
Biochemical Characterization, Antifungal Activity, and Relative Gene Expression of Two Mentha Essential Oils Controlling Fusarium oxysporum, the Causal Agent of Lycopersicon esculentum Root Rot

 ,
,  ,
, 
Abstract
:1. Introduction
2. Results
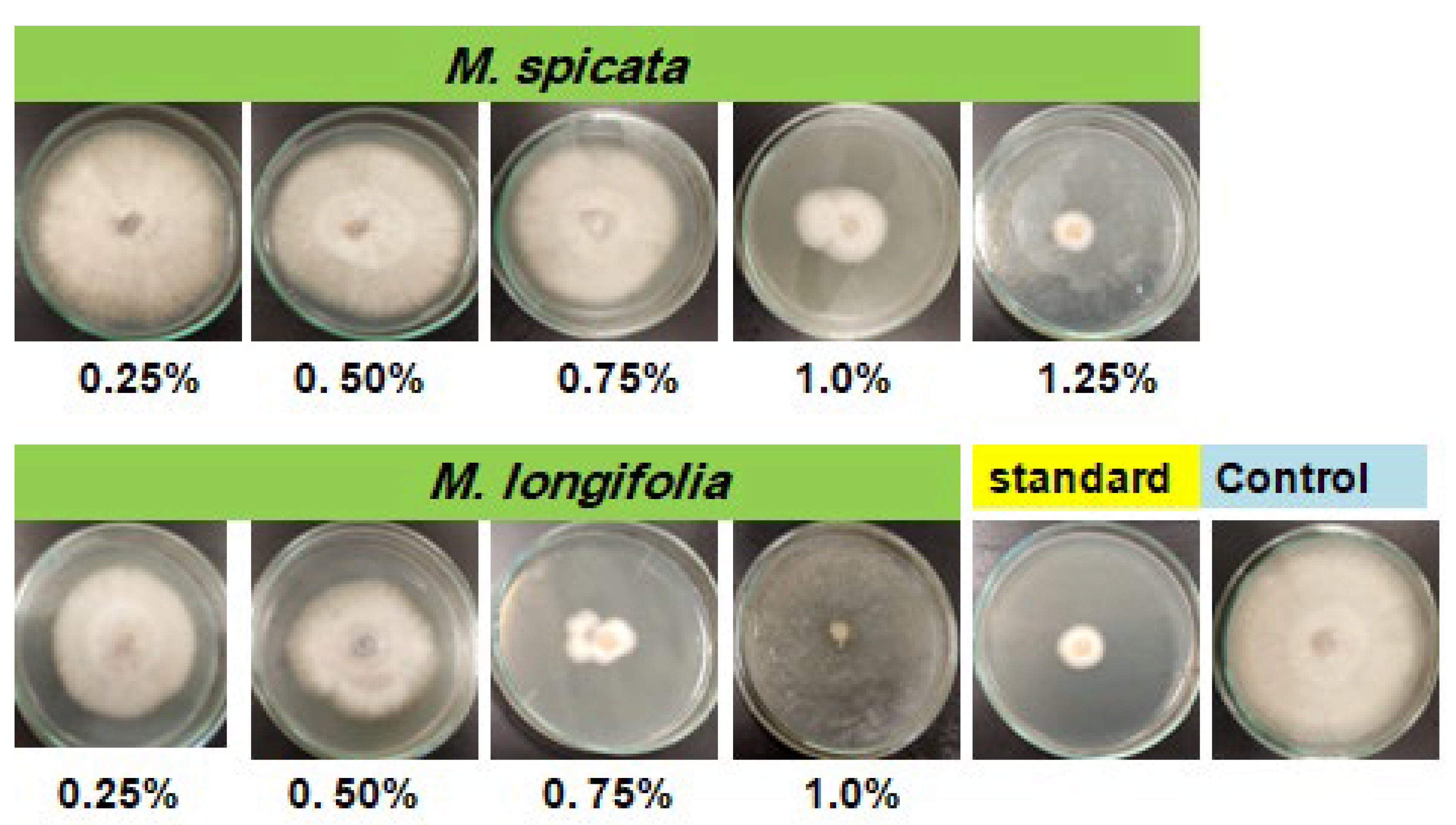
2.1. Screening of Antifungal Activity of Studied EOs
2.2. Chemical Composition of Mentha EOs
2.3. Effects of EOs on Tomato Growth Parameters
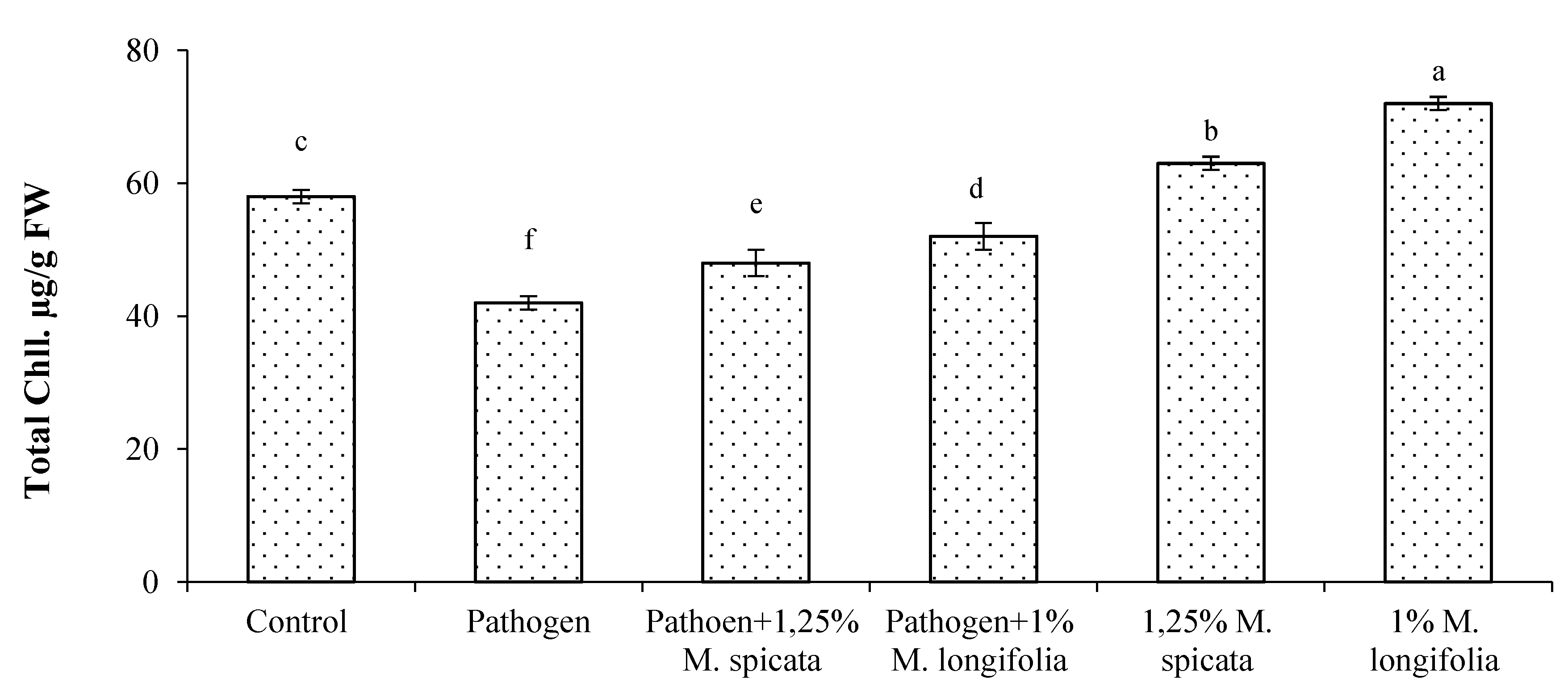
2.4. Protein, Total Phenols, Flavonoids, Malondialdehyde Contents, and Antioxidant Enzymes
2.5. qRT-PCR of the Plant Defence System
3. Discussion
4. Materials and Methods
4.1. Sample Collection, Identification, and Preparation
4.1.1. Fungal Isolate and Tomato Variety
4.1.2. Medicinal Plant Materials
4.2. Extraction of Essential Oils
4.3. GC–MS of Essential Oils
4.4. Preparation of EOs
4.5. In Vitro Antifungal Activity of EOs
4.6. Preparation of Fungal Suspension
4.7. Greenhouse Experiments
4.7.1. Disease Assessments
4.7.2. Analysis of Plant Growth Parameters
4.8. Electrolytes Leakage
4.9. Determination of Total Phenolic and Flavonoid Contents
4.10. Assay of Antioxidant Enzymes
4.11. Gene Expression
4.12. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gupta, S.; Rashotte, A.M. Expression patterns and regulation of SlCRF3 and SlCRF5 in response to cytokinin and abiotic stresses in tomato (Solanum lycopersicum). J. Plant Physiol. 2014, 171, 349–358. [Google Scholar] [CrossRef] [PubMed]
- Bergougnoux, V. The history of tomato: From domestication to biopharming. Biotechnol. Adv. 2014, 32, 170–189. [Google Scholar] [CrossRef] [PubMed]
- Panthee, D.R.; Chen, F. Genomics of fungal disease resistance in tomato. Curr. Genom. 2010, 11, 30–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rongai, D.; Pulcini, P.; Pesce, B.; Milano, F. Antifungal activity of pomegranate peel extract against fusarium wilt of tomato. Eur. J. Plant Pathol. 2017, 147, 229–238. [Google Scholar] [CrossRef]
- Asha, B.B.; Nayaka, C.S.; Shankar, U.A.; Srinivas, C.; Niranjana, S.R. Biological control of F. oxysporum f. sp. lycopersici causing wilt of tomato by Pseudomonas fluorescens. Int. J. Microbiol. Res. 2011, 3, 79–84. [Google Scholar]
- Pandey, K.K.; Gupta, R.C. Pathogenic and cultural variability among Indian isolates of Fusarium oxysporum f. sp. lycopersici causing wilt in tomato. Indian Phytopathol. 2014, 67, 383–387. [Google Scholar]
- Kirankumar, R.; Jagadeesh, K.S.; Krishnaraj, P.U.; Patil, M.S. Enhanced growth promotion of tomato and nutrient uptake by plant growth promoting rhizobacterial isolates in presence of tobacco mosaic virus pathogen. Karnataka J. Agric. Sci. 2010, 21, 309–311. [Google Scholar]
- Singh, V.K.; Singh, H.B.; Upadhyay, R.S. Role of fusaric acid in the development of ‘Fusarium wilt’ symptoms in tomato: Physiological, biochemical and proteomic perspectives. Plant Physiol. Biochem. 2017, 118, 320–332. [Google Scholar] [CrossRef]
- Bhat, R.; Rai, R.V.; Karim, A.A. Mycotoxins in food and feed: Present status and future concerns. Compr. Rev. Food Sci. Saf. 2010, 9, 57–81. [Google Scholar] [CrossRef]
- Gulluce, M. Antimicrobial and antioxidant properties of the essential oils and methanol extract from Mentha longifolia L. ssp. Longifolia. Food Chem. 2007, 103, 1449–1456. [Google Scholar] [CrossRef]
- Moghaddam, M.; Pourbaige, M.; Tabar, H.K.; Farhadi, N.; Hosseini, S.M.A. Composition and antifungal activity of peppermint (Mentha piperita) essential oil from Iran. J. Essent. Oil Bear. Plants 2013, 16, 506–512. [Google Scholar] [CrossRef]
- Hussain, A.I.; Anwar, F.; Nigam, P.S.; Ashraf, M.; Gilani, A.H. Seasonal variation in content, chemical composition and antimicrobial and cytotoxic activities of essential oils from four Mentha species. J. Sci. Food Agric. 2010, 90, 1827–1836. [Google Scholar] [CrossRef]
- Della Pepa, T.; Elshafie, H.S.; Capasso, R.; De Feo, V.; Camele, I.; Nazzaro, F.; Scognamiglio, M.R.; Caputo, L. Antimicrobial and phytotoxic activity of Origanum heracleoticum and O. majorana essential oils growing in cilento (Southern Italy). Molecules 2019, 24, 2576. [Google Scholar] [CrossRef] [Green Version]
- Sonam, K.A.; Kumar, V.; Guleria, I.; Sharma, M.; Kumar, A.; Alruways, M.W.; Khan, N.; Raina, R. Antimicrobial Potential and Chemical Profiling of Leaves Essential Oil of Mentha Species Growing under North-West Himalaya Conditions. J. Pure Appl. Microbiol. 2021, 15, 2229–2243. [Google Scholar] [CrossRef]
- Rizwana, H.; Alwhibi, M.S. Biosynthesis of silver nanoparticles using leaves of Mentha pulegium, their characterization, and antifungal properties. Green Process Synth. 2021, 10, 824–834. [Google Scholar] [CrossRef]
- Hajlaoui, H.; Trabels, N.; Noumi, E. Biological activities of the essential oils and methanol extract of two cultivated mint species (Mentha logifolia and Mentha pulegium) used in the Tunisian folkloric medicine. World J. Microbiol. Biotechnol. 2009, 25, 2227–2238. [Google Scholar] [CrossRef]
- Singh, Y.R.; Sandeep, K.; Anupam, D. Antifungal properties of essential oil Mentha spicata l. var. MSS-5. Indian J. Crop Sci. 2006, 1, 197–200. [Google Scholar]
- Kadoglidou, K.; Chatzopoulou, P.; Maloupa, E.; Kalaitzidis, A.; Ghoghoberidze, S.; Katsantonis, D. Mentha and Oregano Soil Amendment Induces Enhancement of Tomato Tolerance against Soilborne Diseases, Yield and Quality. Agronomy 2020, 10, 23. [Google Scholar] [CrossRef] [Green Version]
- Brahmi, F.; Adjaoud, A.; Marongiu, B.; Porcedda, S.; Piras, A.; Falconieri, D.; Yalaoui-Guellal, D.; Elsebai, M.F.; Madani, K.; Chibane, M. Chemical composition and in vitro antimicrobial, insecticidal and antioxidant activities of the essential oils of Mentha pulegium L. and Mentha rotundifolia (L.) Huds growing in Algeria. Ind. Crops Prod. 2016, 88, 96–105. [Google Scholar] [CrossRef]
- Silva, C.L.; Câmara, J.S. Profiling of volatiles in the leaves of Lamiaceae species based on headspace solid phase microextraction and mass spectrometry. Food Res. Int. 2013, 51, 378–387. [Google Scholar] [CrossRef]
- Bayan, Y.; Aksit, H. Antifungal activity of volatile oils and plant extracts from Sideritis germanicopolitana BORNM growing in Turkey. Egypt J. Biol. Pest Control 2016, 26, 333–337. [Google Scholar]
- Kumar, P.; Mishra, S.; Malik, A.; Satya, S. Insecticidal properties of Mentha species: A review. Ind. Crops Prod. 2011, 34, 802–817. [Google Scholar] [CrossRef]
- Al-Harbi, N.A.; Al Attar, N.M.; Hikal, D.M.; Mohamed, S.E.; Abdel Latef, A.A.H.; Ibrahim, A.A.; Abdein, M.A. Evaluation of insecticidal effects of plants essential oils extracted from basil, black seeds and lavender against Sitophilus oryzae. Plants 2021, 10, 829. [Google Scholar] [CrossRef]
- Sharma, A.; Rajendran, S.; Srivastava, A.; Sharma, S.; Kundu, B. Antifungal activities of selected essential oils against Fusarium oxysporum f. sp. lycopersici 1322, with emphasis on Syzygium aromaticum essential oil. J. Biosci. Bioeng. 2017, 123, 308–313. [Google Scholar] [CrossRef]
- Ben-Jabeur, M.; Ghabri, E.; Myriam, M.; Hamada, W. Thyme essential oil as a defense inducer of tomato against gray mold and Fusarium wilt. Plant Physiol. Biochem. 2015, 94, 35–40. [Google Scholar] [CrossRef]
- De Vos, M.; Van Oosten, V.R.; Van Poecke, R.M.; Van Pelt, J.A.; Pozo, M.J.; Mueller, M.J.; Pieterse, C.M. Signal signature and transcriptome changes of Arabidopsis during pathogen and insect attack. Mol. Plant Microbe Interact. 2005, 18, 923–937. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Liu, W.; Qi, X.; Liu, Z.; Xie, W.; Wang, Y. Genome-wide identification, expression profiling, and SSR marker development of the bZIP transcription factor family in Medicago truncatula. Biochem. Syst. Ecol. 2015, 61, 218–228. [Google Scholar] [CrossRef]
- Nan, H.; Li, W.; Lin, Y.; Gao, L. Genome-Wide Analysis of WRKY genes and their response to salt stress in the wild progenitor of Asian cultivated rice, Oryza rufipogon. Front. Genet. 2020, 11, 359. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, L. The WRKY transcription factor superfamily: Its origin in eukaryotes and expansion in plants. BMC Evol. Biol. 2005, 5, 1. [Google Scholar] [CrossRef] [Green Version]
- Wen, F.; Zhu, H.; Li, P.; Jiang, M.; Mao, W.; Ong, C.; Chu, Z. Genome-wide evolutionary characterization and expression analyses of WRKY family genes in Brachypodium distachyon. DNA Res. 2014, 21, 327–339. [Google Scholar] [CrossRef] [Green Version]
- Guo, C.; Guo, R.; Xu, X.; Gao, M.; Li, X.; Song, J.; Zheng, Y.; Wang, X. Evolution and expression analysis of the grape (Vitis vinifera L.) WRKY gene family. J. Exp. Bot. 2014, 65, 1513–1528. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Zhang, S. MAPK cascades in plant disease resistance signaling. Ann. Rev. Phytopathol. 2013, 51, 245–266. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Qamar, S.A.; Chen, Z.; Mengiste, T. Arabidopsis WRKY33 transcription factor is required for resistance to necrotrophic fungal pathogens. Plant J. 2006, 48, 592–605. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Meng, X.; Wang, R.; Mao, G.; Han, L.; Liu, Y.; Zhang, S. Dual-level regulation of ACC synthase activity by MPK3/MPK6 cascade and its downstream WRKY transcription factor during ethylene induction in Arabidopsis. PLoS Genet. 2012, 8, 1002767. [Google Scholar] [CrossRef]
- Szczechura, W.; Staniaszek, M.; Habdas, H. Fusarium oxysporum F. sp. Radicis lycopersici—The cause of fusarium crown and root rot in tomato cultivation. J. Plant Protec. Res. 2013, 53, 172–176. [Google Scholar] [CrossRef]
- Ozbay, N.; Newman, S.E. Fusarium crown and root rot of tomato and control methods. Plant Pathol. J. 2004, 3, 9–18. [Google Scholar] [CrossRef] [Green Version]
- Cosentino, C.; Labella, C.; Elshafie, H.S.; Camele, I.; Musto, M.; Paolino, R.; Freschi, P. Effects of different heat treatments on lysozyme quantity and antimicrobial activity of jenny milk. J. Dairy Sci. 2016, 99, 5173–5179. [Google Scholar] [CrossRef] [Green Version]
- Elshafie, H.S.; Viggiani, L.; Mostafa, M.S.; El-Hashash, M.A.; Bufo, S.A.; Camele, I. Biological activity and chemical identification of ornithine lipid produced by Burkholderia gladioli pv. agaricicola ICMP 11096 using LC-MS and NMR analyses. J. Biol. Res. 2017, 90, 96–103. [Google Scholar] [CrossRef]
- Ravensberg, W. Crop protection in 2030: Towards a natural, efficient, safe and sustainable approach. In Proceedings of the IBMA International Symposium, Swansea University, Swansea, UK, 7–9 September 2015. [Google Scholar]
- Camele, I.; Elshafie, H.S.; Caputo, L.; Sakr, S.H.; De Feo, V. Bacillus mojavensis: Biofilm formation and biochemical investigation of its bioactive metabolites. J. Biol. Res. 2019, 92, 39–45. [Google Scholar] [CrossRef]
- Elshafie, H.S.; Devescovi, G.; Venturi, V.; Camele, I.; Bufo, S.A. Study of the regulatory role of N-acyl homoserine lactones mediated quorum sensing in the biological activity of Burkholderia gladioli pv. agaricicola causing soft rot of Agaricus spp. Front. Microbiol. 2019, 10, 2695. [Google Scholar] [CrossRef]
- Sofo, A.; Elshafie, H.S.; Scopa, A.; Mang, S.M.; Camele, I. Impact of airborne zinc pollution on the antimicrobial activity of olive oil and the microbial metabolic profiles of Zn-contaminated soils in an Italian olive orchard. J. Trace Elem. Med. Biol. 2018, 49, 276–284. [Google Scholar] [CrossRef]
- Elshafie, H.S.; Racioppi, R.; Bufo, S.A.; Camele, I. In vitro study of biological activity of four strains of Burkholderia gladioli pv. agaricicola and identification of their bioactive metabolites using GC–MS. Saudi J. Biol. Sci. 2017, 24, 295–301. [Google Scholar] [CrossRef] [Green Version]
- Raveau, R.; Fontaine, J.; Lounès-Hadj Sahraoui, A. Essential oils as potential alternative biocontrol products against plant pathogens and weeds: A Review. Foods 2020, 9, 365. [Google Scholar] [CrossRef] [Green Version]
- Elshafie, H.S.; Caputo, L.; De Martino, L.; Gruľová, D.; Zheljazkov, V.D.; De Feo, V.; Camele, I. Biological investigations of essential oils extracted from three Juniperus species and evaluation of their antimicrobial, antioxidant and cytotoxic activities. J. Appl. Microbiol. 2020, 129, 1261–1271. [Google Scholar] [CrossRef]
- Camele, I.; Grul’ová, D.; Elshafie, H.S. Chemical composition and antimicrobial properties of Mentha piperita cv. ‘Kristinka’ essential oil. Plants 2021, 10, 1567. [Google Scholar] [CrossRef]
- Ali, H.M.; Elgat, W.; El-Hefny, M.; Salem, M.Z.M.; Taha, A.S.; Al Farraj, D.A.; Elshikh, M.S.; Hatamleh, A.A.; Abdel-Salam, E.M. New Approach for Using of Mentha longifolia L. and Citrus reticulata L. Essential Oils as Wood-Biofungicides: GC-MS, SEM, and MNDO Quantum Chemical Studies. Materials 2021, 14, 18. [Google Scholar] [CrossRef]
- Ragab, M.M.M.; Ashour, A.M.A.; Abdel-Kader, M.M.; ElMohamady, R.; Abdel-Aziz, A. In vitro evaluation of some fungicides alternatives against Fusarium oxysporum the causal of wilt disease of pepper (Capsicum annum L.). Int. J. Agric. For. 2012, 2, 70–77. [Google Scholar]
- Arnal-Schnebelen, B.; Hadji-Minaglou, F.; Peroteau, J.F.; Ribeyre, F.; de Billerbeck, V.G. Essential oils in infectious gynaecological disease: A statistical study of 658 cases. Int. J. Aromatherapy. 2004, 14, 192–197. [Google Scholar] [CrossRef]
- Sharma, N.; Tripathi, A. Effects of Citrus sinensis (L.) Osbeck epicarp essential oil on growth and morphogenesis of Aspergillus niger Van Tieghem. Microbiol. Res. 2008, 163, 337–344. [Google Scholar] [CrossRef]
- Camele, I.; Elshafie, H.S.; De Feo, V.; Caputo, L. Anti-quorum sensing and antimicrobial effect of Mediterranean plant essential oils against phytopathogenic bacteria. Front. Microbiol. 2019, 10, 2619. [Google Scholar] [CrossRef]
- Regmi, S.; Jha, S.K. Antifungal activity of plant essential oils against Fusarium oxysporum schlecht. and Aspergillus niger van tiegh. from papaya. Inter. J. Curr. Trends Sci. Tech. 2017, 8, 20196–20204. [Google Scholar]
- Gruľová, D.; Caputo, L.; Elshafie, H.S.; Baranová, B.; De Martino, L.; Sedlák, V.; Camele, I.; De Feo, V. Thymol chemotype Origanum vulgare L. essential oil as a potential selective bio-based herbicide on monocot plant species. Molecules 2020, 25, 595. [Google Scholar] [CrossRef] [Green Version]
- Yakhlef, G.; Hambaba, L.; Pinto, D.; Silva, A.M.S. Chemical composition and insecticidal, repellent and antifungal activities of essential oil of Mentha rotundifolia (L.) from Algeria. Ind. Crops Prod. 2020, 158, 8. [Google Scholar] [CrossRef]
- Krishna Kishore, G.; Pande, S.; Harish, S. Evaluation of essential oils and their components for broad-spectrum antifungal activity and control of late leaf spot and crown rot diseases in peanut. Plant Dis. 2007, 91, 375–379. [Google Scholar] [CrossRef] [Green Version]
- Oussalah, M.; Caillet, S.; Saucier, L.; Lacroix, M. Antimicrobial effects of selected plant essential oils on the growth of a Pseudomonas putida strain isolated from meat. Meat Sci. 2006, 73, 236–244. [Google Scholar] [CrossRef]
- Chen, C.J.; Li, Q.Q.; Zeng, Z.Y.; Duan, S.S.; Wang, W.; Xu, F.R.; Cheng, Y.X.; Dong, X. Efficacy and mechanism of Mentha haplocalyx and Schizonepeta tenuifolia essential oils on the inhibition of Panax notoginseng pathogens. Ind. Crops Prod. 2020, 145, 12. [Google Scholar] [CrossRef]
- Elshafie, H.S.; Ghanney, N.; Mang, S.M.; Ferchichi, A.; Camele, I. An in vitro attempt for controlling severe phytopathogens and human pathogens using essential oils from Mediterranean plants of genus Schinus. J. Med. Food 2016, 19, 266–273. [Google Scholar] [CrossRef]
- Elshafie, H.S.; Sakr, S.; Mang, S.M.; De Feo, V.; Camele, I. Antimicrobial activity and chemical composition of three essential oils extracted from Mediterranean aromatic plants. J. Med. Food. 2016, 19, 1096–1103. [Google Scholar] [CrossRef] [PubMed]
- Burt, S. Essential oils: Their antibacterial properties and potential applications in foods a review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar] [CrossRef] [PubMed]
- Bayan, Y.; Küsek, M. Chemical composition and antifungal and antibacterial activity of Mentha spicata L. volatile oil. Cienc. Investig. Agrar. 2018, 45, 64–69. [Google Scholar] [CrossRef] [Green Version]
- Desam, N.R.; Al-Rajab, A.J.; Sharma, M.; Mylabathula, M.M.; Gowkanapalli, R.R.; Albratty, M. Chemical composition, antibacterial and antifungal activities of Saudi Arabian Mentha longifolia L. essential oil. J. Coast. Life Med. 2017, 5, 441–446. [Google Scholar] [CrossRef]
- Kumar, P.; Mishra, S.; Malik, A.; Satya, S. Compositional analysis and insecticidal activity of Eucalyptus globulus (family: Myrtaceae) essential oil against housefly (Musca domestica). Acta Trop. 2012, 122, 212–218. [Google Scholar] [CrossRef]
- Devi, K.P.; Nisha, S.A.; Sakthivel, R.; Pandian, S.K. Eugenol (an essential oil of clove) acts as an antibacterial agent against Salmonella typhi by disrupting the cellular membrane. J. Ethnopharmacol. 2010, 130, 107–115. [Google Scholar] [CrossRef]
- Phukan, U.J.; Jeena, G.S.; Shukla, R.K. WRKY transcription factors: Molecular regulation and stress responses in plants. Front. Plant Sci. 2016, 7, 760. [Google Scholar] [CrossRef] [Green Version]
- Després, C.; Chubak, C.; Rochon, A.; Clark, R.; Bethune, T.; Desveaux, D.; Fobert, P. The Arabidopsis NPR1 disease resistance protein is a novel cofactor that confers redox regulation of DNA binding activity to the basic domain/leucine zipper transcription factor TGA1. Plant Cell 2003, 15, 2181–2191. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Amornsiripanitch, N.; Dong, X. A genomic approach to identify regulatory nodes in the transcriptional network of systemic acquired resistance in plants. PLoS Pathog. 2006, 2, 1024–1050. [Google Scholar] [CrossRef] [Green Version]
- Koo, S.C.; Moon, B.C.; Kim, J.K.; Kim, C.Y.; Sung, S.J.; Kim, M.C.; Cho, M.; Cheong, Y. OsBWMK1 mediates SA-dependent defense responses by activating the transcription factor OsWRKY33. Biochem. Biophys. Res. Commun. 2009, 387, 365–370. [Google Scholar] [CrossRef]
- Lai, Z.; Vinod, K.M.; Zheng, Z.; Fan, B.; Chen, Z. Roles of Arabidopsis WRKY3 and WRKY4 transcription factors in plant responses to pathogens. BMC Plant Biol. 2008, 8, 68. [Google Scholar] [CrossRef] [Green Version]
- Dong, J.; Chen, C.; Chen, Z. Expression profile of the Arabidopsis WRKY gene superfamily during plant defense response. Plant Mol. Biol. 2003, 51, 21–37. [Google Scholar] [CrossRef]
- Xu, X.; Chen, C.; Fan, B.; Chen, Z. Physical and functional interactions between pathogen-induced Arabidopsis WRKY18, WRKY40, and WRKY60 transcription factors. Plant Cell 2006, 18, 1310–1326. [Google Scholar] [CrossRef] [Green Version]
- Périno, S.; Chemat-Djenni, Z.; Petitcolas, E.; Giniès, C.; Chemat, F. Downscaling of industrial turbo-distillation to laboratory turbo-clevenger for extraction of essential oils. Application of concepts of green analytical chemistry. Molecules 2019, 24, 2734. [Google Scholar] [CrossRef] [Green Version]
- Elshafie, H.S.; Mancini, E.; Sakr, S.; De Martino, L.; Mattia, C.A.; De Feo, V.; Camele, I. Antifungal activity of some constituents of Origanum vulgare L. essential oil against postharvest disease of peach fruit. J. Med. Food 2015, 18, 929–934. [Google Scholar] [CrossRef]
- Tatsadjieu, N.L.; Dongmo, P.J.; Ngassoum, M.B.; Etoa, F.X.; Mbofung, C.M.F. Investigations on the essential oil of Lippia rugosa from Cameroon for its potential use as antifungal agent against Aspergillus flavus Link ex. Fries. Food Control 2009, 20, 161–166. [Google Scholar] [CrossRef]
- Abd-El-Moity, T.H. Effect of single and mixture of Trichoderma harzianum isolates on controlling three different soil borne pathogens. Egyptian J. Microbiol. 1985, 111–120. [Google Scholar]
- Filion, M.; St-Arnaud, M.; Jabaji-Hare, S.H. Quantification of Fusarium solani f. sp. phaseoli in mycorrhizal bean plants and surrounding mycorrhizosphere soil using real-time polymerase chain reaction and direct isolations on selective media. Phytopathology 2003, 93, 229–235. [Google Scholar] [CrossRef] [Green Version]
- Lichtenthaler, H.; Wellburn, A. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef] [Green Version]
- Blum, A.; Ebercon, A. Cell membrane stability as a measure of drought and heat tolerance in wheat. Crop Sci. 1981, 21, 43–47. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts. I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophisics 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Slinkard, K.; Singleton, V.L. Total phenol analysis: Automation and comparison with manual methods. Am. J. Enol. Vitic. 1977, 28, 49–55. [Google Scholar]
- Chavan, J.J.; Gaikwad, N.B.; Kshirsagar, P.R.; Dixit, G.B. Total phenolics, flavonoids and antioxidant properties of three Ceropegia species from Western Ghats of India. South Afr. J. Bot. 2013, 88, 273–277. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Beyer, W.F., Jr.; Fridovich, I. Assaying for superoxide dismutase activity: Some large consequences of minor changes in conditions. Anal. Biochem. 1987, 161, 559–566. [Google Scholar] [CrossRef]
- Bergmeyer, H.U. Methods of Enzymatic Analysis, 2nd ed.; Academic Press: New York, NY, USA, 1974; p. 855. [Google Scholar]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Rebouças, E.D.L.; Costa, J.J.N.; Passos, M.J.; Passos, J.R.S.; Hurk, R.; Silva, J.R.V. Real time PCR and importance of housekeepings genes for normalization and quantification of mRNA expression in different tissues. Braz. Arch. Biol. Technol. 2013, 56, 143–154. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-DDCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Medeiros, F.C.L.; Resende, M.L.V.; Medeiros, F.H.V.; Zhang, H.M.; Pare, P.W. Defense gene expression induced by a coffee-leaf extract formulation in tomato. Physiol. Mol. Plant Pathol. 2009, 74, 175–183. [Google Scholar] [CrossRef]
- Hafez, E.E.; Hashem, M.; Balbaa, M.M.; El-Saadani, M.A.; Ahmed, S.A. Induction of new defensin genes in tomato plants via pathogens-biocontrol agent interaction. J. Plant Pathol. Microbiol. 2013, 4, 167. [Google Scholar]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Quantitative ID | Component Identified | Retention Time (min) | Retention Index (RI) | Area (%) | Identification |
|---|---|---|---|---|---|
| 1 | Piperitone | 5.20 | 1245 | 12.34 | RI, MS |
| 2 | IR-alpha-pinene | 7.82 | 933 | 0.76 | RI, MS |
| 3 | D-limonene | 11.83 | 1031 | 0.77 | RI, MS |
| 4 | Cis-P-menthan | 17.63 | 984 | 0.99 | RI, MS |
| 5 | Menthone | 18.13 | 1150 | 0.67 | RI, MS |
| 6 | P-menthan-3-ol alcohol | 18.55 | 1164 | 2.27 | RI, MS |
| 7 | Alpha-terpin | 19.49 | 1187 | 6.11 | RI, MS |
| 8 | Gamma-terpineol | 19.73 | 1185 | 0.82 | RI, MS |
| 9 | β-caryophyllene | 27.19 | 1325 | 18.7 | RI, MS |
| 10 | Butanedioic acid | 32.28 | 1580 | 6.26 | RI, MS |
| 11 | Ethyl 4-heptyl ester | 36.54 | 1378 | 8.83 | RI, MS |
| 12 | Adipic acid | 56.70 | 1507 | 13.24 | RI, MS |
| 13 | Menthol | 66.70 | 1182 | 12.08 | RI, MS |
| 14 | Thymol | 72.10 | 1290 | 14.62 | RI, MS |
| 15 | Others | - | - | 1.54 | RI, MS |
| Quantitative ID | Component Identified | Retention Time (min) | Retention Index (RI) | Area (%) | Identification |
|---|---|---|---|---|---|
| 1 | Alpha piene | 4.05 | 933 | 6.87 | RI, MS |
| 2 | β-pinene | 12.21 | 964 | 0.54 | RI, MS |
| 3 | D-limonene | 12.43 | 1031 | 10.33 | RI, MS |
| 4 | Borneol | 12.72 | 1165 | 0.38 | RI, MS |
| 5 | β-terpinyl acetate | 16.89 | 1267 | 0.47 | RI, MS |
| 6 | Menthol | 24.32 | 1182 | 44.17 | RI, MS |
| 7 | Menthone | 26.48 | 1150 | 0.55 | RI, MS |
| 8 | Menthyl acetate | 29.63 | 1294 | 18.36 | RI, MS |
| 9 | Linalool | 31.74 | 1083 | 0.43 | RI, MS |
| 10 | Eugenol | 43.6 | 1209 | 0.64 | RI, MS |
| 11 | Carvone | 46.89 | 1242 | 0.33 | RI, MS |
| 12 | Thymol | 5.08 | 1290 | 11.23 | RI, MS |
| 13 | Cavacrol | 59.55 | 1298 | 0.36 | RI, MS |
| 14 | Cis-jasmone | 63.41 | 1394 | 0.56 | RI, MS |
| 15 | Cinerolone | 63.52 | 1641 | 0.53 | RI, MS |
| 16 | Caryophyllene | 64.32 | 1418 | 0.74 | RI, MS |
| 17 | β-farnesene | 64.38 | 1452 | 0.35 | RI, MS |
| 18 | β-cubebene | 64.57 | 1389 | 0.36 | RI, MS |
| 19 | Alpha-cadinol | 64.78 | 1627 | 0.82 | RI, MS |
| 20 | Others | - | - | 1.98 | RI, MS |
| EOs Treatment | PH (cm) | SFW (g) | SDW (g) | RFW (g) | RDW (g) | |
|---|---|---|---|---|---|---|
| Control | Control Plants | 24.32 ± 0.02 c | 15.44 ± 0.02 c | 1.98 ± 0.01 c | 1.52 ± 0.01 c | 0.16 ± 0.01 c |
| 1.25% M. spicata | 28.9 ± 0.01 b | 15.96 ± 0.02 b | 2.09 ± 0.01 b | 1.64 ± 0.01b | 0.18 ± 0.01 b | |
| 1% M. longifolia | 32.42 ± 0.02 a | 16.33 ± 0.01 a | 2.34 ± 0.01 a | 1.87 ± 0.01 a | 0.24 ± 0.01 a | |
| Pathogen | Infected Plants | 16.54 ± 0.02 f | 9.52 ± 0.03 f | 0.97 ± 0.01 f | 1.21 ± 0.02 f | 0.1 ± 0.01 f |
| 1.25% M. spicata | 18.76 ± 0.01 e | 12.97 ± 0.01 e | 1.67 ± 0.01 e | 1.34 ± 0.15 e | 0.12 ± 0.01 e | |
| 1% M. longifolia | 19.86 ± 0.02 d | 13.64 ± 0.01 d | 1.85 ± 0.01 d | 1.41 ± 0.02 d | 0.14 ± 0.01 d |
| Gene Name | Sequence | ID & Reference | |
|---|---|---|---|
| Chitinase (PR3) | F | 5′- ATGGCGGAAACTGTCCTAGTGGAA -3′ | Medeiros et al. [88] |
| R | 5′ ACATGGTCTACCATCAGCTTGCCA -3′ | ||
| Defensin (PR12) | F | 5′- TCACCAAACTATTGGATTTCAA -3′ | Hafez et al. [89] |
| R | 5′- GACTCAATTTTTGACTTCTTAATCC -3′ | ||
| WRKY1 | F | 5′- CGCAACTCAAAGAGACGGAAG-3′ | Solyc07g047960.2.1 |
| R | 5′- CATTGACTACATCCACTTCACTGC-3′ | ||
| WRKY4 | F | 5’- CGTTGCACATACCCTGGATG -3′ | Solyc05g012770.2.1 |
| R | 5′- GGCCTCCAAGTTGCAATCTC -3′ | ||
| WRKY33 | R | 5′- CCACCTCCTTCACTTCCATT -3′ | Solyc09g014990.2.1 |
| F | 5′- GATGGAAAACTCCCAGTCGT -3′ | ||
| WRKY53 | F | 5′- CACATACCGAGGCTCCCATAA -3′ | Solyc08g008280.2.1 |
| R | 5′- CCTGTTGGATAAACGGCTTGG -3′ | ||
| β-Actin | F | 5′- TCCTTCTTGGGTATGGAATCCT-3′ | NM_007393.5 |
| R | 5′- CAGCACTGTGTTGGCATAGA-3′ | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soliman, S.A.; Hafez, E.E.; Al-Kolaibe, A.M.G.; Abdel Razik, E.-S.S.; Abd-Ellatif, S.; Ibrahim, A.A.; Kabeil, S.S.A.; Elshafie, H.S. Biochemical Characterization, Antifungal Activity, and Relative Gene Expression of Two Mentha Essential Oils Controlling Fusarium oxysporum, the Causal Agent of Lycopersicon esculentum Root Rot. Plants 2022, 11, 189. https://doi.org/10.3390/plants11020189
Soliman SA, Hafez EE, Al-Kolaibe AMG, Abdel Razik E-SS, Abd-Ellatif S, Ibrahim AA, Kabeil SSA, Elshafie HS. Biochemical Characterization, Antifungal Activity, and Relative Gene Expression of Two Mentha Essential Oils Controlling Fusarium oxysporum, the Causal Agent of Lycopersicon esculentum Root Rot. Plants. 2022; 11(2):189. https://doi.org/10.3390/plants11020189
Chicago/Turabian StyleSoliman, Seham A., Elsayed E. Hafez, Abdu M. G. Al-Kolaibe, El-Sayed S. Abdel Razik, Sawsan Abd-Ellatif, Amira A. Ibrahim, Sanaa S. A. Kabeil, and Hazem S. Elshafie. 2022. "Biochemical Characterization, Antifungal Activity, and Relative Gene Expression of Two Mentha Essential Oils Controlling Fusarium oxysporum, the Causal Agent of Lycopersicon esculentum Root Rot" Plants 11, no. 2: 189. https://doi.org/10.3390/plants11020189