arbuscular Mycorrhizal Fungi or Trichoderma Harzianum Alters the Shoot Hormonal Profile in Melon Plants. Phytochemistry 2011, 72, 223–229. [Google Scholar] [CrossRef]
- Jentschel, K.; Thiel, D.; Rehn, F.; Ludwig-Müller, J. Arbuscular Mycorrhiza Enhances Auxin Levels and Alters Auxin Biosynthesis in Tropaeolum Majus during Early Stages of Colonization. Physiol. Plant 2007, 129, 320–333. [Google Scholar] [CrossRef]
- Meixner, C.; Ludwig-Müller, J.; Miersch, O.; Gresshoff, P.; Staehelin, C.; Vierheilig, H. Lack of Mycorrhizal Autoregulation and Phytohormonal Changes in the Supernodulating Soybean Mutant Nts1007. Planta 2005, 222, 709–715. [Google Scholar] [CrossRef] [PubMed]
- Stumpe, M.; Carsjens, J.G.; Stenzel, I.; Göbel, C.; Lang, I.; Pawlowski, K.; Hause, B.; Feussner, I. Lipid Metabolism in Arbuscular Mycorrhizal Roots of Medicago Truncatula. Phytochemistry 2005, 66, 781–791. [Google Scholar] [CrossRef] [Green Version]
- Miransari, M.; Abrishamchi, A.; Khoshbakht, K.; Niknam, V. Plant Hormones as Signals in Arbuscular Mycorrhizal Symbiosis. Crit. Rev. Biotechnol. 2014, 34, 123–133. [Google Scholar] [CrossRef]
- Iqbal, R.; Raza, M.A.S.; Valipour, M.; Saleem, M.F.; Zaheer, M.S.; Ahmad, S.; Toleikiene, M.; Haider, I.; Aslam, M.U.; Nazar, M.A. Potential Agricultural and Environmental Benefits of Mulches—A Review. Bull. Natl. Res. Cent. 2020, 44, 75. [Google Scholar] [CrossRef]
- Liu, C.A.; Jin, S.L.; Zhou, L.M.; Jia, Y.; Li, F.M.; Xiong, Y.C.; Li, X.G. Effects of Plastic Film Mulch and Tillage on Maize Productivity and Soil Parameters. Eur. J. Agron. 2009, 31, 241–249. [Google Scholar] [CrossRef]
- Sekara, A.; Pokluda, R.; Cozzolino, E.; Del Piano, L.; Cuciniello, A.; Caruso, G. Plant Growth, Yield, and Fruit Quality of Tomato Affected by Biodegradable and Non-Degradable Mulches. Hort. Sci. 2019, 46, 138–145. [Google Scholar] [CrossRef] [Green Version]
- Kader, M.A.; Singha, A.; Begum, M.A.; Jewel, A.; Khan, F.H.; Khan, N.I. Mulching as Water-Saving Technique in Dryland Agriculture: Review Article. Bull. Natl. Res. Cent. 2019, 43, 147. [Google Scholar] [CrossRef]
- Verdú, A.M.C.; Mas, M.T.; Josa, R.; Ginovart, M. The Effect of a Prototype Hydromulch on Soil Water Evaporation under Controlled Laboratory Conditions. J. Hydrol. Hydromech. 2020, 68, 404–410. [Google Scholar] [CrossRef]
- Claramunt, J.; Mas, M.T.; Pardo, G.; Cirujeda, A.; Verdú, A.M.C. Mechanical Characterization of Blends Containing Recycled Paper Pulp and Other Lignocellulosic Materials to Develop Hydromulches for Weed Control. Biosyst. Eng. 2020, 191, 35–47. [Google Scholar] [CrossRef]
- López-Marín, J.; Romero, M.; Gálvez, A.; del Amor, F.M.; Piñero, M.C.; Brotons-Martínez, J.M. The Use of Hydromulching as an Alternative to Plastic Films in an Artichoke (Cynara Cardunculus cv. Symphony) Crop: A Study of the Economic Viability. Sustainability 2021, 13, 5313. [Google Scholar] [CrossRef]
- Albacete, A.; Ghanem, M.E.; Dodd, I.C.; Pérez-Alfocea, F. Principal Component Analysis of Hormone Profiling Data Suggests an Important Role for Cytokinins in Regulating Leaf Growth and Senescence of Salinized Tomato. Plant Signal. Behav. 2010, 5, 45–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pareek, A.; Dhankher, O.P.; Foyer, C.H. Mitigating the Impact of Climate Change on Plant Productivity and Ecosystem Sustainability. J. Exp. Bot. 2020, 71, 451–456. [Google Scholar] [CrossRef]
- Raklami, A.; Bechtaoui, N.; Tahiri, A.I.; Anli, M.; Meddich, A.; Oufdou, K. Use of Rhizobacteria and Mycorrhizae Consortium in the Open Field as a Strategy for Improving Crop Nutrition, Productivity and Soil Fertility. Front. Microbiol. 2019, 10, 1106. [Google Scholar] [CrossRef] [Green Version]
- Song, D.; Li, M.; Wang, Q. Evaluation of Models of the Photosynthetic Light Response of Spring Wheat under Different Mulching Treatments in a Desert Oasis in Northwestern China. Crop Sci. 2020, 61, 2036–2047. [Google Scholar] [CrossRef]
- Kader, M.A.; Senge, M.; Mojid, M.A.; Ito, K. Recent Advances in Mulching Materials and Methods for Modifying Soil Environment. Soil Tillage Res. 2017, 168, 155–166. [Google Scholar] [CrossRef]
- Joner, E.J.; Jakobsen, I. Growth and Extracellular Phosphatase Activity of Arbuscular Mycorrhizal Hyphae as Influenced by Soil Organic Matter. Soil Biol. Biochem. 1995, 27, 1153–1159. [Google Scholar] [CrossRef]
- Guo, Y.; Ni, Y.; Hu, J. Effects of Rhizobium, Arbuscular Mycorrhiza and Lime on Nodulation, Growth and Nutrient Uptake of Lucerne in Acid Purplish Soil in China. Trop. Grassl. 2010, 44, 109–114. [Google Scholar]
- Garcés-Ruiz, M.; Calonne-Salmon, M.; Plouznikoff, K.; Misson, C.; Navarrete-Mier, M.; Cranenbrouck, S.; Declerck, S. Dynamics of Short-Term Phosphorus Uptake by Intact Mycorrhizal and Non-Mycorrhizal Maize Plants Grown in a Circulatory Semi-Hydroponic Cultivation System. Front. Plant. Sci. 2017, 8, 147. [Google Scholar] [CrossRef]
- Zhang, J.; Bi, Y.; Song, Z.; Xiao, L.; Christie, P. Arbuscular Mycorrhizal Fungi Alter Root and Foliar Responses to Fissure-Induced Root Damage Stress. Ecol. Indic. 2021, 127, 107800. [Google Scholar] [CrossRef]
- Wu, Q.-S.; Liu, C.-Y.; Zhang, D.-J.; Zou, Y.-N.; He, X.-H.; Wu, Q.-H. Mycorrhiza Alters the Profile of Root Hairs in Trifoliate Orange. Mycorrhiza 2016, 26, 237–247. [Google Scholar] [CrossRef]
- Fausett, J.B.; Rom, C.R. The Effects of Transitioning a Mature High-Density Orchard from Standard Herbicide Ground-Cover Management System to Organic Ground-Cover Management Systems. Res. Ser. Ark. Agric. Exp. Stn. 2001, 483, 33–36. [Google Scholar]
- Zhu, X.; Song, F.; Liu, S.; Liu, F. Arbuscular Mycorrhiza Improve Growth, Nitrogen Uptake, and Nitrogen Use Efficiency in Wheat Grown under Elevated CO2. Mycorrhiza 2016, 26, 133–140. [Google Scholar]
- Liu, H.; Song, F.; Liu, S.; Li, X.; Liu, F.; Zhu, X. Arbuscular Mycorrhiza Improves Nitrogen Use Efficiency in Soybean Grown under Partial Root-Zone Drying Irrigation. Arch. Agron. Soil Sci. 2019, 65, 269–279. [Google Scholar] [CrossRef]
- Sheng, M.; Tang, M.; Chen, H.; Yang, B.; Zhang, F.; Huang, Y. Influence of Arbuscular Mycorrhizae on Photosynthesis and Water Status of Maize Plants under Salt Stress. Mycorrhiza 2008, 18, 287–296. [Google Scholar] [CrossRef]
- Ruiz-Lozano, J.M. Arbuscular Mycorrhizal Symbiosis and Alleviation of Osmotic Stress. New Perspectives for Molecular Studies. Mycorrhiza 2003, 13, 309–317. [Google Scholar] [CrossRef]
- López-Ráez, J.; Verhage, A.; Fernández, I.; García, J.M.; Azcón-Aguilar, C.; Flors, V.; Pozo, M. Hormonal and Transcriptional Profiles Highlight Common and Differential Host Responses to Arbuscular Mycorrhizal Fungi and the Regulation of the Oxylipin Pathway. J. Exp. Bot. 2010, 61, 2589–2601. [Google Scholar] [CrossRef] [Green Version]
- Niu, L.; Yan, Y.; Hou, P.; Bai, W.; Zhao, R.; Wang, Y.; Li, S.; Du, T.; Zhao, M.; Song, J.; et al. Influence of Plastic Film Mulching and Planting Density on Yield, Leaf Anatomy, and Root Characteristics of Maize on the Loess Plateau. Crop J. 2020, 8, 548–564. [Google Scholar] [CrossRef]
- Jahromi, F.; Aroca, R.; Porcel, R.; Ruiz-Lozano, J.M. Microbial Ecology Influence of Salinity on the In Vitro Development of Glomus Intraradices and on the In Vivo Physiological and Molecular Responses of Mycorrhizal Lettuce Plants. Microb. Ecol. 2008, 55, 45–53. [Google Scholar] [CrossRef]
- Wilkinson, S.; Kudoyarova, G.R.; Veselov, D.S.; Arkhipova, T.N.; Davies, W.J. Plant Hormone Interactions: Innovative Targets for Crop Breeding and Management. J. Exp. Bot. 2012, 63, 3499–3509. [Google Scholar] [PubMed]
- Drüge, U.; Schonbeck, F. Effect of Vesicular-Arbuscular Mycorrhizal Infection on Transpiration, Photosynthesis and Growth of Flax (Linum Usitatissimum L.) in Relation to Cytokinin Levels. J. Plant Physiol. 1993, 141, 40–48. [Google Scholar]
- Kohler, J.; Hernández, J.A.; Caravaca, F.; Roldán, A. Plant-Growth-Promoting Rhizobacteria and Arbuscular Mycorrhizal Fungi Modify Alleviation Biochemical Mechanisms in Water-Stressed Plants. Funct. Plant Biol. 2008, 35, 141–151. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The Water-Culture Method for Growing Plants without Soil. In California Agricultural Experiment Station; College of Agriculture University of California: Berkley, CA, USA, 1950; p. 347. [Google Scholar]
- Walker, C. A Simple Blue Staining Technique for Arbuscular Mycorrhizal and Other Root-Inhabiting Fungi. Inoculum 2005, 56, 68–69. [Google Scholar]
- Giovannetti, M.; Mosse, B. An Evaluation of Techniques for Measuring Vesicular Arbuscular Mycorrhizal Infection in Roots. New Phytol. 1980, 84, 489–500. [Google Scholar] [CrossRef]
- Nagata, M.; Yamashita, I. Simple Method for Simultaneous Determination of Chlorophyll and Carotenoids in Tomato Fruit. NSKGA 1992, 39, 925–928. [Google Scholar] [CrossRef] [Green Version]
- Maxwell, K.; Johnson, G.N. Chlorophyll Fluorescence-a Practical Guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar]
- Gerloff, G.C.; Gabelman, W.H. Genetic Basis of Inorganic Plant Nutrition; Läuchli, A., Bieleski, R.L., Eds.; Springer: New York, NY, USA, 1983; Volume 15B. [Google Scholar]
- Baligar, V.C.; Fageria, N.K.; He, Z.L. Nutrient Use Efficiency in Plants. Commun. Soil Sci. Plant Anal. 2001, 32, 921–950. [Google Scholar] [CrossRef]
- Albacete, A.; Martínez-Andújar, C.; Martínez-Pérez, A.; Thompson, A.J.; Dodd, I.C.; Pérez-Alfocea, F. Unravelling Rootstockscion Interactions to Improve Food Security. J. Exp. Bot. 2015, 66, 2211–2226. [Google Scholar] [CrossRef] [Green Version]
- Großkinsky, D.K.; Albacete, A.; Jammer, A.; Krbez, P.; van der Graaff, E.; Pfeifhofer, H.; Roitsch, T. A Rapid Phytohormone and Phytoalexin Screening Method for Physiological Phenotyping. Mol. Plant. 2014, 7, 1053–1056. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AMF | Mulch | NO3− (mg g−1 DW) | P5+ (mg g−1 DW) | K+ (mg g−1 DW) | Mg2+ (mg g−1 DW) | Ca2+ (mg g−1 DW) | SO4−2 (mg g−1 DW) |
| NM | BS | 22.83 ± 4.18 a * | 5.65 ± 0.44 a * | 51.51 ± 1.91 b | 6.06 ± 0.64 a * | 6.44 ± 0.85 a | 7.81 ± 1.83 a |
| PE | 14.22 ± 3.15 a | 5.57 ± 0.40 a * | 54.50 ± 1.22 ab | 5.35 ± 0.80 a | 7.35 ± 0.54 a | 7.56 ± 1.03 a | |
| MS | 12.84 ± 2.10 a* | 5.07 ± 0.29 a * | 58.44 ± 1.87 a | 3.80 ± 0.14 a | 8.33 ± 0.22 a | 7.64 ± 0.95 a | |
| M | BS | 7.60 ± 0.45 a | 3.14 ± 0.32 a | 54.20 ± 2.68 a | 5.89 ± 0.62 a | 6.72 ± 0.93 a | 9.52 ± 1.06 a |
| PE | 7.19 ± 0.92 a | 2.84 ± 0.34 a | 54.54 ± 2.33 a | 4.96 ± 0.34 ab | 6.86 ± 0.33 a | 12.67 ± 1.67 a * | |
| MS | 2.06 ± 0.05 b | 2.71 ± 0.27 a | 53.99 ± 2.90 a | 3.84 ± 0.29 b | 9.18 ± 0.81 a | 7.61 ± 1.32 a | |
| AMF | Mulch | Cu2+ (mg kg−1 DW) | Mn2+ (mg kg−1 DW) | Zn2+ (mg kg−1 DW) | B3+ (mg kg−1 DW) | Na+ (mg g−1 DW) | |
| NM | BS | 6.43 ± 0.96 a | 44.75 ± 50.92 a | 95.34 ± 10.03 a | 37.55 ± 3.03 a | 6.33 ± 0.97 a | |
| PE | 5.47 ± 0.38 a | 34.99 ± 32.19 a | 90.74 ± 9.91 a | 38.42 ± 4.79 a * | 6.67 ± 1.11 a * | ||
| MS | 4.65 ± 0.22 a | 42.06 ± 47.53 a | 78.72 ± 5.49 a | 34.93 ± 5.49 a | 5.23 ± 0.34 a | ||
| M | BS | 7.20 ± 0.72 a * | 41.12 ± 27.92 a | 100.49 ± 10.44 a | 27.48 ± 4.37 a | 4.81 ± S0.83 a | |
| PE | 5.90 ± 0.29 ab | 32.85 ± 25.06 a | 87.25 ± 13.54 a | 32.02 ± 4.44 a | 3.46 ± 0.35 a | ||
| MS | 4.25 ± 0.78 b | 31.49 ± 35.15 a | 64.15 ± 9.41 a | 36.74 ± 3.64 a | 5.12 ± 0.92 a |
| AMF | Mulch | NO3− (mg g−1 DW) | P5+ (mg g−1 DW) | K+ (mg g−1 DW) | Mg2+ (mg g−1 DW) | Ca2+ (mg g−1 DW) | SO4−2 (mg g−1 DW) |
| NM | BS | 5.53 ± 0.56 a | 7.26 ± 0.91 a * | 41.49 ± 5.10 a | 4.76 ± 0.73 a | 6.19 ± 0.71 a | 14.91 ± 1.34 b |
| PE | 5.97 ± 0.77 a | 8.44 ± 0.32 a * | 51.27 ± 1.12 a | 5.28 ± 0.24 a | 6.66 ± 0.32 a | 12.32 ± 0.69 b | |
| MS | 4.08 ± 1.08 a * | 3.92 ± 0.38 b * | 39.58 ± 3.49 a | 4.16 ± 0.48 a | 5.99 ± 0.68 a | 22.24 ± 0.71 a | |
| M | BS | 2.61 ± 0.08 a | 1.93 ± 0.49 a | 70.39 ± 1.52 a | 7.45 ± 1.29 a | 8.54 ± 1.63 a | 18.64 ± 0.80 ab * |
| PE | 2.66 ± 0.23 a | 1.45 ± 0.10 a | 50.76 ± 3.87 a | 5.52 ± 0.22 a | 7.40 ± 0.44 a | 14.11 ± 1.25 b * | |
| MS | 1.22 ± 0.17 b | 2.08 ± 0.36 a | 87.47 ± 1.49 a * | 8.36 ± 1.10 a * | 10.33 ± 1.37 a * | 24.18 ± 2.56 a | |
| AMF | Mulch | Cu2+ (mg kg−1 DW) | Mn2+ (mg kg−1 DW) | Zn2+ (mg kg−1 DW) | B3+ (mg kg−1 DW) | Na+ (mg g−1 DW) | |
| NM | BS | 20.56 ± 1.03 ab | 81.23 ± 5.94 a | 30.03 ± 2.89 a | 13.76 ± 2.26 a | 8.29 ± 0.81 ab | |
| PE | 22.72 ± 8.83 a | 68.12 ± 4.95 ab | 38.22 ± 0.70 a | 18.10 ± 0.66 a | 9.12 ± 0.33 a | ||
| MS | 12.65 ± 0.85 b | 59.77 ± 2.93 a | 29.37 ± 3.24 a | 12.35 ± 1.31 a | 6.53 ± 0.62 b | ||
| M | BS | 27.37 ± 6.25 a | 114.85 ± 8.25 a | 34.85 ± 2.67 a | 21.65 ± 2.77 a | 11.19 ± 2.51 a | |
| PE | 35.42 ± 7.85 a | 63.75 ± 5.20 a | 39.15 ± 3.77 a | 20.57 ± 0.57 a | 10.22 ± 0.72 a | ||
| MS | 22.75 ± 2.77 a * | 187.35 ± 10.11 a * | 45.85 ± 1.35 b | 25.47 ± 1.81 a * | 11.07 ± 1.68 a * |
| AMF | Mulch | NUE | PUE | KUE |
|---|---|---|---|---|
| NM | BS | 25.03 ± 2.58 b | 802.94 ± 93.98 b | 108.74 ± 6.90 a |
| PE | 39.02 ± 6.32 ab | 717.66 ± 29.99 b | 94.66 ± 1.88 a | |
| MS | 59.28 ± 7.97 a | 1127.48 ± 75.91 a | 102.22 ± 2.56 a * | |
| M | BS | 90.88 ± 3.73 b * | 1985.08 ± 82.96 a * | 83.02 ± 8.21 a |
| PE | 91.08 ± 15.97 b * | 2346.22 ± 105.89 a * | 95.19 ± 2.74 a | |
| MS | 335.93 ± 26.96 a * | 1290.88 ± 280.68 a * | 74.01 ± 9.04 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Romero-Muñoz, M.; Gálvez, A.; Martínez-Melgarejo, P.A.; Piñero, M.C.; del Amor, F.M.; Albacete, A.; López-Marín, J. The Interaction between Hydromulching and Arbuscular Mycorrhiza Improves Escarole Growth and Productivity by Regulating Nutrient Uptake and Hormonal Balance. Plants 2022, 11, 2795. https://doi.org/10.3390/plants11202795
Romero-Muñoz M, Gálvez A, Martínez-Melgarejo PA, Piñero MC, del Amor FM, Albacete A, López-Marín J. The Interaction between Hydromulching and Arbuscular Mycorrhiza Improves Escarole Growth and Productivity by Regulating Nutrient Uptake and Hormonal Balance. Plants. 2022; 11(20):2795. https://doi.org/10.3390/plants11202795
Chicago/Turabian StyleRomero-Muñoz, Miriam, Amparo Gálvez, Purificación A. Martínez-Melgarejo, María Carmen Piñero, Francisco M. del Amor, Alfonso Albacete, and Josefa López-Marín. 2022. "The Interaction between Hydromulching and Arbuscular Mycorrhiza Improves Escarole Growth and Productivity by Regulating Nutrient Uptake and Hormonal Balance" Plants 11, no. 20: 2795. https://doi.org/10.3390/plants11202795