
Can Ethylene Inhibitors Enhance the Success of Olive Somatic Embryogenesis?

 ,
,  , , and
, , and
Abstract
:1. Introduction
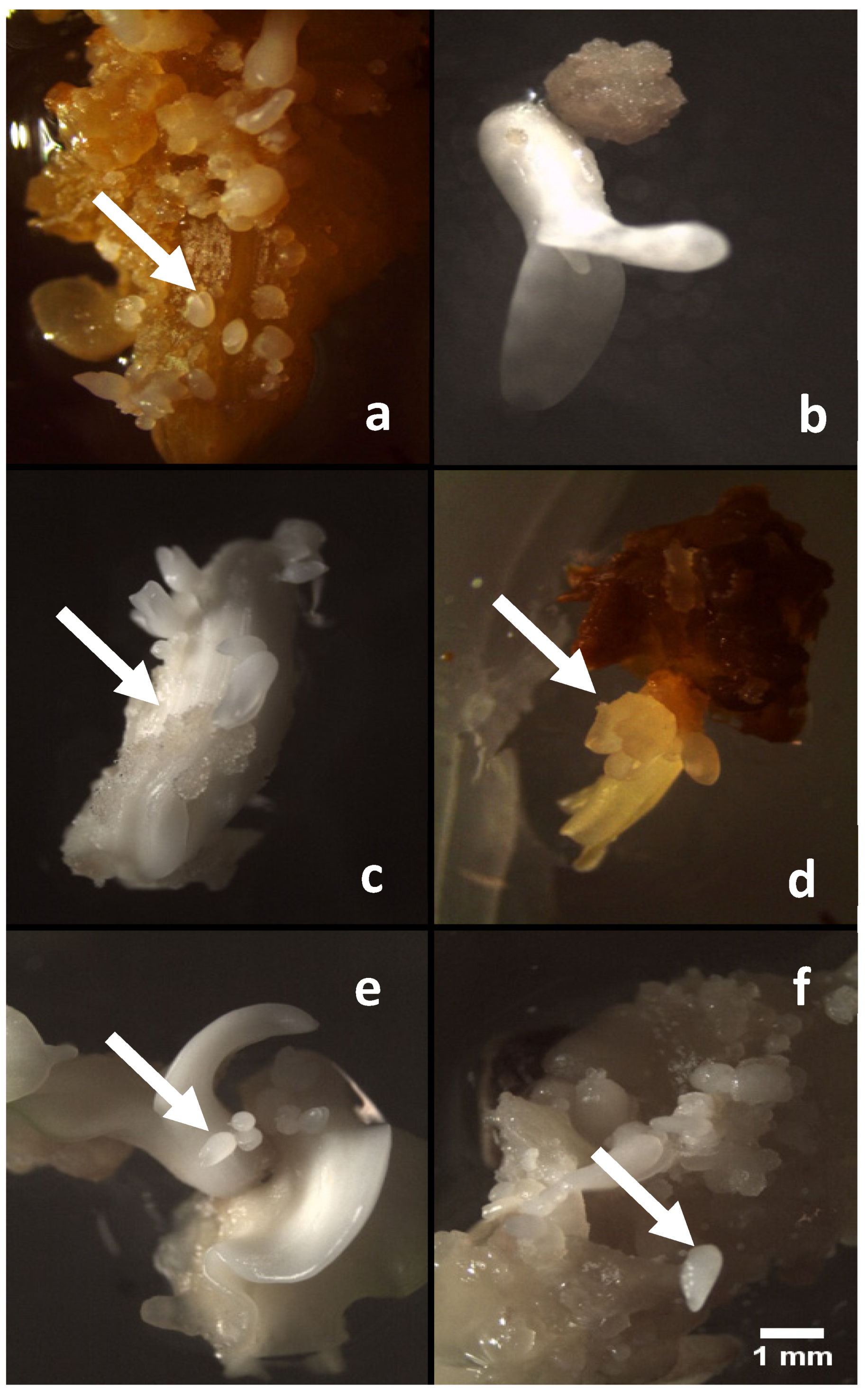
2. Results and Discussion
3. Materials and Methods
3.1. Plant Material
3.2. Somatic Embryogenesis Induction
3.3. Expression Phase and Maturation
3.4. Culture Conditions
3.5. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Rugini, E.; Silvestri, C. Somatic Embryogenesis in Olive (Olea europaea L. subsp. europaea var. sativa and var. sylvestris). In In Vitro Embryogenesis in Higher Plants; Germana, M.A., Lambardi, M., Eds.; Humana Press: New York, NY, USA, 2016; pp. 341–349. [Google Scholar]
- Rugini, E.; Cristofori, V.; Silvestri, C. Genetic improvement of olive (Olea europaea L.) by conventional and in vitro biotechnology methods. Biotechnol. Adv. 2016, 34, 687–696. [Google Scholar] [CrossRef]
- Rugini, E. In vitro propagation of some olive cultivars with different root-ability and medium development using analytical data from developing shoots and embryos. Sci. Hortic. 1984, 24, 123–134. [Google Scholar] [CrossRef]
- Regni, L.; Micheli, M.; Del Pino, A.M.; Palmerini, C.A.; D’Amato, R.; Facchin, S.L.; Famiani, F.; Peruzzi, A.; Mairech, H.; Proietti, P. The first evidence of the beneficial effects of se-supplementation on in vitro cultivated olive tree explants. Plants 2021, 10, 1630. [Google Scholar] [CrossRef]
- Leva, A.R.; Muleo, R.; Petruccelli, R. Long-term somatic embryogenesis from immature olive cotyledons. J. Hortic. Sci. Biotechnol. 1995, 70, 417–421. [Google Scholar] [CrossRef]
- Rugini, E. Somatic embryogenesis and plant regeneration in olive (Olea europaea L.). Plant Cell Tissue Org. Cult. 1988, 14, 207–214. [Google Scholar] [CrossRef]
- Orinos, P.; Mitrakos, K. Rhizogenesis and somatic embryogenesis in calli from wild olive (Olea europaea var sylvestris (Miller) Lehr) mature zygotic embryos. Plant Cell Tissue Organ Cult. 1991, 27, 183–187. [Google Scholar] [CrossRef]
- Mitrakos, K.; Alexaki, A.; Papadimitriou, P. Dependence of olive morphogenesis on callus origin and age. J. Plant Physiol. 1992, 139, 269–273. [Google Scholar] [CrossRef]
- Cerezo, S.; Mercado, J.A.; Pliego-Alfaro, F. An efficient regeneration system via somatic embryogenesis in olive. Plant Cell Tissue Org. Cult. 2011, 106, 337–344. [Google Scholar] [CrossRef] [Green Version]
- Shibli, R.A.; Shatnawi, M.; Abu, E.; Al-Juboory, K.H. Somatic embryogenesis and plant recovery from callus of ‘Nabali’ Olive (Olea europaea L.). Sci. Hortic. 2001, 88, 243–256. [Google Scholar] [CrossRef]
- Rugini, E.; Caricato, G. Somatic embryogenesis and plant recovery from mature tissues of olive cultivars (Olea europaea L.) ‘Canino’ and ‘Moraiolo’. Plant Cell Rep. 1995, 14, 257–260. [Google Scholar] [CrossRef]
- Mazri, M.A.; Belkoura, I.; Pliego-Alfaro, F.; Belkoura, M. Somatic embryogenesis from leaf and petiole explants of the Moroccan olive cultivar dahbia. Sci. Hortic. 2013, 159, 88–95. [Google Scholar] [CrossRef]
- Toufik, I.; Guenoun, F.; Belkoura, I. Embryogenesis expression from somatic explants of olive (Olea europaea L.) cv Picual. Moroccan J. Biol. 2014, 11, 17–25. [Google Scholar]
- Capelo, A.M.; Silva, S.; Brito, G.; Santos, C. Somatic embryogenesis induction in leaves and petioles of a mature wild olive. Plant Cell Tissue Org. Cult. 2010, 103, 237–242. [Google Scholar] [CrossRef]
- Narváez, I.; Martín, C.; Jiménez-Díaz, R.M.; Mercado, J.A.; Pliego-Alfaro, F. Plant regeneration via somatic embryogenesis in mature wild olive genotypes resistant to the defoliating pathotype of verticillium dahliae. Front. Plant Sci. 2019, 10, 1471. [Google Scholar] [CrossRef]
- Sánchez-Romero, C. Somatic embryogenesis in olea spp. Plant Cell Tissue Org Cult. 2019, 138, 403–426. [Google Scholar] [CrossRef]
- Kumar, V.; Ramakrishna, A.; Ravishankar, G.A. Influence of different ethylene inhibitors on somatic embryogenesis and secondary embryogenesis from Coffea canephora P ex Fr. In Vitro Cell. Dev. Biol. Plant 2007, 43, 602–607. [Google Scholar] [CrossRef]
- Schaller, G.E.; Binder, B.M. Inhibitors of ethylene biosynthesis and signaling. In Ethylene Signaling: Methods and Protocols; Binder, B.M., Schaller, G.E., Eds.; Humana Press: New York, NY, USA, 2017; pp. 223–235. [Google Scholar]
- Sgamma, T.; Thomas, B.; Muleo, R. Ethylene inhibitor silver nitrate enhances regeneration and genetic transformation of prunus avium (L.) cv stella. Plant Cell Tissue Org. Cult. 2015, 120, 79–88. [Google Scholar] [CrossRef]
- Fuentes, S.R.; Calheiros, M.B.; Manetti-Filho, J.; Vieira, L.G. The effects of silver nitrate and different carbohydrate sources on somatic embryogenesis in Coffea canephora. Plant Cell Tissue Org. Cult. 2000, 60, 5–13. [Google Scholar] [CrossRef]
- Huang, Y.F.; Chen, C.T.; Kao, C.H. Salicylic acid inhibits the biosynthesis of ethylene in detached rice leaves. Plant Growth Reg. 1993, 12, 79–82. [Google Scholar] [CrossRef]
- Roustan, J.P.; Latche, A.; Fallot, J. Role of ethylene on induction and expression of carrot somatic embryogenesis: Relationship with polyamine metabolism. Plant Sci. 1994, 103, 223–229. [Google Scholar] [CrossRef]
- Bais, H.P.; Sudha, G.S.; Ravishankar, G.A. Influence of putrescine AgNO3 and polyamine inhibitors on the morphogenetic response in untransformed and transformed tissues of Cichorium intybus and their regenerants. Plant Cell Rep. 2001, 20, 547–555. [Google Scholar] [CrossRef]
- Kumar, V.; Parvatam, G.; Ravishankar, G.A. AgNO3: A potential regulator of ethylene activity and plant growth modulator. Elect. J. Biotechnol. 2009, 12, 8–9. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.B.; Yang, S.F. Auxin-induced ethylene production and its inhibition by aminoethyoxyvinylglycine and cobalt ion. Plant Physiol. 1979, 64, 1074–1077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leslie, C.A.; Romani, R.J. Inhibition of ethylene biosynthesis by salicylic acid. Plant Physiol. 1988, 88, 833–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neves, M.; Correia, S.; Cavaleiro, C.; Canhoto, J. Modulation of Organogenesis and Somatic Embryogenesis by Ethylene: An Overview. Plants 2021, 10, 1208. [Google Scholar] [CrossRef] [PubMed]
- Ikeuchi, M.; Ogawa, Y.; Iwase, A.; Sugimoto, K. Plant regeneration: Cellular origins and molecular mechanisms. Development 2016, 143, 1442–1451. [Google Scholar] [CrossRef] [Green Version]
- Bhatia, S.; Bera, T. Somatic embryogenesis and organogenesis. In Modern Applications of Plant Biotechnology in Pharmaceutical Sciences; Bhatia, S., Sharma, K., Dahiya, R., Bera, T., Eds.; Academic Press: Cambridge, MA, USA, 2015; pp. 209–230. [Google Scholar]
- Tamimi, S.M. Effects of ethylene inhibitors, silver nitrate (AgNO3) cobalt chloride (CoCl2) and aminooxyacetic acid (AOA), on in vitro shoot induction and rooting of Banana (Musa acuminata L.). Afr. J. Biotechnol. 2015, 14, 2510–2516. [Google Scholar]
- Giridhar, P.; Indu, E.P.; Vinod, K.; Chandrashekar, A.; Ravishankar, G.A. Direct somatic embryogenesis from Coffea arabica L and Coffea canephora P ex Fr. under the influence of ethylene action inhibitor-silver nitrate. Acta Physiol. Plant 2004, 26, 299–305. [Google Scholar] [CrossRef]
- Qin, Y.; Zhang, S.; Zhang, L.X.; Zhu, D.; Asghar, S. Response of strawberry cv. Toyonokain vitro to silver nitrate (AgNO3). Hortscience 2005, 40, 747–751. [Google Scholar] [CrossRef]
- Gong, Y.; Gao, F.; Tang, K. In vitro high frequency direct root and shoot regeneration in sweet potato using the ethylene inhibitor silver nitrate. S. Afr. J. Bot. 2005, 71, 110–113. [Google Scholar] [CrossRef] [Green Version]
- Abdellatef, E.; Ahmed, M.; Daffall, H.; Khalafalla, M. Enhancement of adventitious shoot regeneration in sesame (Sesamum indicum L.) cultivar Promo ky using ethylene inhibitors. J. Phytol. 2010, 2, 61–67. [Google Scholar]
- Osman, M.G.; Khalafalla, M. Promotion of in vitro shoot formation from shoot tip of tomato (Lycopersicon esculentum mill. cv. omdurman) by ethylene inhibitors. Int. J. Curr. Res. 2010, 4, 82–86. [Google Scholar]
- Dikash, S.; Devala, D.; Punyarani, K.; Henary, S.; Brojendro, S.S.; Brajakishor, S.; Sunitibala, H.D. Silver nitrate and different culture vessels influence high frequency microrhizome induction in vitro and enhancement growth of turmeric plantlet during ex vitro acclimatization. Not. Sci. Biol. 2012, 4, 67–78. [Google Scholar]
- Pua, E.C.; Chi, G.L. De novo shoot morphogenesis and plant growth of mustard (Brassica juncea) in vitro in relation to ethylene. Physiol. Plant 1993, 88, 467–474. [Google Scholar] [CrossRef]
- Abeles, F.B.; Morgan, P.W.; Saltiveit, M.E. Ethylene in Plant Biology, 2nd ed.; Academic Press: San Diego, CA, USA, 1992. [Google Scholar]
- Palei, S.; Das, A.K.; Rout, G.R. In vitro studies of strawberry—An important fruit crop: A review. J. Plant Sci. Res. 2015, 31, 115–131. [Google Scholar]
- Saiprasad, G.V.S.; Raghuveer, P. Influence of ethylene inhibitors and ethrel on production of protocorm like bodies in orchid-dendrobium ‘sonia’. J. Horticul. Sci. 2007, 2, 13–18. [Google Scholar]
- Sankhla, D.; Sankhla, N.; Davis, T.D. Promotion of in vitro shoot formation from excised roots of silk tree by an oxime ether derivative and other ethylene inhibitors. Plant Cell Rep. 1995, 15, 143–146. [Google Scholar] [CrossRef] [PubMed]
- Kong, D.; Shen, H.; Li, N. Influence of AgNO3 on somatic embryo induction and development in Manchurian ash (Fraxinus mandshurica Rupr.). Afr. J. Biotechnol. 2011, 11, 120–125. [Google Scholar]
- Al-Khayri, J.M.; Al-Bahrany, A.M. Genotype-dependent in vitro response of date palm (Phoenix dactylifera L.) cultivars to silver nitrate. Sci. Hortic. 2004, 99, 153–162. [Google Scholar] [CrossRef]
- Kumar, S.; Sarkar, A.K.; Kuhikannan, C. Regeneration of plants from leaflet explants of tissue culture raised Safed Siris (Albizia procera). Plant Cell Tissue Organ Cult. 1998, 54, 137–143. [Google Scholar] [CrossRef]
- Nitsch, J.P.; Nitsch, C. Haploid plants from pollen grains. Science 1969, 163, 85–87. [Google Scholar] [CrossRef]
- Zhao, X.C.; Qu, X.; Mathews, D.E.; Schaller, G.E. Effect of ethylene pathway mutations upon expression of the ethylene receptor ETR1 from Arabidopsis. Plant Physiol. 2002, 130, 1983–1991. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Explant Source | Contamination Rate (%) | Explants Forming Shoots (%) |
|---|---|---|
| Zygotic Embryo of Leccino | 2.1 ± 0.7 | - |
| Nodes of CS-3T | 56.4 ± 3.8 a | 78.4 ± 8.8 |
| Nodes of F7P3 | 33.2 ± 2.9 b | 68.9 ± 10.2 |
| Explant Source | Explant Forming Callus (%) | Amount of Callus | Callus Traits |
|---|---|---|---|
| Zygotic Embryo Leccino | 100 | 2.8 ± 0.3 a | White and friable |
| Variety CS-3T | 90 ± 5 | 2.2 ± 0.3 b | White/yellowish compact |
| Variety F7P3 | 95 ± 3 | 1.9 ± 0.2 b | Yellowish and compact/friable |
| Ethylene Inhibitors | Embryogenic Callus (%) | Number of Somatic Embryos per Callus |
|---|---|---|
| Control | 15.3 ± 2.1 cd | 3.8 ± 0.3 d |
| AgNO3 20 µM | 23.8 ± 4.2 c | 9.2 ± 0.3 bc |
| AgNO3 40 µM | 68.7 ± 8.1 a | 18.6 ± 0.2 a |
| SA 20 µM y | 26.0 ± 4.5 bc | 6.0 ± 0.3 cd |
| SA 40 µM | 60.4 ± 4.1 b | 8.5 ± 0.2 bc |
| CoCl2 20 µM | 12.2 ± 1.5 d | 7.3 ± 0.3 c |
| CoCl2 40 µM | 28.1 ± 4.0 bc | 12.2 ± 0.2 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bashir, M.A.; Silvestri, C.; Salimonti, A.; Rugini, E.; Cristofori, V.; Zelasco, S. Can Ethylene Inhibitors Enhance the Success of Olive Somatic Embryogenesis? Plants 2022, 11, 168. https://doi.org/10.3390/plants11020168
Bashir MA, Silvestri C, Salimonti A, Rugini E, Cristofori V, Zelasco S. Can Ethylene Inhibitors Enhance the Success of Olive Somatic Embryogenesis? Plants. 2022; 11(2):168. https://doi.org/10.3390/plants11020168
Chicago/Turabian StyleBashir, Muhammad Ajmal, Cristian Silvestri, Amelia Salimonti, Eddo Rugini, Valerio Cristofori, and Samanta Zelasco. 2022. "Can Ethylene Inhibitors Enhance the Success of Olive Somatic Embryogenesis?" Plants 11, no. 2: 168. https://doi.org/10.3390/plants11020168