
Effects of Seed Endophytic Bacteria on Life History and Reproductive Traits in a Cosmopolitan Weed, Capsella bursa-pastoris

Abstract
:1. Introduction
2. Materials and Methods
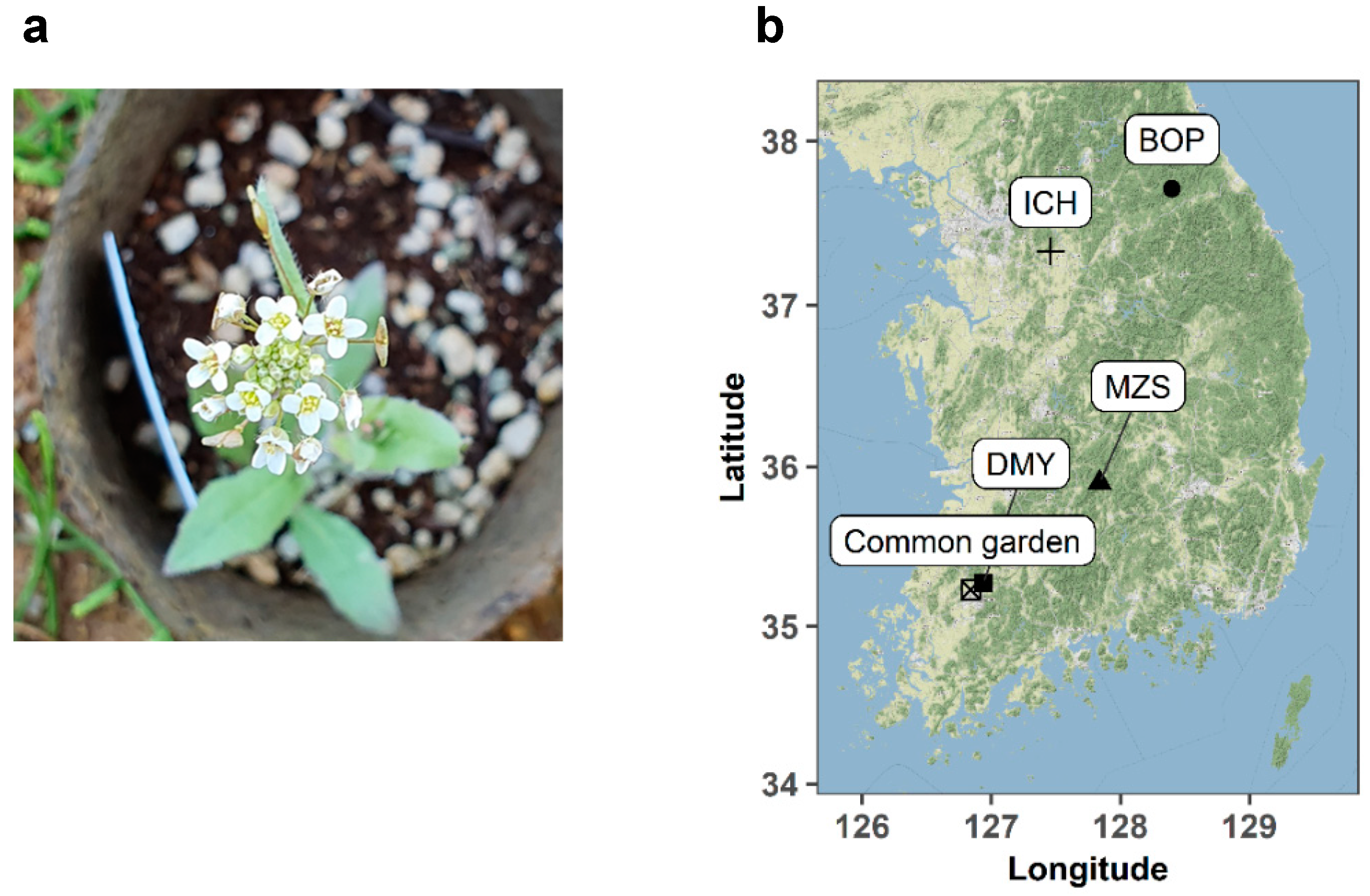
2.1. Seed Collection
2.2. Bacterial Isolation, Identification, and Phenotyping
2.3. Experimental Design
2.4. Statistical Analyses
3. Results
3.1. Bacterial Phenotype
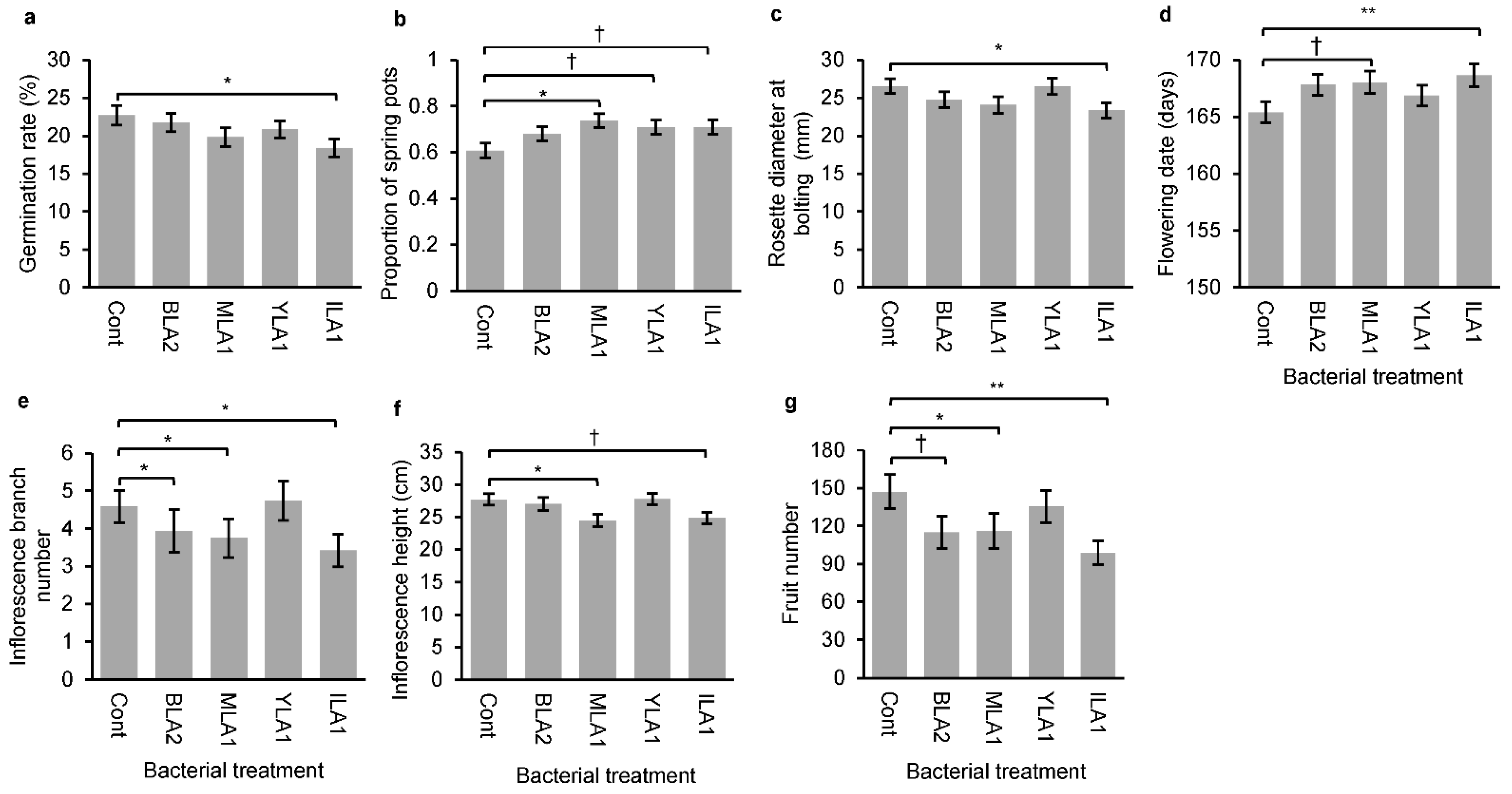
3.2. Effects of Bacterial Inoculation on Plant Traits
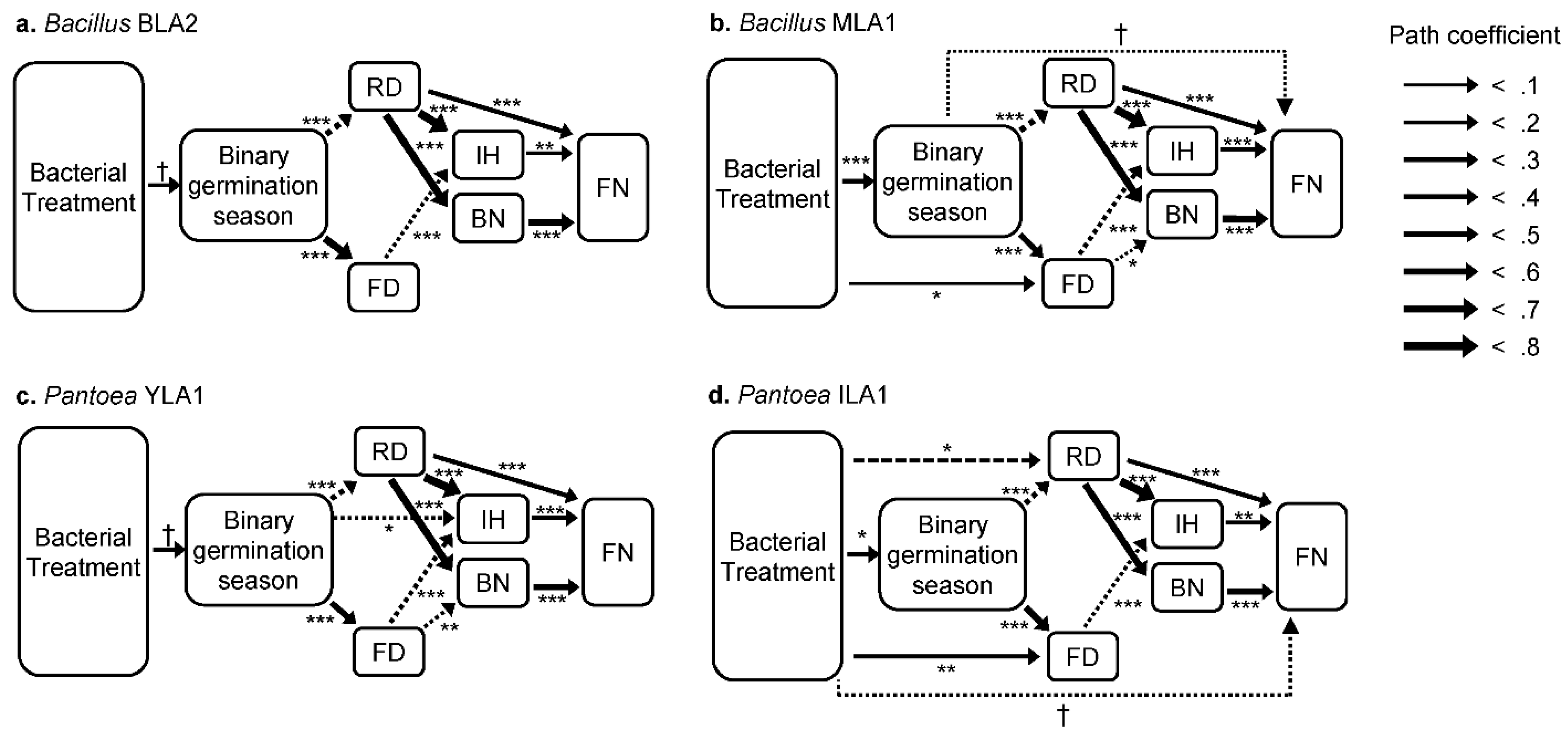
3.3. Direct and Indirect Effects of Bacterial Inoculation on Plant Traits
3.4. Association between Post-Germination Traits and Fitness
4. Discussion
4.1. Effects of Bacterial Treatments on Seed Germination Behavior
4.2. Effects of Bacterial Treatments on Post-Germination Traits
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vandenkoornhuyse, P.; Quaiser, A.; Duhamel, M.; Le Van, A.; Dufresne, A. The importance of the microbiome of the plant holobiont. New Phytol. 2015, 206, 1196–1206. [Google Scholar] [CrossRef]
- Hardoim, P.R.; Van Overbeek, L.S.; Berg, G.; Pirttilä, A.M.; Compant, S.; Campisano, A.; Döring, M.; Sessitsch, A. The hidden world within plants: Ecological and evolutionary considerations for defining functioning of microbial endophytes. Microbiol. Mol. Biol. Rev. 2015, 79, 293–320. [Google Scholar] [CrossRef] [Green Version]
- Petipas, R.H.; Geber, M.A.; Lau, J.A. Microbe-mediated adaptation in plants. Ecol. Lett. 2021, 24, 1302–1317. [Google Scholar] [CrossRef]
- Truyens, S.; Weyens, N.; Cuypers, A.; Vangronsveld, J. Bacterial seed endophytes: Genera, vertical transmission and interaction with plants. Environ. Microbiol. Rep. 2015, 7, 40–50. [Google Scholar] [CrossRef]
- Shade, A.; Jacques, M.-A.; Barret, M. Ecological patterns of seed microbiome diversity, transmission, and assembly. Curr. Opin. Microbiol. 2017, 37, 15–22. [Google Scholar] [CrossRef] [Green Version]
- Verma, S.K.; Kharwar, R.N.; White, J.F. The role of seed-vectored endophytes in seedling development and establishment. Symbiosis 2019, 78, 107–113. [Google Scholar] [CrossRef]
- Soares, M.; Li, H.; Kowalski, K.P.; Bergen, M.; Torres, M.; White, J.F. Functional role of bacteria from invasive Phragmites australis in promotion of host growth. Microb. Ecol. 2016, 72, 407–417. [Google Scholar] [CrossRef]
- Jeong, S.; Kim, T.-M.; Choi, B.; Kim, Y.; Kim, E. Invasive Lactuca serriola seeds contain endophytic bacteria that contribute to drought tolerance. Sci. Rep. 2021, 11, 13307. [Google Scholar] [CrossRef]
- Roff, D. Evolution of Life Histories: Theory and Analysis; Chapman & Hall: London, UK, 1993. [Google Scholar]
- O’Brien, A.M.; Ginnan, N.A.; Rebolleda-Gómez, M.; Wagner, M.R. Microbial effects on plant phenology and fitness. Am. J. Bot. 2021, 108, 1824–1837. [Google Scholar] [CrossRef]
- Wagner, M.R.; Lundberg, D.S.; Coleman-Derr, D.; Tringe, S.G.; Dangl, J.L.; Mitchell-Olds, T. Natural soil microbes alter flowering phenology and the intensity of selection on flowering time in a wild Arabidopsis relative. Ecol. Lett. 2014, 17, 717–726. [Google Scholar] [CrossRef] [Green Version]
- Santoyo, G.; Moreno-Hagelsieb, G.; del Carmen Orozco-Mosqueda, M.; Glick, B.R. Plant growth-promoting bacterial endophytes. Microbiol. Res. 2016, 183, 92–99. [Google Scholar] [CrossRef]
- Taulé, C.; Vaz-Jauri, P.; Battistoni, F. Insights into the early stages of plant–endophytic bacteria interaction. World J. Microbiol. Biotechnol. 2021, 37, 13. [Google Scholar] [CrossRef] [PubMed]
- Choi, B.; Jeong, S.; Kim, E. Variation of the seed endophytic bacteria among plant populations and their plant growth-promoting activities in a wild mustard plant species, Capsella bursa-pastoris. Ecol. Evol. 2022, 12, e8683. [Google Scholar] [CrossRef] [PubMed]
- Donohue, K.; Rubio de Casas, R.; Burghardt, L.; Kovach, K.; Willis, C.G. Germination, postgermination adaptation, and species ecological ranges. Annu. Rev. Ecol. Evol. Syst. 2010, 41, 293–319. [Google Scholar] [CrossRef]
- Burghardt, L.T.; Metcalf, C.J.E.; Wilczek, A.M.; Schmitt, J.; Donohue, K. Modeling the influence of genetic and environmental variation on the expression of plant life cycles across landscapes. Am. Nat. 2015, 185, 212–227. [Google Scholar] [CrossRef] [Green Version]
- Donohue, K. Niche construction through phenological plasticity: Life history dynamics and ecological consequences. New Phytol. 2005, 166, 83–92. [Google Scholar] [CrossRef]
- Cheplick, G.P. Can endosymbiotic microbes modulate natural selection in plant populations? An example with Lolium perenne and its fungal endophyte. Symbiosis 2018, 76, 321–327. [Google Scholar] [CrossRef]
- Gremer, J.R.; Wilcox, C.J.; Chiono, A.; Suglia, E.; Schmitt, J. Germination timing and chilling exposure create contingency in life history and influence fitness in the native wildflower Streptanthus tortuosus. J. Ecol. 2020, 108, 239–255. [Google Scholar] [CrossRef] [Green Version]
- Neuffer, B. Native range variation in Capsella bursa-pastoris (Brassicaceae) along a 2500 km latitudinal transect. Flora 2011, 206, 107–119. [Google Scholar] [CrossRef]
- Aksoy, A.; Dixon, J.M.; Hale, W.H. Capsella bursa-pastoris (L.) Medikus (Thlaspi bursa-pastoris L., Bursa bursa-pastoris (L.) Shull, Bursa pastoris (L.) Weber). J. Ecol. 1998, 86, 171–186. [Google Scholar] [CrossRef]
- Chang, Y.; Ha, Y.; Maeng, D. Germination behavoir of Capsella bursa-pastoris medicus and the density effect on barley grain yield. Korean J. Weed Sci. 1986, 6, 13–17. [Google Scholar]
- Neuffer, B.; Bartelheim, S. Gen-ecology of Capsella bursa-pastoris from an altitudinal transsect in the Alps. Oecologia 1989, 81, 521–527. [Google Scholar] [CrossRef]
- Sunar, K.; Dey, P.; Chakraborty, U.; Chakraborty, B. Biocontrol efficacy and plant growth promoting activity of Bacillus altitudinis isolated from Darjeeling hills, India. J. Basic Microbiol. 2015, 55, 91–104. [Google Scholar] [CrossRef]
- Bhattacharyya, C.; Bakshi, U.; Mallick, I.; Mukherji, S.; Bera, B.; Ghosh, A. Genome-guided insights into the plant growth promotion capabilities of the physiologically versatile Bacillus aryabhattai strain AB211. Front. Microbiol. 2017, 8, 411. [Google Scholar] [CrossRef] [Green Version]
- Castagno, L.; Estrella, M.; Sannazzaro, A.; Grassano, A.; Ruiz, O. Phosphate-solubilization mechanism and in vitro plant growth promotion activity mediated by Pantoea eucalypti isolated from Lotus tenuis rhizosphere in the Salado River Basin (Argentina). J. Appl. Microbiol. 2011, 110, 1151–1165. [Google Scholar] [CrossRef]
- Sharavin, D. Halotolerant phosphate solubilizing IAA-synthesizing representatives of Pantoea genus promote proline synthesis in wheat seedlings under salt stress. J. Agric. Environ. 2021, 2, 18. [Google Scholar]
- Glick, B.R. Plant growth-promoting bacteria: Mechanisms and applications. Scientifica 2012, 2012, 963401. [Google Scholar] [CrossRef] [Green Version]
- Etesami, H.; Emami, S.; Alikhani, H.A. Potassium solubilizing bacteria (KSB): Mechanisms, promotion of plant growth, and future prospects. J. Soil Sci. Plant Nutr. 2017, 17, 897–911. [Google Scholar] [CrossRef]
- Nautiyal, C.S. An efficient microbiological growth medium for screening phosphate solubilizing microorganisms. FEMS Microbiol. Lett. 1999, 170, 265–270. [Google Scholar] [CrossRef]
- Fiske, C.H.; Subbarow, Y. The colorimetric determination of phosphorus. J. Biol. Chem. 1925, 66, 375–400. [Google Scholar] [CrossRef]
- Schwyn, B.; Neilands, J. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Murakami, C.; Tanaka, A.R.; Sato, Y.; Kimura, Y.; Morimoto, K. Easy detection of siderophore production in diluted growth media using an improved CAS reagent. J. Microbiol. Methods 2021, 189, 106310. [Google Scholar] [CrossRef]
- Patten, C.L.; Glick, B.R. Role of Pseudomonas putida indoleacetic acid in development of the host plant root system. Appl. Environ. Microbiol. 2002, 68, 3795–3801. [Google Scholar] [CrossRef] [Green Version]
- Penrose, D.M.; Glick, B.R. Methods for isolating and characterizing ACC deaminase-containing plant growth-promoting rhizobacteria. Physiol. Plant. 2003, 118, 10–15. [Google Scholar] [CrossRef] [Green Version]
- Lowry, O.; Rosebrough, N.; Farr, A.L.; Randall, R. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Govindarajan, M.; Balandreau, J.; Kwon, S.-W.; Weon, H.-Y.; Lakshminarasimhan, C. Effects of the inoculation of Burkholderia vietnamensis and related endophytic diazotrophic bacteria on grain yield of rice. Microb. Ecol. 2008, 55, 21–37. [Google Scholar] [CrossRef]
- Griffith, C.; Kim, E.; Donohue, K. Life-history variation and adaptation in the historically mobile plant Arabidopsis thaliana (Brassicaceae) in North America. Am. J. Bot. 2004, 91, 837–849. [Google Scholar] [CrossRef] [Green Version]
- Choi, B.; Jeong, H.; Kim, E. Phenotypic plasticity of Capsella bursa-pastoris (Brassicaceae) and its effect on fitness in response to temperature and soil moisture. Plant Species Biol. 2019, 34, 5–10. [Google Scholar] [CrossRef]
- Mitchell, R.J. Path analysis. In Design and Analysis of Ecological Experiments, 1st ed.; Scheiner, S.M., Gurevitch, J., Eds.; Oxford University Press: Oxford, UK, 2001. [Google Scholar]
- Lande, R.; Arnold, S.J. The measurement of selection on correlated characters. Evolution 1983, 37, 1210–1226. [Google Scholar] [CrossRef]
- Mitchell-Olds, T.; Shaw, R.G. Regression analysis of natural selection: Statistical inference and biological interpretation. Evolution 1987, 41, 1149–1161. [Google Scholar] [CrossRef]
- Lenth, R. emmeans: Estimated Marginal Means, aka Least-Squares Means, R Package Version 1.8.1-1. 2022. Available online: https://cran.r-project.org/web/packages/emmeans/emmeans.pdf (accessed on 30 August 2022).
- Verma, S.K.; Kingsley, K.; Irizarry, I.; Bergen, M.; Kharwar, R.; White, J., Jr. Seed-vectored endophytic bacteria modulate development of rice seedlings. J. Appl. Microbiol. 2017, 122, 1680–1691. [Google Scholar] [CrossRef]
- Graeber, K.; Nakabayashi, K.; Miatton, E.; Leubner-Metzger, G.; Soppe, W.J. Molecular mechanisms of seed dormancy. Plant Cell Environ. 2012, 35, 1769–1786. [Google Scholar] [CrossRef] [PubMed]
- Penfield, S. Seed dormancy and germination. Curr. Biol. 2017, 27, R874–R878. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Su, K.; Li, C.; White, J.F. Interactive effects of Epichloë endophyte, dormancy-breaking treatments and geographic origin on seed germination of Achnatherum inebrians. Microorganisms 2021, 9, 2183. [Google Scholar] [CrossRef] [PubMed]
- Linkies, A.; Leubner-Metzger, G. Beyond gibberellins and abscisic acid: How ethylene and jasmonates control seed germination. Plant Cell Rep. 2012, 31, 253–270. [Google Scholar] [CrossRef] [PubMed]
- Corbineau, F.; Xia, Q.; Bailly, C.; El-Maarouf-Bouteau, H. Ethylene, a key factor in the regulation of seed dormancy. Front. Plant Sci. 2014, 5, 539. [Google Scholar] [CrossRef] [Green Version]
- Glick, B.R.; Todorovic, B.; Czarny, J.; Cheng, Z.; Duan, J.; McConkey, B. Promotion of plant growth by bacterial ACC deaminase. Crit. Rev. Plant Sci. 2007, 26, 227–242. [Google Scholar] [CrossRef]
- Wu, M.; Wu, J.; Gan, Y. The new insight of auxin functions: Transition from seed dormancy to germination and floral opening in plants. Plant Growth Regul. 2020, 91, 169–174. [Google Scholar] [CrossRef]
- Donohue, K. Germination timing influences natural selection on life-history characters in Arabidopsis thaliana. Ecology 2002, 83, 1006–1016. [Google Scholar] [CrossRef]
- Akiyama, R.; Ågren, J. Conflicting selection on the timing of germination in a natural population of Arabidopsis thaliana. J. Evol. Biol. 2014, 27, 193–199. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Source Population | Strains (Accession Number) | The Closest Type Strains | rDNA Sequence Similarity (%) | Plant Growth-Promoting Activity (mean ± se) | |||
|---|---|---|---|---|---|---|---|
| ACC Deaminase (nmol α-KB/mg Protein • h) | Phosphate Solubilization (mg/L of PO4) | Siderophore Production (psu, % of Siderophore Units) | Auxin Production (μg/mL) | ||||
| BOP | BLA2 (ON329310) | Bacillus altitudinis 41KF2b | 1410/1414 (99.72) | - | 27.75 ± 1.36 | 2.96 ± 1.19 | 2.68 ± 0.19 |
| MZS | MLA1 (ON329312) | Bacillus aryabhattai B8W22 | 1409/1410 (99.93) | 37.56 ± 0.38 | 653.95 ± 4.93 | 2.77 ± 1.18 | 21.16 ± 0.53 |
| DMY | YLA1 (ON329311) | Pantoea eucalypti LMG 24198 | 1339/1347 (99.41) | 33.88 ± 0.84 | 293.24 ± 5.48 | 24.51 ± 0.28 | 1.94 ± 0.14 |
| ICH | ILA1 (ON329314) | Pantoea vagans LMG 24199 | 1377/1379 (99.85) | 30.51 ± 0.33 | 87.34 ± 3.60 | 26.37 ± 0.50 | 3.87 ± 0.36 |
| Trait | Treatment (d.f = 4) |
|---|---|
| Germination rate | 2.74 * |
| Germination season | 10.20 * |
| Bolting success | 0.52 |
| Flowering date | 2.75 * |
| Rosette diameter | 2.45 * |
| Inflorescence height | 3.11 * |
| Branch number | 3.50 ** |
| Fruit number | 3.64 * |
| Control (N = 68) | Bacillus BLA2 (N = 49) | Bacillus MLA1 (N = 39) | Pantoea YLA1 (N = 53) | |||||
|---|---|---|---|---|---|---|---|---|
| Traits | S | β | S | β | S | β | S | β |
| FD | −0.43 *** (0.12) | −0.16 (0.10) | −0.07 (0.13) | −0.02 (0.05) | −0.47 ** (0.15) | −0.06 (0.07) | −0.22 (0.11) | −0.02 (0.07) |
| RD | 0.66 *** (0.09) | 0.16 (0.16) | 0.78 *** (0.11) | 0.20 * (0.08) | 0.91 *** (0.10) | 0.25 * (0.10) | 0.78 *** (0.09) | 0.37 * (0.15) |
| IH | 0.72 *** (0.09) | 0.28 (0.17) | 0.49 ** (0.14) | 0.12 (0.07) | 0.80 *** (0.12) | 0.19 * (0.09) | 0.73 *** (0.08) | 0.27 * (0.13) |
| BN | 0.60 *** (0.09) | 0.31 ** (0.11) | 1.20 *** (0.07) | 0.97 *** (0.07) | 0.99 *** (0.07) | 0.66 *** (0.08) | 0.63 *** (0.10) | 0.27 ** (0.10) |
| PantoeaILA1 (N = 44) | FS (Trait × Treatment) | Fβ (Trait × Treatment) | ||||||
| Traits | S | β | ||||||
| FD | −0.20 (0.13) | 0.05 (0.07) | 1.55 | 1.34 | ||||
| RD | 0.90 *** (0.13) | 0.33 ** (0.11) | 1.17 | 0.31 | ||||
| IH | 0.67 *** (0.11) | 0.18 (0.11) | 0.09 | 0.56 | ||||
| BN | 0.87 *** (0.08) | 0.69 *** (0.07) | 8.35 *** | 7.72 *** | ||||
| Control (N = 96) | Bacillus BLA2 (N = 111) | Bacillus MLA1 (N = 115) | Pantoea YLA1 (N = 109) | |||||
|---|---|---|---|---|---|---|---|---|
| Traits | S | β | S | β | S | β | S | β |
| FD | −0.41 *** (0.12) | −0.04 (0.07) | −0.21 * (0.10) | 0.06 (0.05) | −0.23 ** (0.08) | 0.01 (0.04) | −0.23 * (0.12) | −0.01 (0.04) |
| RD | 0.98 *** (0.08) | 0.32 * (0.13) | 0.71 *** (0.07) | 0.23 ** (0.07) | 0.65 *** (0.05) | 0.14 * (0.06) | 1.05 *** (0.07) | 0.35 *** (0.08) |
| IH | 0.87 *** (0.09) | 0.22 (0.11) | 0.72 *** (0.07) | 0.28 *** (0.07) | 0.71 *** (0.05) | 0.34 *** (0.07) | 0.95 *** (0.08) | 0.15 * (0.07) |
| BN | 0.99 *** (0.07) | 0.61 *** (0.09) | 0.81 *** (0.06) | 0.56 *** (0.06) | 0.69 *** (0.05) | 0.39 *** (0.05) | 1.14 *** (0.05) | 0.82 *** (0.06) |
| PantoeaILA1 (N = 105) | FS (Trait × Treatment) | Fβ (Trait × Treatment) | ||||||
| Traits | S | β | ||||||
| FD | −0.21 ** (0.08) | −0.06 (0.05) | 1.03 | 1.40 | ||||
| RD | 0.52 *** (0.06) | 0.29 ** (0.09) | 10.03 *** | 0.80 | ||||
| IH | 0.48 *** (0.06) | 0.03 (0.09) | 5.81 *** | 1.68 | ||||
| BN | 0.55 *** (0.06) | 0.40 *** (0.06) | 16.62 *** | 7.68 *** | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, B.; Kim, T.-M.; Jeong, S.; Kim, Y.; Kim, E. Effects of Seed Endophytic Bacteria on Life History and Reproductive Traits in a Cosmopolitan Weed, Capsella bursa-pastoris. Plants 2022, 11, 2642. https://doi.org/10.3390/plants11192642
Choi B, Kim T-M, Jeong S, Kim Y, Kim E. Effects of Seed Endophytic Bacteria on Life History and Reproductive Traits in a Cosmopolitan Weed, Capsella bursa-pastoris. Plants. 2022; 11(19):2642. https://doi.org/10.3390/plants11192642
Chicago/Turabian StyleChoi, Byungwook, Tae-Min Kim, Seorin Jeong, Yousuk Kim, and Eunsuk Kim. 2022. "Effects of Seed Endophytic Bacteria on Life History and Reproductive Traits in a Cosmopolitan Weed, Capsella bursa-pastoris" Plants 11, no. 19: 2642. https://doi.org/10.3390/plants11192642