
Alpha-Mangostin as a New Therapeutic Candidate for Concanavalin A-Induced Autoimmune Hepatitis: Impact on the SIRT1/Nrf2 and NF-κB Crosstalk

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. General Procedures
2.2. Extraction and Isolation of α-MN
Spectral Data of α-MN
2.3. Animals
2.4. Induction of AIH and Experimental Groups
- Control: the vehicle for 7 days.
- α-MN: only α-MN (50 mg/kg, dissolved in soybean oil) orally for 7 days.
- Con A: only Con A (15 mg/kg, single IV dose).
- α-MN 25 + Con A, mice were given α-MN (25 mg/kg, orally once a day for 7 days) prior to the Con A injection in the last day.
- α-MN 50 + Con A, mice were given α-MN (50 mg/kg, orally once a day for 7 days) prior to the Con A injection in the last day.
2.5. Biochemical Investigation
2.5.1. Serum Indices of AIH
2.5.2. Myeloperoxidase (MPO)
2.5.3. Oxidative Parameters and Antioxidants
2.5.4. NF-κB and Inflammatory Cytokines
2.5.5. Nrf2 and Heme Oxygenase-1 (HO-1)
2.6. Liver Histopathology
2.7. RT-PCR Assessment
2.8. Immunohistochemistry (IHC)
2.9. Statistics
3. Results
3.1. α-MN Alleviated Con A-Induced Hepatic Lesions
3.2. α-MN Decreased the Con A-Induced Infiltration of the Neutrophiles and the CD4+ T Cells into the Liver
3.3. α-MN Ameliorated the Con A-Induced Oxidative Stress and the Enhanced Antioxidants
3.4. α-MN Counteracted the Con A-Induced Suppression in the SIRT1/Nrf2/HO-1 Signaling
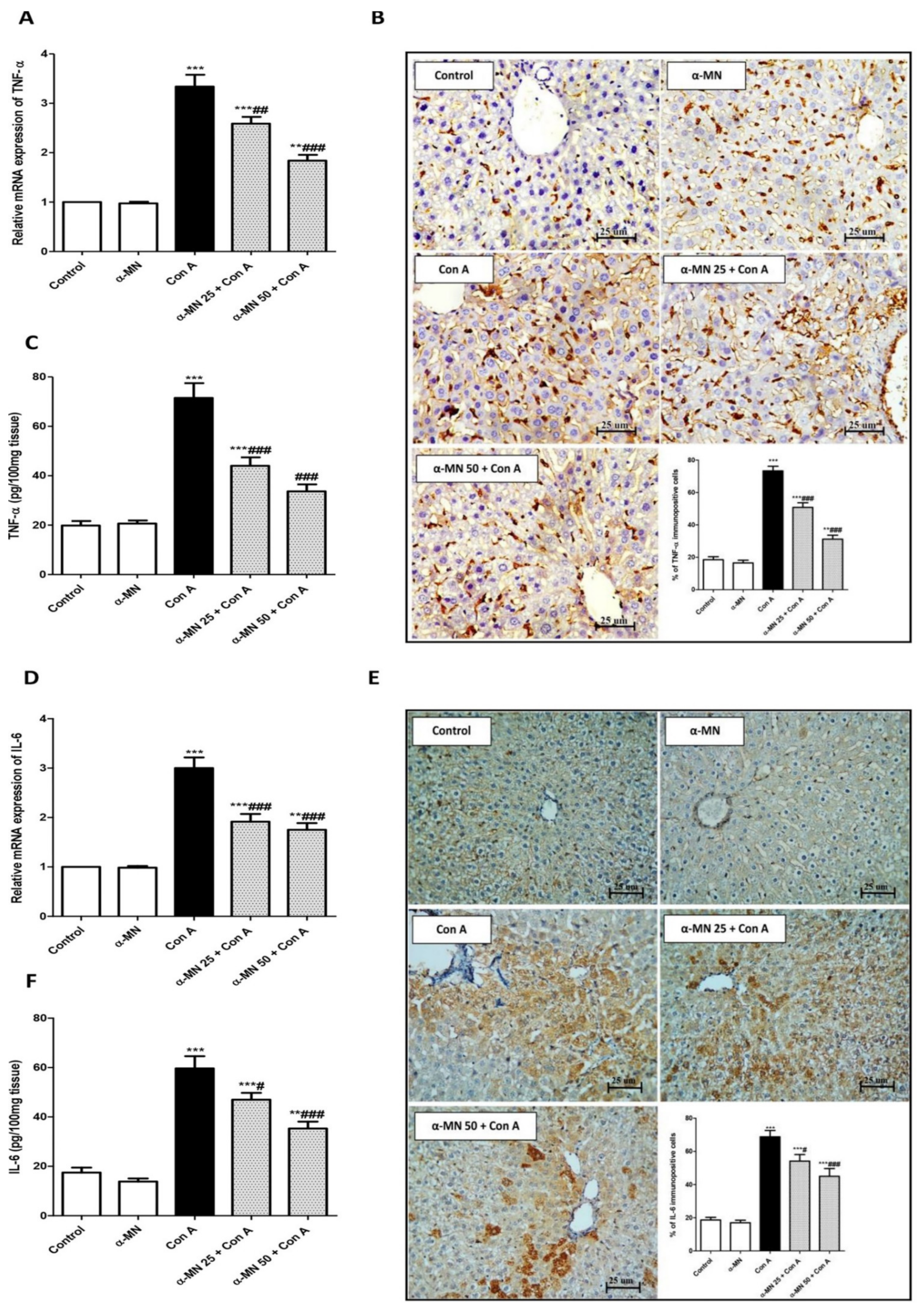
3.5. α-MN Prohibited the NF-κB/TNF-α/IL-6 Signaling Activation Induced by Con A
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hao, J.; Sun, W.; Xu, H. Pathogenesis of concanavalin A induced autoimmune hepatitis in mice. Int. Immunopharmacol. 2022, 102, 108411. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.R.M.; Sirwi, A.; Eid, B.G.; Mohamed, S.; Mohamed, G.A. Summary of natural products ameliorate Concanavalin A-induced liver injury: Structures, sources, pharmacological effects, and mechanisms of action. Plants 2021, 10, 228. [Google Scholar] [CrossRef]
- Ding, Y.; Yu, Z.; Zhang, C. Diallyl trisulfide protects against concanavalin A-induced acute liver injury in mice by inhibiting inflammation, oxidative stress and apoptosis. Life Sci. 2021, 278, 119631. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Li, Q.; Zhou, C.; Zhao, Y.; Li, R.; Zhang, Y. Demethyleneberberine attenuates concanavalin A-induced autoimmune hepatitis in mice through inhibition of NF-kappaB and MAPK signaling. Int. Immunopharmacol. 2020, 80, 106137. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.H.; Li, H.L.; Li, Y.X.; You, Y.; Guan, Y.Y.; Zhang, S.L.; Liu, L.X.; Bao, W.L.; Zhou, Y.; Shen, X.Y. Protective effects of a traditional Chinese herbal formula Jiang-Xian HuGan on Concanavalin A-induced mouse hepatitis via NF-kappaB and Nrf2 signaling pathways. J. Ethnopharmacol. 2018, 217, 118–125. [Google Scholar] [CrossRef]
- El-Agamy, D.S.; Shaaban, A.A.; Almaramhy, H.H.; Elkablawy, S.; Elkablawy, M.A. Pristimerin as a novel hepatoprotective agent against experimental autoimmune hepatitis. Front. Pharmacol. 2018, 9, 292. [Google Scholar] [CrossRef]
- Sharawy, M.H.; El-Kashef, D.H.; Shaaban., A.A.; El-Agamy, D.S. Anti-fibrotic activity of sitagliptin against concanavalin A-induced hepatic fibrosis. Role of Nrf2 activation/NF-kappaB inhibition. Int. Immunopharmacol. 2021, 100, 108088. [Google Scholar] [CrossRef]
- Ahmed, N.; Bakhashwain, N.F.; Alsehemi, A.F.; El-Agamy, D.S. Hepatoprotective role of vardenafil against experimentally induced hepatitis in mice. J. Biochem. Mol. Toxicol. 2017, 31, e21867. [Google Scholar] [CrossRef]
- Heymann, F.; Hamesch, K.; Weiskirchen, R.; Tacke, F. The concanavalin A model of acute hepatitis in mice. Lab. Anim. 2015, 49 (Suppl. S1), 12–20. [Google Scholar] [CrossRef]
- Magadula, J.J. A bioactive isoprenylated xanthone and other constituents of Garcinia edulis. Fitoterapia 2010, 81, 420–423. [Google Scholar] [CrossRef]
- Gutierrez-Orozco, F.; Failla, M.L. Biological activities and bioavailability of mangosteen xanthones: A critical review of the current evidence. Nutrients 2013, 5, 3163–3183. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.R.M.; Mohamed, G.A.; Khayat, M.T.; Ahmed, S.; Abo-Haded, H.; Alshali, K.Z. Mangostanaxanthone VIIII, a new xanthone from Garcinia mangostana pericarps, α-amylase inhibitory activity, and molecular docking studies. Rev. Bras. Farmacogn. 2019, 29, 206–212. [Google Scholar] [CrossRef]
- Ibrahim, S.R.M.; Mohamed, G.A.; Elfaky, M.A.; Al Haidari, R.A.; Zayed, M.F.; El-Kholy, A.A.; Ross, S.A. Garcixanthone A, a new cytotoxic xanthone from the pericarps of Garcinia mangostana. J. Asian Nat. Prod. Res. 2019, 21, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.R.M.; Abdallah, H.M.; El-Halawany, A.M.; Nafady, A.M.; Mohamed, G.A. Mangostanaxanthone VIII, a new xanthone from Garcinia mangostana and its cytotoxic activity. Nat. Prod. Res. 2019, 33, 258–265. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, G.A.; Ibrahim, S.R.M.; Shaaban, M.I.A.; Ross, S.A. Mangostanaxanthones I and II, new xanthones from the pericarp of Garcinia mangostana. Fitoterapia 2014, 98, 215–221. [Google Scholar] [CrossRef]
- Mohamed, G.A.; Al-Abd, A.M.; El-Halawany, A.M.; Abdallah, H.M.; Ibrahim, S.R.M. New xanthones and cytotoxic constituents from Garcinia mangostana fruit hulls against human hepatocellular, breast, and colorectal cancer cell lines. J. Ethnopharmacol. 2017, 198, 302–312. [Google Scholar] [CrossRef]
- Chavan, T.; Muth, A. The diverse bioactivity of alpha-mangostin and its therapeutic implications. Future Med. Chem. 2021, 13, 1679–1694. [Google Scholar] [CrossRef]
- Chen, G.; Li, Y.; Wang, W.; Deng, L. Bioactivity and pharmacological properties of alpha-mangostin from the mangosteen fruit: A review. Expert Opin. Ther. Pat. 2018, 28, 415–427. [Google Scholar] [CrossRef]
- Larson, R.T.; Lorch, J.M.; Pridgeon, J.W.; Becnel, J.J.; Clark, G.G.; Lan, Q. The biological activity of alpha-mangostin, a larvicidal botanic mosquito sterol carrier protein-2 inhibitor. J. Med. Entomol. 2010, 47, 249–257. [Google Scholar]
- Supawadee, S.; Thanet, S.; Wisut, P.; Somneuk, N.; Sirinun, N.; Ramida, W. Investigation of Therapeutic Effects of α-mangostin on thioacetamide-induced cirrhosis in rats. J. Med. Assoc. Thail. 2015, 98 (Suppl. S9), S91–S97. [Google Scholar]
- Tsai, S.Y.; Chung, P.C.; Owaga, E.E.; Tsai, I.J.; Wang, P.Y.; Tsai, J.I.; Yeh, T.S.; Hsieh, R.H. Alpha-mangostin from mangosteen (Garcinia mangostana Linn.) pericarp extract reduces high fat-diet induced hepatic steatosis in rats by regulating mitochondria function and apoptosis. Nutr. Metab. 2016, 13, 88. [Google Scholar] [CrossRef] [PubMed]
- Lestari, N.; Louisa, M.; Soetikno, V.; Suwana, A.G.; Ramadhan, P.A.; Akmal, T.; Arozal, W. Alpha mangostin inhibits the proliferation and activation of acetaldehyde induced hepatic stellate cells through TGF-β and ERK 1/2 pathways. J. Toxicol. 2018, 2018, 5360496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alhakamy, N.A.; Mohamed, G.A.; Fahmy, U.A.; Eid, B.G.; Ahmed, O.; Al-Rabia, M.W.; Khedr, A.; Nasrullah, M.Z.; Ibrahim, S. New alpha-amylase inhibitory metabolites from pericarps of Garcinia mangostana. Life 2022, 12, 384. [Google Scholar] [CrossRef]
- Liandhajani; Iwo, M.I.; Sukrasno; Soemardji, A.A.; Hanafi, M. Sunscreen activity of α-mangostin from the pericarps of Garcinia mangostana. Linn. J. Appl. Pharm. Sci. 2013, 3, 70–73. [Google Scholar]
- El-Agamy, D.S. Pirfenidone ameliorates concanavalin A-induced hepatitis in mice via modulation of reactive oxygen species/nuclear factor kappa B signalling pathways. J. Pharm. Pharmacol. 2016, 68, 1559–1566. [Google Scholar] [CrossRef]
- Márquez-Valadez, B.; Lugo-Huitrón, R.; Valdivia-Cerda, V.; Miranda-Ramírez, L.R.; Pérez-De La Cruz, V.; González-Cuahutencos, O.; Rivero-Cruz, I.; Mata, R.; Santamaría, A.; Pedraza-Chaverrí, J. The natural xanthone alpha-mangostin reduces oxidative damage in rat brain tissue. Nutr. Neurosci. 2009, 12, 35–42. [Google Scholar] [CrossRef]
- Pérez-Rojas, J.M.; Cruz, C.; García-López, P.; Sánchez-González, D.J.; Martínez-Martínez, C.M.; Ceballos, G.; Espinosa, M.; Meléndez-Zajgla, J.; Pedraza-Chaverri, J. Renoprotection by alpha-mangostin is related to the attenuation in renal oxidative/nitrosative stress induced by cisplatin nephrotoxicity. Free Radic. Res. 2009, 43, 1122–1132. [Google Scholar] [CrossRef]
- Sampath, P.; Vijayaraghavan, K. Cardioprotective effect of alpha-mangostin, a xanthone derivative from mangosteen on tissue defense system against isoproterenol-induced myocardial infarction in rats. J. Biochem. Mol. Toxicol. 2007, 21, 336–339. [Google Scholar] [CrossRef]
- Tian, X.; Liu, Y.; Liu, X.; Gao, S.; Sun, X. Glycyrrhizic acid ammonium salt alleviates Concanavalin A-induced immunological liver injury in mice through the regulation of the balance of immune cells and the inhibition of hepatocyte apoptosis. Biomed. Pharmacother. 2019, 120, 109481. [Google Scholar] [CrossRef]
- Kim, H.M.; Kim, Y.M.; Huh, J.H.; Lee, E.S.; Kwon, M.H.; Lee, B.R.; Ko, H.J.; Chung, C.H. alpha-Mangostin ameliorates hepatic steatosis and insulin resistance by inhibition C-C chemokine receptor 2. PLoS ONE 2017, 12, e0179204. [Google Scholar]
- Fu, T.; Li, H.; Zhao, Y.; Cai, E.; Zhu, H.; Li, P.; Liu, J. Hepatoprotective effect of α-mangostin against lipopolysaccharide/d-galactosamine-induced acute liver failure in mice. Biomed. Pharmacother. 2018, 106, 896–901. [Google Scholar] [CrossRef] [PubMed]
- Fu, T.; Wang, S.; Liu, J.; Cai, E.; Li, H.; Li, P.; Zhao, Y. Protective effects of alpha-mangostin against acetaminophen-induced acute liver injury in mice. Eur. J. Pharmacol. 2018, 827, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Tan, Y.P.; Zhao, L.; Wang, L.; Fu, N.J.; Zheng, S.P.; Shen, X.F. Anticancer activity of dietary xanthone alpha-mangostin against hepatocellular carcinoma by inhibition of STAT3 signaling via stabilization of SHP1. Cell Death Dis. 2020, 11, 63. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, G.A.; Ibrahim, S.R.M.; El-Agamy, D.S.; Elsaed, W.M.; Sirwi, A.; Asfour, H.Z.; Koshak, A.E.; Elhady, S.S. Cucurbitacin E glucoside alleviates concanavalin A-induced hepatitis through enhancing SIRT1/Nrf2/HO-1 and inhibiting NF-ĸB/NLRP3 signaling pathways. J. Ethnopharmacol. 2022, 292, 115223. [Google Scholar] [CrossRef] [PubMed]
- Fei, M.; Xie, Q.; Zou, Y.; He, R.; Zhang, Y.; Wang, J.; Bo, L.; Li, J.; Deng, X. Alpha-lipoic acid protects mice against concanavalin A-induced hepatitis by modulating cytokine secretion and reducing reactive oxygen species generation. Int. Immunopharmacol. 2016, 35, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Eisvand, F.; Imenshahidi, M.; Ghasemzadeh Rahbardar, M.; Tabatabaei Yazdi, S.A.; Rameshrad, M.; Razavi, B.M.; Hosseinzadeh, H. Cardioprotective effects of alpha-mangostin on doxorubicin-induced cardiotoxicity in rats. Phytother. Res. 2022, 36, 506–524. [Google Scholar] [CrossRef] [PubMed]
- Herrera-Aco, D.R.; Medina-Campos, O.N.; Pedraza-Chaverri, J.; Sciutto-Conde, E.; Rosas-Salgado, G.; Fragoso-González, G. Alpha-mangostin: Anti-inflammatory and antioxidant effects on established collagen-induced arthritis in DBA/1J mice. Food Chem. Toxicol. 2019, 124, 300–315. [Google Scholar] [CrossRef]
- Tatiya-Aphiradee, N.; Chatuphonprasert, W.; Jarukamjorn, K. Ethanolic Garcinia mangostana extract and alpha-mangostin improve dextran sulfate sodium-induced ulcerative colitis via the suppression of inflammatory and oxidative responses in ICR mice. J. Ethnopharmacol. 2021, 265, 113384. [Google Scholar] [CrossRef]
- Huang, T.H.; Chen, C.C.; Liu, H.M.; Lee, T.Y.; Shieh, S.H. Resveratrol pretreatment attenuates Concanavalin A-induced hepatitis through reverse of aberration in the immune response and regenerative capacity in aged mice. Sci. Rep. 2017, 7, 2705. [Google Scholar] [CrossRef]
- Xu, X.; Hu, Y.; Zhai, X.; Lin, M.; Chen, Z.; Tian, X.; Zhang, F.; Gao, D.; Ma, X.; Lv, L.; et al. Salvianolic acid A preconditioning confers protection against concanavalin A-induced liver injury through SIRT1-mediated repression of p66shc in mice. Toxicol. Appl. Pharmacol. 2013, 273, 68–76. [Google Scholar] [CrossRef]
- Singh, V.; Ubaid, S. Role of silent information regulator 1 (SIRT1) in regulating oxidative stress and inflammation. Inflammation 2020, 43, 1589–1598. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Song, C.; Chen, J.; Zhou, L.; Jiang, X.; Cao, X.; Sun, Y.; Zhang, Q. Limonin ameliorates acetaminophen-induced hepatotoxicity by activating Nrf2 antioxidative pathway and inhibiting NF-κB inflammatory response via upregulating Sirt1. Phytomedicine 2020, 69, 153211. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.Y.; Deng, J.S.; Huang, W.C.; Jiang, W.P.; Huang, G.J. Attenuation of lipopolysaccharide-induced acute lung injury by Hispolon in mice, through regulating the TLR4/PI3K/Akt/mTOR and Keap1/Nrf2/HO-1 pathways, and suppressing oxidative stress-mediated eR Stress-induced apoptosis and autophagy. Nutrients 2020, 12, 1742. [Google Scholar] [CrossRef] [PubMed]
- Franceschelli, S.; Pesce, M.; Ferrone, A.; Patruno, A.; Pasqualone, L.; Carlucci, G.; Ferrone, V.; Carlucci, M.; de Lutiis, M.A.; Grilli, A.; et al. A Novel biological role of alpha-mangostin in modulating inflammatory response through the activation of SIRT-1 signaling pathway. J. Cell. Physiol. 2016, 231, 2439–2451. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Song, Z.; Wang, H.; Li, Q.; Cui, Z.; Zhang, Y. Angelica sinensis polysaccharide attenuates concanavalin A-induced liver injury in mice. Int. Immunopharmacol. 2016, 31, 140–148. [Google Scholar] [CrossRef] [PubMed]
- AboElnazar, S.Y.; Ghoneim, H.E.; Ghazy, A.A.; El-Shamandy, M.I.; Kazem, A.H.; Rashwan, E.K. Concanavalin-A as a model for onduction of murine autoimmune hepatitis: Role of TNF-α and NF-κβ during the acute phase. Egypt J. Immunol. 2020, 27, 19–30. [Google Scholar]
- Guan, H.; Li, J.; Tan, X.; Luo, S.; Liu, Y.; Meng, Y.; Wu, B.; Zhou, Y.; Yang, Y.; Chen, H.; et al. Natural xanthone alpha-mangostin inhibits LPS-induced microglial inflammatory responses and memory impairment by blocking the TAK1/NF-kappaB signaling pathway. Mol. Nutr. Food Res. 2020, 64, e2000096. [Google Scholar] [CrossRef]
- Yang, K.; Yin, Q.; Mao, Q.; Dai, S.; Wang, L.; Dong, J.; Zuo, J. Metabolomics analysis reveals therapeutic effects of alpha-mangostin on collagen-induced arthritis in rats by down-regulating nicotinamide phosphoribosyltransferase. Inflammation 2019, 42, 741–753. [Google Scholar] [CrossRef]
- You, B.H.; Chae, H.S.; Song, J.; Ko, H.W.; Chin, Y.W.; Choi, Y.H. alpha-Mangostin ameliorates dextran sulfate sodium-induced colitis through inhibition of NF-kappaB and MAPK pathways. Int. Immunopharmacol. 2017, 49, 212–221. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene (Mouse) | Accession | PCR Product (bp) | Sequence (5′-3′) |
|---|---|---|---|
| SIRT1 | NM_019812.3 | 111 | F: CGATGACAGAACGTCACACG |
| R: ATTGTTCGAGGATCGGTGCC | |||
| Nrf2 | NM_010902 | 170 | F: AAGAATAAAGTCGCCGCCCA |
| R: AGATACAAGGTGCTGAGCCG | |||
| HO-1 | NM_010442 | 200 | F: CCTCACAGATGGCGTCACTT |
| R: TGGGGGCCAGTATTGCATTT | |||
| GCLc | NM_010295 | 182 | F: CTTTGGGTCGCAAGTAGGAAGC |
| R: GGGCGTCCCGTCCGTTC | |||
| NQO1 | NM_008706 | 111 | F: CATTGCAGTGGTTTGGGGTG |
| R: TCTGGAAAGGACCGTTGTCG | |||
| NF-ĸB | AY521463.1 | 102 | F: AGGAAGGCAAAGCGAATCCA |
| R: TCAGAACCAAGAAGGACGCC | |||
| TNF-α | NM_011609.4 | 102 | F: GCTGTTGCCCCTGGTTATCT |
| R: ATGGAGTAGACTTCGGGCCT | |||
| IL-6 | NM_031168.2 | 79 | F: AGTCCTTCCTACCCCAATTTCC |
| R: GGTCTTGGTCCTTAGCCACT | |||
| β-actin | NM_007393.5 | 198 | F: TGAGCTGCGTTTTACACCCT |
| R: GCCTTCACCGTTCCAGTTTT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shehata, A.M.; Elbadawy, H.M.; Ibrahim, S.R.M.; Mohamed, G.A.; Elsaed, W.M.; Alhaddad, A.A.; Ahmed, N.; Abo-Haded, H.; El-Agamy, D.S. Alpha-Mangostin as a New Therapeutic Candidate for Concanavalin A-Induced Autoimmune Hepatitis: Impact on the SIRT1/Nrf2 and NF-κB Crosstalk. Plants 2022, 11, 2441. https://doi.org/10.3390/plants11182441
Shehata AM, Elbadawy HM, Ibrahim SRM, Mohamed GA, Elsaed WM, Alhaddad AA, Ahmed N, Abo-Haded H, El-Agamy DS. Alpha-Mangostin as a New Therapeutic Candidate for Concanavalin A-Induced Autoimmune Hepatitis: Impact on the SIRT1/Nrf2 and NF-κB Crosstalk. Plants. 2022; 11(18):2441. https://doi.org/10.3390/plants11182441
Chicago/Turabian StyleShehata, Ahmed M., Hossein M. Elbadawy, Sabrin R. M. Ibrahim, Gamal A. Mohamed, Wael M. Elsaed, Aisha A. Alhaddad, Nishat Ahmed, Hany Abo-Haded, and Dina S. El-Agamy. 2022. "Alpha-Mangostin as a New Therapeutic Candidate for Concanavalin A-Induced Autoimmune Hepatitis: Impact on the SIRT1/Nrf2 and NF-κB Crosstalk" Plants 11, no. 18: 2441. https://doi.org/10.3390/plants11182441