
Molecular Breeding of Water-Saving and Drought-Resistant Rice for Blast and Bacterial Blight Resistance

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
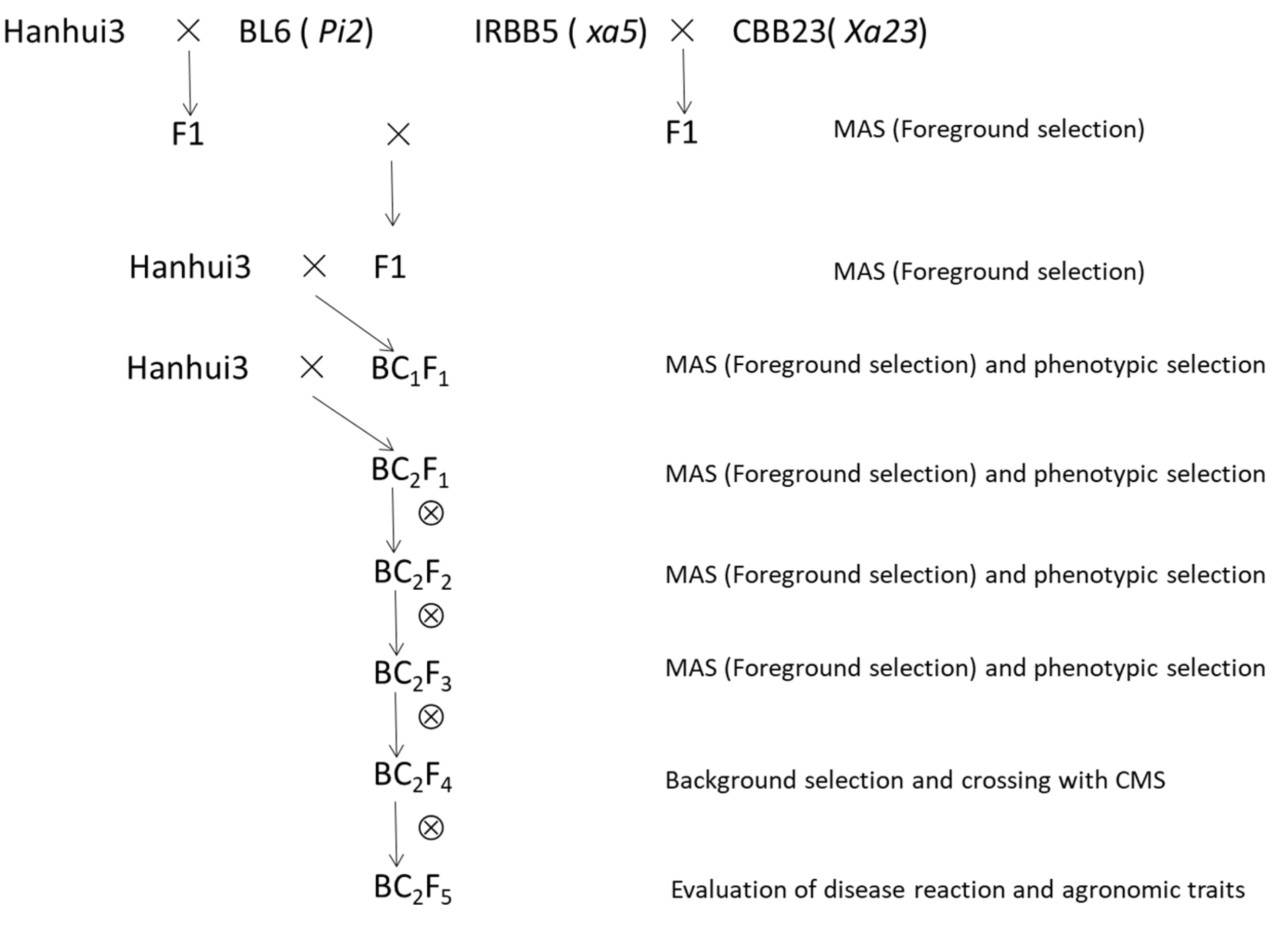
2.1. Rice Materials
2.2. Marker-Assisted Selection of NIL for Blast and BB Resistance
2.3. DNA Extraction, Markers, and Genotyping
2.4. Screening for Blast Resistance
2.5. Screening for BB Resistance
2.6. Evaluation of Major Agronomic Traits under Field Conditions
2.7. Data Analysis
3. Results
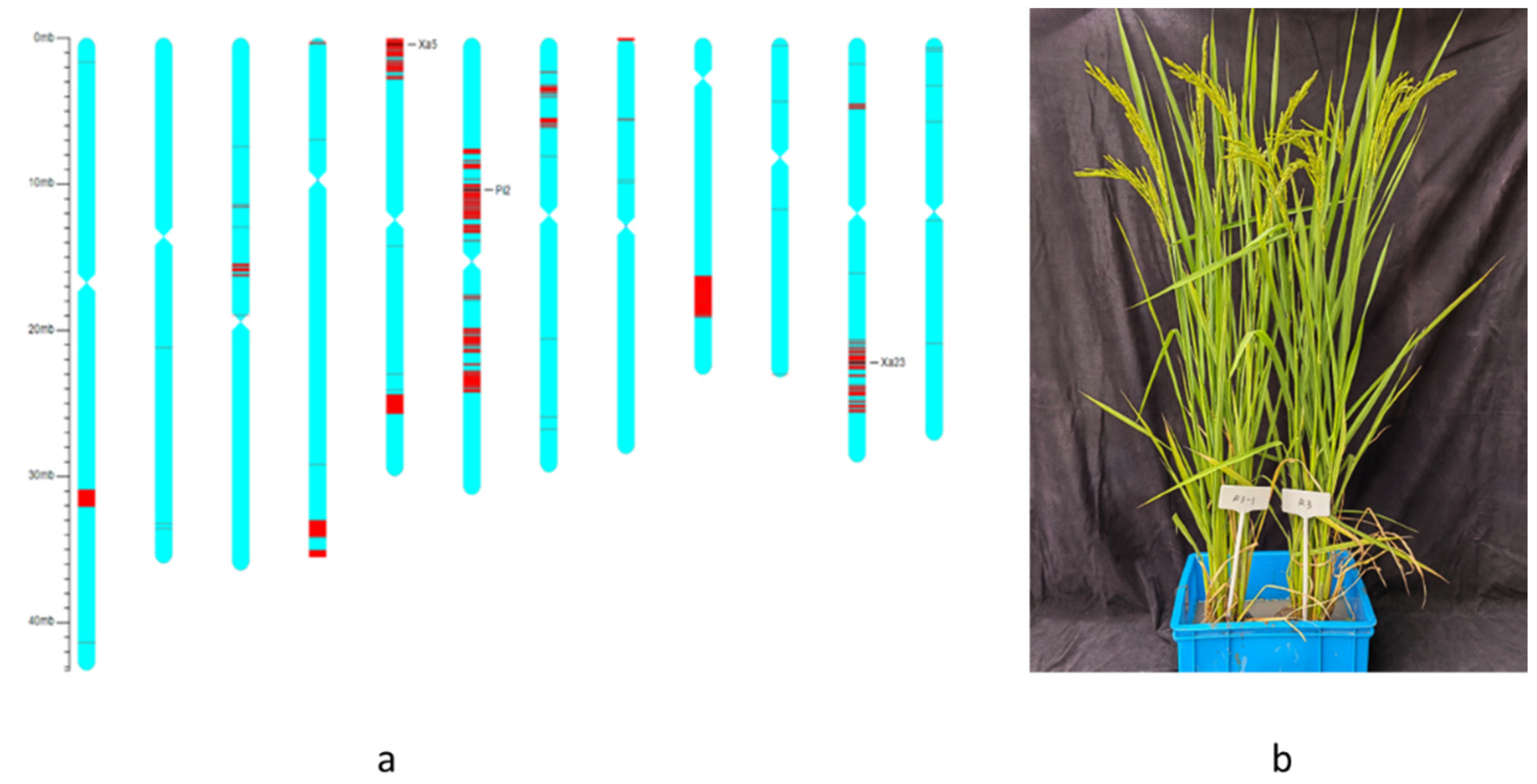
3.1. Development of NIL with Pi2, xa5, and Xa23 Genes Using Marker-Assisted Selection
3.2. Blast and BB Resistance of the Improved NIL and Its Hybrids
3.3. The Performance of Agronomic Traits of the NIL and Its Hybrids
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhang, Q.F. Strategies for developing green super rice. Proc. Natl. Acad. Sci. USA 2007, 104, 16402–16409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, L.P. Breeding for superior high yielding of hybrid rice. Hybrid. Rice 1997, 12, 1–3, (In Chinese with English Abstract). [Google Scholar]
- Khush, G.S. What is will take to feed five billion rice consumers by 2030. Plant Mol. Biol. 2005, 59, 1–6. [Google Scholar] [CrossRef]
- Khush, G.S.; Jena, K.K. Current status and future prospects for research on blast resistance in rice (Oryza sativa L.). In Advances in Genetics, Genomics and Control of Rice Blast Disease; Springer: Dordrecht, The Netherlands, 2009; pp. 1–10. [Google Scholar]
- Barman, S.R.; Chattoo, B.B. Rice blast fungus sequenced. Curr. Sci. 2005, 89, 930–931. [Google Scholar]
- Chen, H.Q.; Chen, Z.X.; Ni, S.; Zuo, S.M.; Pan, X.B.; Zhu, X.D. Pyramiding three genes with resistance to blast by marker assisted selection to improve rice blast resistance of Jin23B. Chin. J. Rice Sci. 2008, 22, 23–27, (In Chinese with English Abstract). [Google Scholar]
- Singh, V.K.; Singh, A.; Singh, S.P.; Ellur, R.K.; Singh, D.; Gopala Krishnan, S.; Bhowmick, P.K.; Nagarajan, M.; Vinod, K.K.; Singh, U.D.; et al. Marker-assisted simultaneous but step wise back cross breeding for pyramiding blast resistance genes Piz5 and Pi54 into an elite Basmati rice restorer line ‘PRR78’. Plant Breed. 2013, 132, 486–495. [Google Scholar]
- Jiang, J.F.; Mou, T.M.; Yu, H.H.; Zhou, F.S. Molecular breeding of thermo-sensitive genic male sterile (TGMS) lines of rice for blast resistance using Pi2 gene. Rice 2015, 8, 11. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.Y.; Yu, L.; Pan, C.H.; Dai, Z.Y.; Li, Y.H.; Xiao, N.; Zhang, X.X.; Ji, H.J.; Huang, N.S.; Zhao, B.H.; et al. Development of near isogenic lines with different alleles of Piz locus and analysis of their breeding effect under Yangdao 6 background. Mol. Breed. 2016, 36, 1–12. [Google Scholar] [CrossRef]
- Miah, G.; Rafii, M.Y.; Ismail, M.R.; Puteh, A.B.; Rahim, H.A.; Latif, M.A. Marker-assisted introgression of broad-spectrum blast resistance genes into the cultivated MR219 rice variety. J. Sci. Food Agric. 2017, 97, 2810–2818. [Google Scholar] [CrossRef]
- Ni, D.H.; Song, F.S.; Ni, J.L.; Zhang, A.F.; Wang, C.L.; Zhao, K.J.; Yang, Y.C.; Wei, P.C.; Yang, J.B.; Li, L. Marker-assisted selection of two-line hybrid rice for disease resistance to rice blast and bacterial blight. Field Crops Res. 2015, 184, 1–8. [Google Scholar] [CrossRef]
- Luo, W.L.; Huang, M.; Guo, T.; Xiao, W.M.; Wang, J.F.; Yang, G.L.; Liu, Y.Z.; Wang, H.; Chen, Z.Q.; Zhuang, C.X. Marker-assisted selection for rice blast resistance genes Pi2 and Pi2 through high-resolution melting of a gene-targeted amplicon. Plant Breed. 2016, 136, 67–73. [Google Scholar] [CrossRef]
- Luo, Y.C.; Ma, T.C.; Zhang, A.F.; Ong, K.H.; Luo, Z.X.; Li, Z.F.; Yang, J.B.; Yin, Z.C. Marker assisted breeding of Chinese elite rice cultivar 9311 for disease resistance to rice blast and bacterial blight and tolerance to submergence. Mol. Breed. 2017, 37, 106–128. [Google Scholar] [CrossRef]
- Qu, S.; Liu, G.; Zhou, B.; Bellizzi, M.; Zeng, L.; Han, B.; Wang, G.L. The broad-spectrum blast resistance gene Pi2 encodes an NBS-LRR protein and is a member of the multiple family in rice. Genetics 2006, 172, 1901–1914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, D.S.; Xia, M.Y.; Li, J.B.; Wan, B.L.; Zha, Z.P.; Dd, X.S.; Qi, H.X. Development of STS marker linked to rice blast resistance gene Pi2 in marker-assisted selection breeding. Chin. J. Rice Sci. 2011, 25, 25–30, (In Chinese with English Abstract). [Google Scholar]
- Srinivasan, B.; Gnanamanickam, S. Identification of a new source of resistance in wild rice, Oryza rufipogon to bacterial blight of rice caused by Indian strains of Xanthomonas oryzae pv. Oryzae. Curr. Sci. 2005, 88, 1229–1231. [Google Scholar]
- Singh, S.; Sidhu, J.S.; Huang, N.; Vikal, Z.Y.; Li, Z.; Brar, D.S.; Dhaliwal, H.S.; Khush, G.S. Pyramiding three bacterial blight resistance genes (xa5, xa13 and Xa21) using marker-assisted selection into indica rice cultivar PR106. Theor. Appl. Genet. 2001, 102, 1011–1015. [Google Scholar] [CrossRef]
- Narayanan, N.N.; Baisakh, N.; Vera Cruz, N.; Gnananmanickam, S.S.; Datta, K.; Datta, S.K. Molecular breeding for the development of blast and bacterial blight resistance in rice cv. IR50. Crop Sci. 2002, 42, 2072–2079. [Google Scholar] [CrossRef]
- Hari, Y.; Srinivasarao, K.; Viraktamath, B.C.; Hariprasad, A.S.; Laha, G.S.; Ilyas Ahmed, M.; Natarajkumar, P.; Ramesha, M.S.; Neeraja, C.N.; Balachandran, S.M.; et al. Marker-assisted improvement of a stable restorer line, KMR-3Rand its derived hybrid KRH2 for bacterial blight resistance and grain quality. Plant Breed. 2011, 130, 608–616. [Google Scholar] [CrossRef]
- Suh, J.P.; Cho, Y.C.; Won, Y.J.; Ahn, E.K.; Baek, M.K.; Kim, M.K.; Kim, B.K.; Jena, K.K. Development of resistant gene-pyramided japonica rice for multiple biotic stresses using molecular marker-assisted selection. Plant Breed. Biotech. 2015, 3, 333–345. [Google Scholar] [CrossRef] [Green Version]
- Hur, Y.J.; Cho, J.H.; Park, H.S.; Noh, T.H.; Park, D.S.; Lee, J.Y.; Sohn, Y.B.; Shin, D.; Song, Y.C.; Kwon, Y.U.; et al. Pyramiding of two rice bacterial blight resistance genes, Xa3 and Xa4, and a closely linked cold-tolerance QTL on chromosome 11. Theor. Appl. Genet. 2016, 129, 1861–1871. [Google Scholar] [CrossRef]
- Zhou, Y.L.; Uzokwe, V.N.E.; Zhang, C.H.; Cheng, L.R.; Wang, L.; Chen, K.; Gao, X.Q.; Sun, Y.; Chen, J.J.; Zhu, L.H.; et al. Improvement of bacterial blight resistance of hybrid rice in China using the Xa23 gene derived from wild rice (Oryza rufipogon). Crop Prot. 2011, 30, 637–644. [Google Scholar] [CrossRef]
- Pan, H.J.; Wang, C.L.; Zhao, K.J.; Zhang, Q.; Fan, Y.L.; Zhou, S.C.; Zhu, L.H. Molecular mapping by PCR-based markers and marker-assisted selection of Xa23, a bacterial blight resistance gene in rice. Acta Agron. Sin. 2003, 29, 501–507, (In Chinese with English Abstract). [Google Scholar]
- Zhang, Q. Highlights in identification and application of resistance genes to bacterial blight. Chin. J. Rice Sci. 2005, 19, 453–459, (In Chinese with English Abstract). [Google Scholar]
- Luo, L.J. Breeding for water-saving and drought-resistance rice (WDR) in China. J. Exp. Bot. 2010, 61, 3509–3517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, A.N.; Li, M.S.; Wang, F.M.; Liu, G.L.; Yu, X.Q. Breeding and utilization of Water-saving and Drought-resistant rice restorer line Hanhui3. Hybrid. Rice 2010, 28, 13–15, (In Chinese with English Abstract). [Google Scholar]
- Carvalho, F.P. Agriculture, pesticides, food security and food safety. Environ. Sci. Policy 2006, 9, 685–692. [Google Scholar] [CrossRef]
- Dellaporta, S.L.; Wood, J.; Hicks, J.B. A plant DNA minipreparation: Version II. Plant Mol. Biol. Rep. 1983, 1, 19–21. [Google Scholar] [CrossRef]
- Sanguinetti, C.J.; Dias, N.E.; Simpson, A.J.G. Rapid silver staining and recovery of PCR products separated on polyacrylamide gels. Biotechniques 1994, 17, 915–919. [Google Scholar]
- Wu, J.H.; Jiang, J.S.; Chen, H.L.; Wang, S.P. Fine mapping of rice blast resistance gene Pi-2(t). Acta Agron. Sin. 2002, 28, 505–509. [Google Scholar]
- Blair, M.W.; Garris, A.J.; Iyer, A.S.; Chapman, B.; Kresovich, S.; McCouch, S.R. High resolution genetic mapping and candidate gene identification at the xa5 locus for bacterial blight resistance in rice (Oryza sativa L.). Theor. Appl. Genet. 2003, 107, 62–73. [Google Scholar] [CrossRef]
- SES. Standard Evaluation System for Rice; International Rice Research Institute: Manila, Philippines, 2002; pp. 1–56. [Google Scholar]
- Wang, F.M.; Liu, Y.; Zhang, A.N.; Kong, D.Y.; Bi, J.G.; Liu, G.L.; Yu, X.Q.; Luo, L.J. Breeding an early maturing, blast resistance water-saving and drought-resistance rice (WDR) cultivar using marker-assisted selection coupled with rapid generation advance. Mol. Breed. 2022, 42, 46. [Google Scholar] [CrossRef]
- Peng, S.B.; Tang, Q.Y.; Zou, Y.B. Current Status and Challenges of rice production in China. Plant Prod. Sci. 2009, 12, 3–8. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.Q.; Liu, G.L.; Li, M.S.; Pan, Z.Q.; Zhang, A.N.; Zhu, J.L.; Luo, L.J. Hanyou 73, a new water-saving and drought-resistant hybrid rice combination. Hybrid. Rice 2016, 31, 79–81, (In Chinese with English Abstract). [Google Scholar]
- Liu, W.Q.; Fan, Y.Y.; Chen, J.; Shi, Y.F.; Wu, J.L. Avoidance of linkage drag between blast resistance gene and the QTL conditioning spikelet fertility based on genotype selection against heading date in paddy. Rice Sci. 2009, 16, 21–26. [Google Scholar] [CrossRef]
- Chu, Z.H.; Fu, B.Y.; Yang, H.; Xu, C.G.; Li, Z.K.; Sanchez, A.; Park, Y.J.; Bennetzen, J.L.; Zhang, Q.F.; Wang, S.P. Targeting xa13, a recessive gene for bacterial blight resistance in rice. Theor. Appl. Genet. 2006, 112, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Li, X.; Wu, C.J.; Yang, C.J.; Hua, H.X.; Gao, G.J.; Xiao, J.H.; He, Y.Q. Pyramiding and evaluation of the brown planthopper resistance genes Bph14 and Bph15 in hybrid rice. Mol. Breed. 2012, 29, 61–69. [Google Scholar] [CrossRef]
- Wang, H.B.; Ye, S.T.; Mou, T.M. Molecular breeding of rice restorer lines and hybrids for brown planthopper (BPH) resistance using the Bph14 and Bph15 genes. Rice 2016, 9, 53. [Google Scholar] [CrossRef] [Green Version]
- Frisch, M.; Bohn, M.; Melchinger, A.E. Comparison of selection strategies for marker-assisted backcrossing of a gene. Crop Sci. 1999, 39, 1295–1301. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| R Genes | Chr. No. | Marker Name | Primer Sequences (5′-3′) | Exp. Size (bp) | Band Type | Reference | |
|---|---|---|---|---|---|---|---|
| Forward Primer (F) | Reverse Primer (R) | ||||||
| Pi2 | 6 | AP22 | GTGCATGAGTCCAGCTCAAA | GTGTACTCCCATGGCTGCTC | 143 | Co-dominant | [30] |
| xa5 | 5 | RM611 | CAACAAGATGGCCTCTTACC | TACAAACAAACAGCTTGTGC | 213 | Co-dominant | [31] |
| Xa23 | 11 | RM206 | CCCATGCGTTTAACTATTCT | CGTTCCATCGATCCGTATGG | 147 | Co-dominant | [23] |
| Rice Line | Resistance Genes Genotype # | Reaction against Blast * | Reaction against BB * | ||||
|---|---|---|---|---|---|---|---|
| Pi2 | xa5 | Xa23 | Score | Disease Reaction | Lesion Length | Disease Reaction | |
| Hanhui3 | -- | -- | -- | 7.5 | S | 15.22 ± 3.24 | S |
| R3-1 | ++ | ++ | ++ | 3.5 | MR | 1.51 ± 0.05 | R |
| 7A/Hanhui3 | -- | -- | -- | 5 | MS | 8.82 ± 2.18 | MS |
| 7A/R3-1 | +- | +- | +- | 3 | MR | 2.28 ± 0.37 | R |
| 5A/Hanhui3 | -- | -- | -- | 5.5 | MS | 9.03 ± 2.31 | MS |
| 5A/R3-1 | +- | +- | +- | 2.5 | MR | 2.09 ± 1.21 | R |
| Rice Line | Days to 50% Flowering | Plant Height (cm) | Panicle Length (cm) | No. of Tillers | No. of Grains per Panicle | Spikelet Fertility (%) | 1000-Seed Weight (g) | Yield per Plant (g) |
|---|---|---|---|---|---|---|---|---|
| Hanhui3 | 102.2 ± 3.5 | 117.45 ± 4.82 | 21.32 ± 1.83 | 6.65 ± 0.64 | 186.54 ± 8.71 | 87.34 ± 6.15 | 26.85 ± 0.44 | 29.11 ± 5.62 |
| R3-1 | 102.7 ± 1.2 | 115.13 ± 3.61 | 22.04 ± 0.87 | 6.75 ± 2.21 | 201.09 ± 7.81 * | 85.27 ± 1.78 | 26.29 ± 0.31 | 28.38 ± 3.61 |
| 7A/Hanhui3 | 88.0 ± 2.1 | 123.50 ± 6.14 | 24.15 ± 0.54 | 7.54 ± 1.04 | 177.32 ± 21.36 | 84.52 ± 2.84 | 29.33 ± 0.88 | 34.21 ± 3.89 |
| 7A/R3-1 | 88.3 ± 1.8 | 122.77 ± 5.57 | 25.72 ± 0.21 * | 7.44 ± 0.69 | 165.51 ± 10.37 | 88.19 ± 1.85 * | 30.68 ± 1.75 | 37.19 ± 2.41 |
| 5A/Hanhui3 | 93.2 ± 4.3 | 116.50 ± 2.64 | 23.52 ± 0.85 | 9.42 ± 0.53 | 182.13 ± 7.62 | 79.42 ± 1.64 | 26.41 ± 1.58 | 40.92 ± 6.48 |
| 5A/R3-1 | 91.4 ± 3.1 | 116.35 ± 5.71 | 24.74 ± 0.36 | 9.82 ± 0.82 | 185.12 ± 11.46 | 85.12 ± 3.68 | 25.98 ± 1.23 | 42.63 ± 5.63 |
| Rice Line | Days to 50% Flowering | Plant Height (cm) | Panicle Length (cm) | No. of Tillers | No. of Grains per Panicle | Spikelet Fertility (%) | 1000-Seed Weight (g) | Yield per Plant (g) |
|---|---|---|---|---|---|---|---|---|
| Hanhui3 | 105.3 ± 3.21 | 87.06 ± 2.46 | 17.92 ± 1.54 | 5.33 ± 2.57 | 89.22 ± 8.52 | 69.30 ± 4.48 | 23.28 ± 1.76 | 15.06 ± 1.22 |
| R3-1 | 105.9 ± 2.42 | 86.50 ± 4.23 | 19.76 ± 2.58 | 5.00 ± 1.52 | 100.58 ± 11.72 | 78.97 ± 5.72 | 24.52 ± 1.51 | 14.95 ± 0.98 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, A.; Liu, Y.; Wang, F.; Kong, D.; Bi, J.; Zhang, F.; Luo, X.; Wang, J.; Liu, G.; Luo, L.; et al. Molecular Breeding of Water-Saving and Drought-Resistant Rice for Blast and Bacterial Blight Resistance. Plants 2022, 11, 2641. https://doi.org/10.3390/plants11192641
Zhang A, Liu Y, Wang F, Kong D, Bi J, Zhang F, Luo X, Wang J, Liu G, Luo L, et al. Molecular Breeding of Water-Saving and Drought-Resistant Rice for Blast and Bacterial Blight Resistance. Plants. 2022; 11(19):2641. https://doi.org/10.3390/plants11192641
Chicago/Turabian StyleZhang, Anning, Yi Liu, Feiming Wang, Deyan Kong, Junguo Bi, Fenyun Zhang, Xingxing Luo, Jiahong Wang, Guolan Liu, Lijun Luo, and et al. 2022. "Molecular Breeding of Water-Saving and Drought-Resistant Rice for Blast and Bacterial Blight Resistance" Plants 11, no. 19: 2641. https://doi.org/10.3390/plants11192641