
Nitrogen and Phosphorus of Plants Associated with Arbuscular and Ectomycorrhizas Are Differentially Influenced by Drought
Abstract
:1. Introduction
2. Results
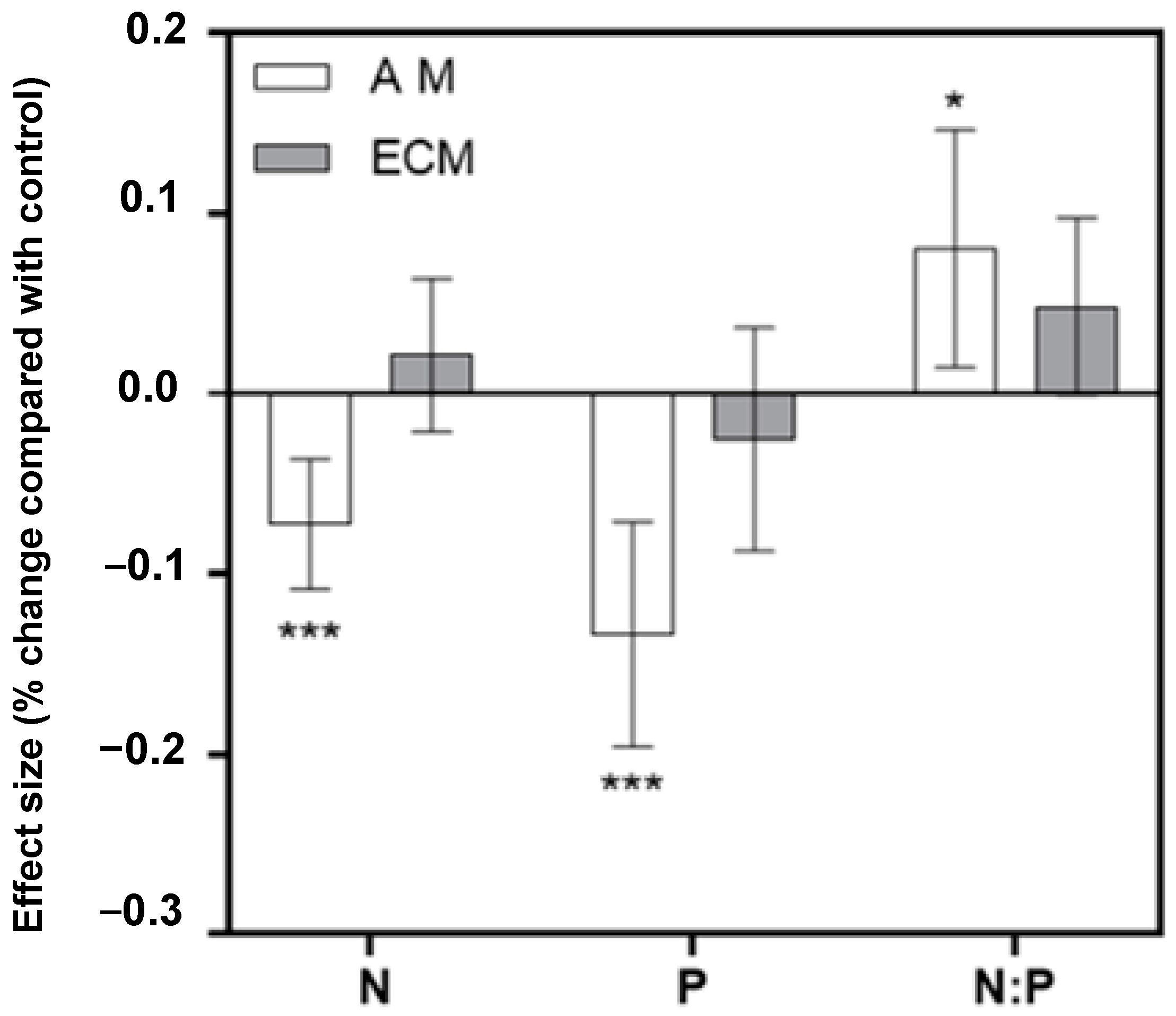
2.1. Overall Differences in the Effects of DS on AM and ECM Plant N, P, and N:P Ratio
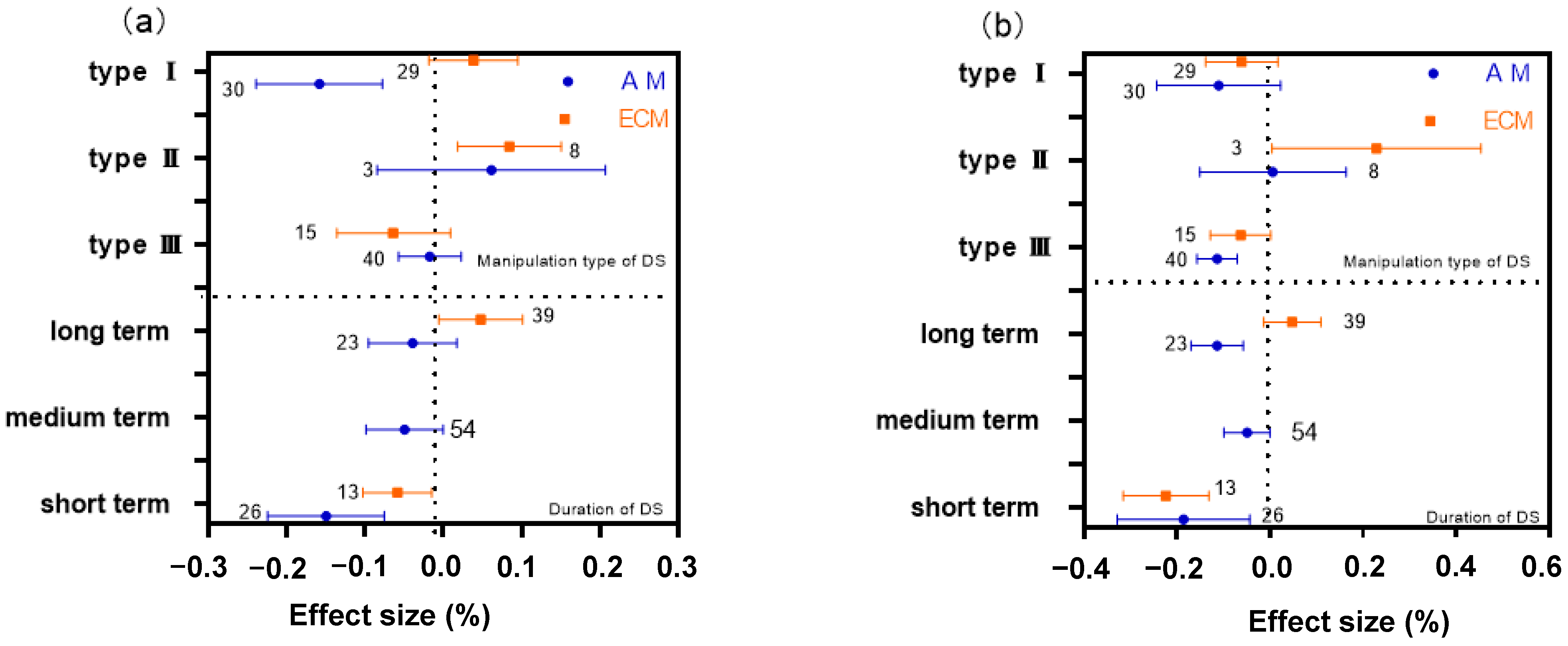
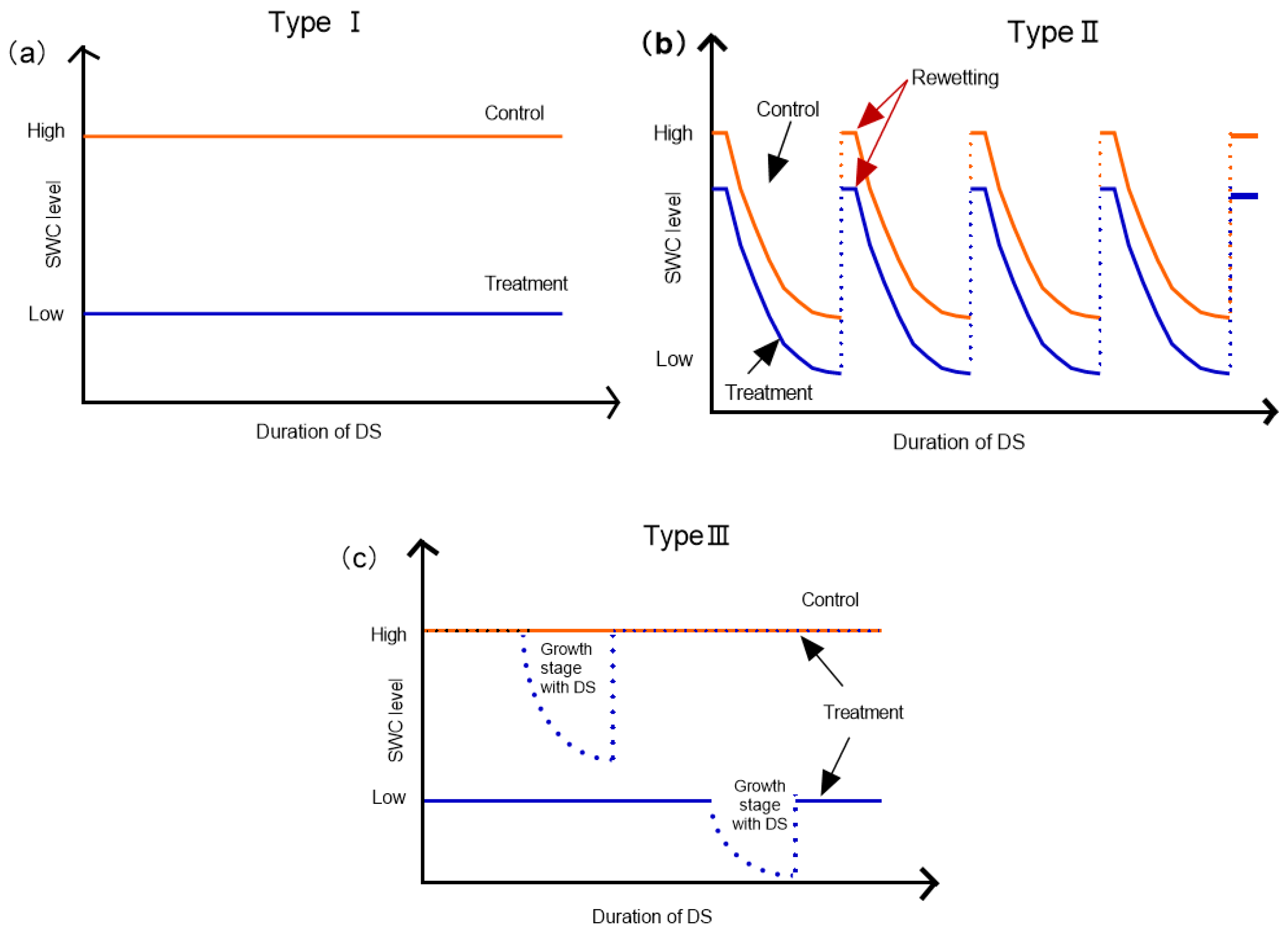
2.2. Differences of DS Type and Manipulation Time on AM and ECM Plant N
2.3. Differences of DS Type and Manipulation Time on AM and ECM Plant P
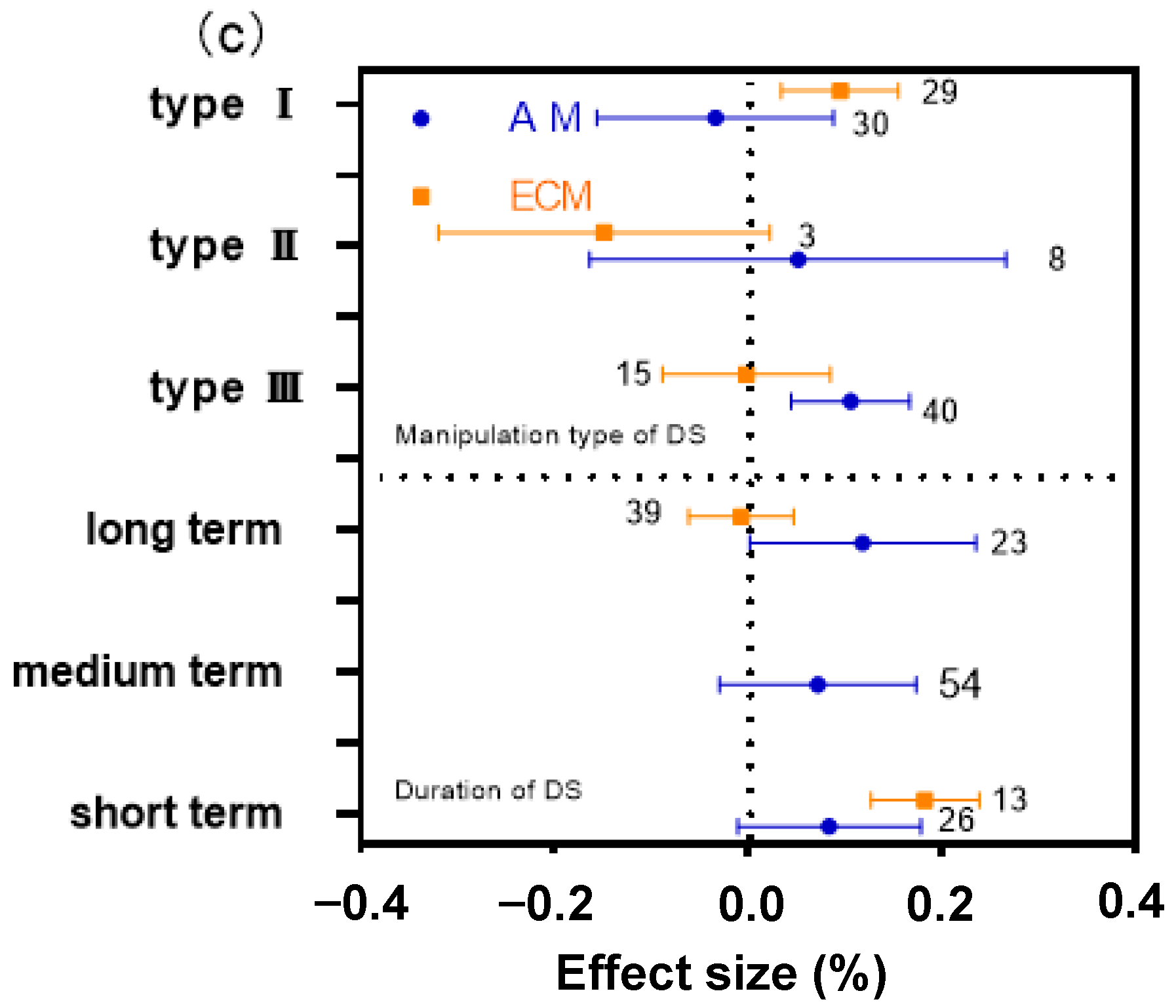
2.4. The Difference of DS Type and Manipulation Time on AM and ECM Plant N:P Ratio
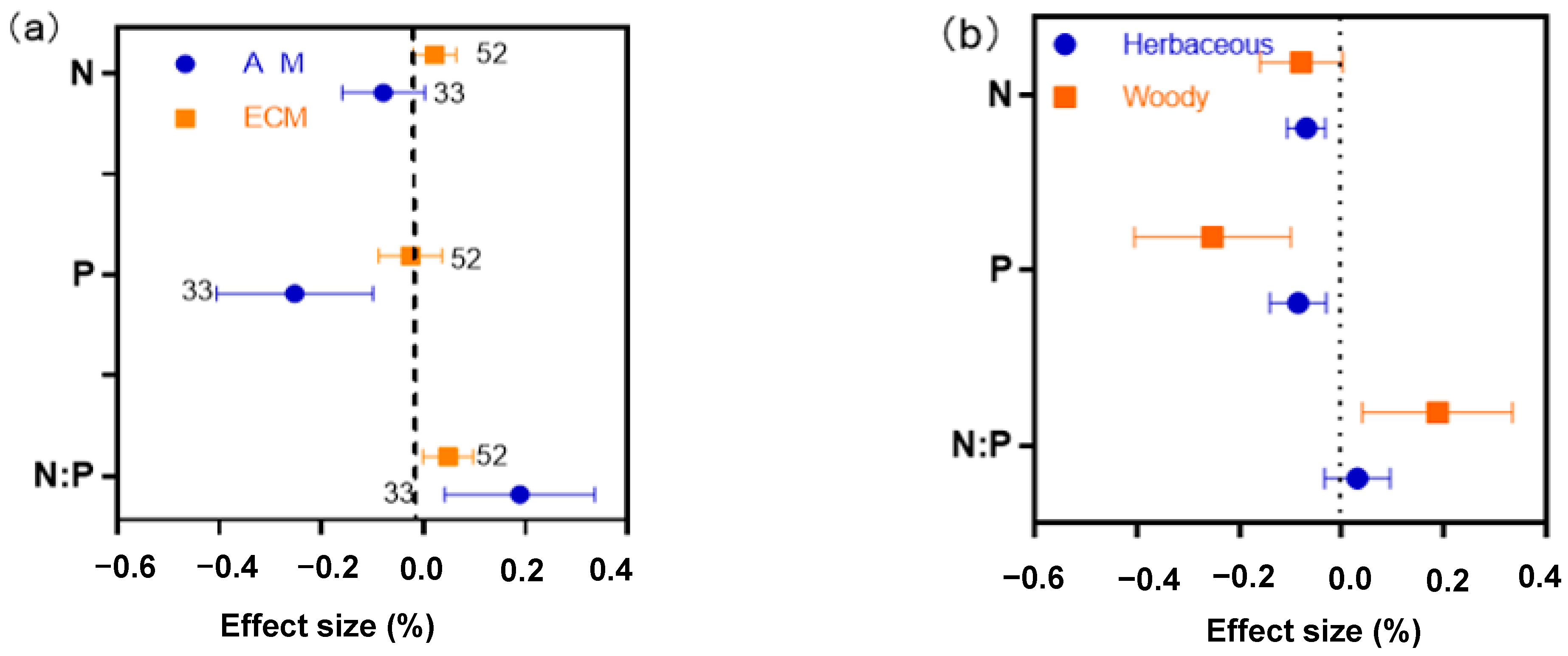
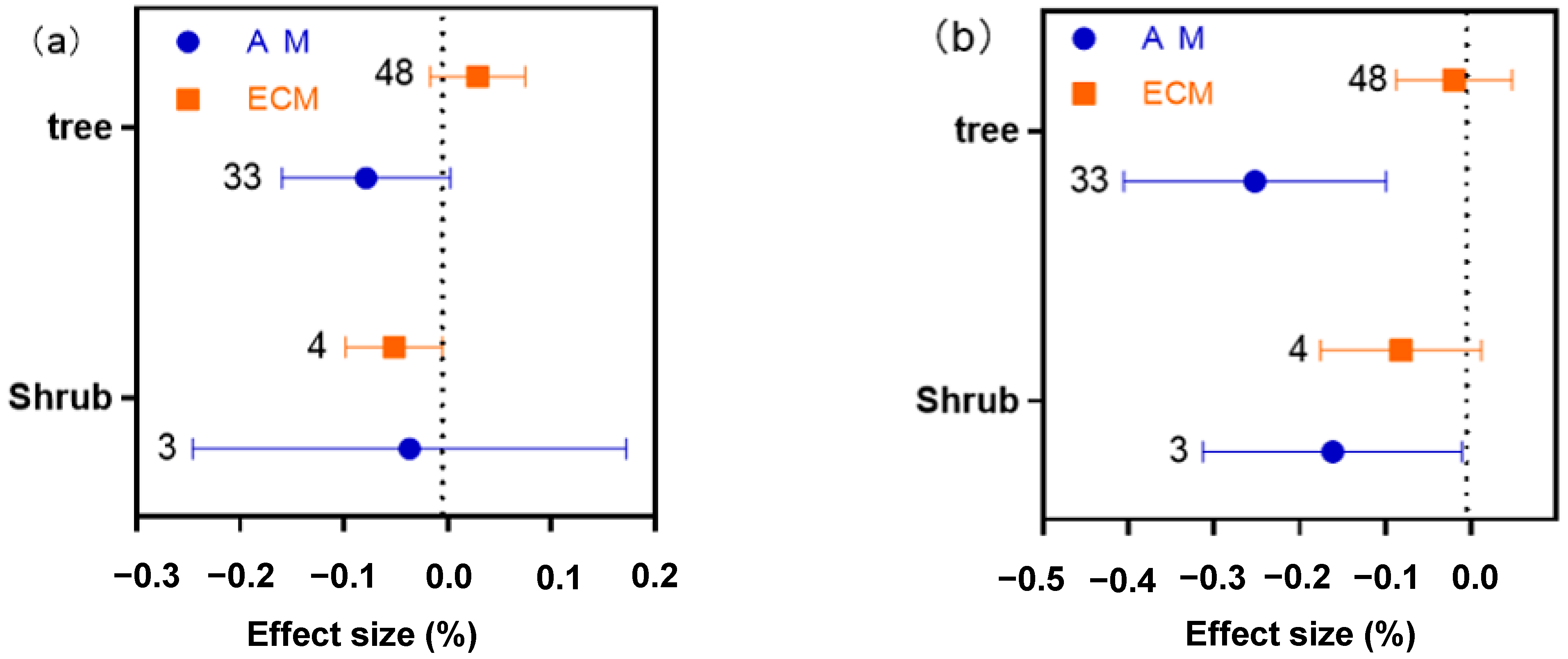
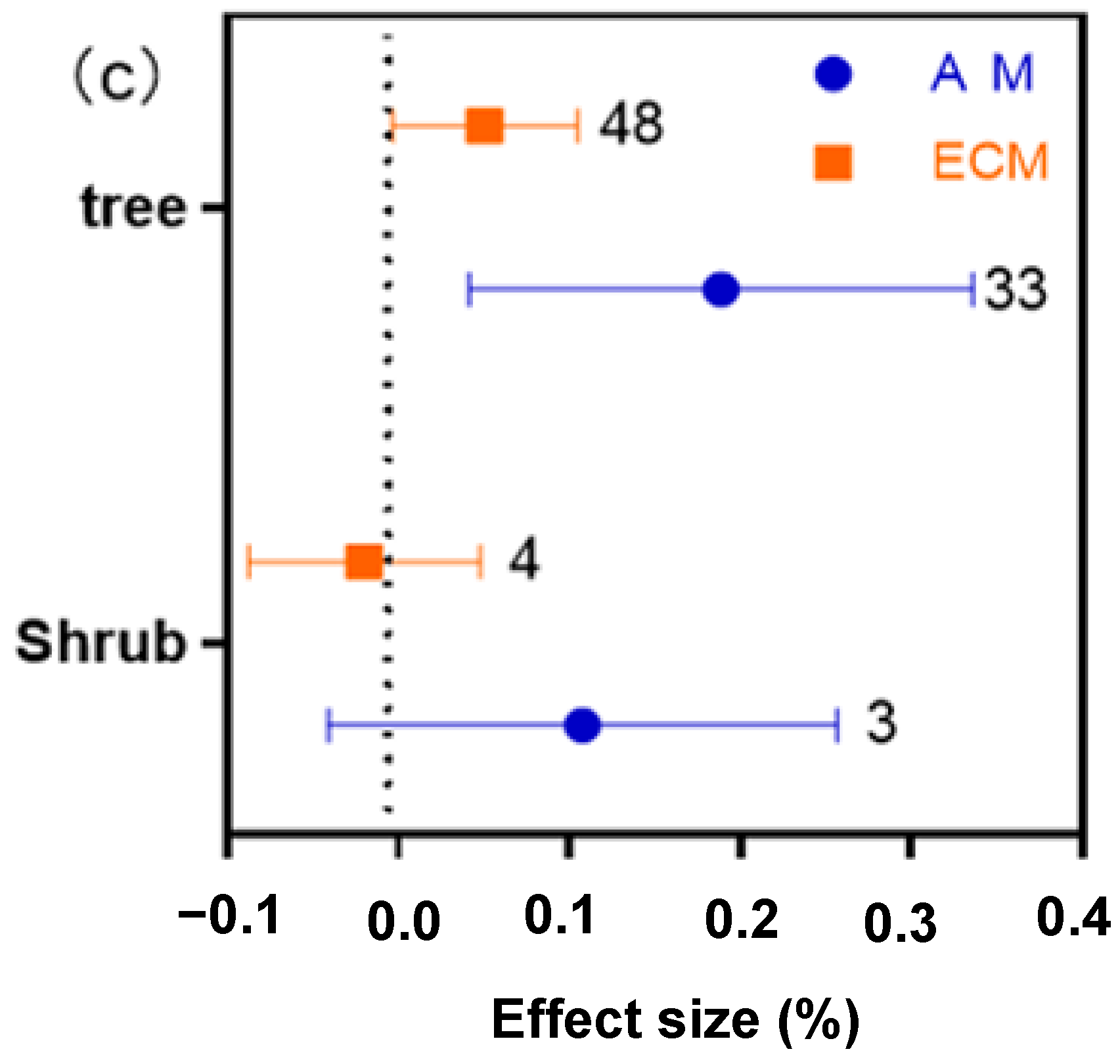
2.5. Different Functional Groups of AM and ECM Showed Different Responses of N, P and N:P Ratio to Drought
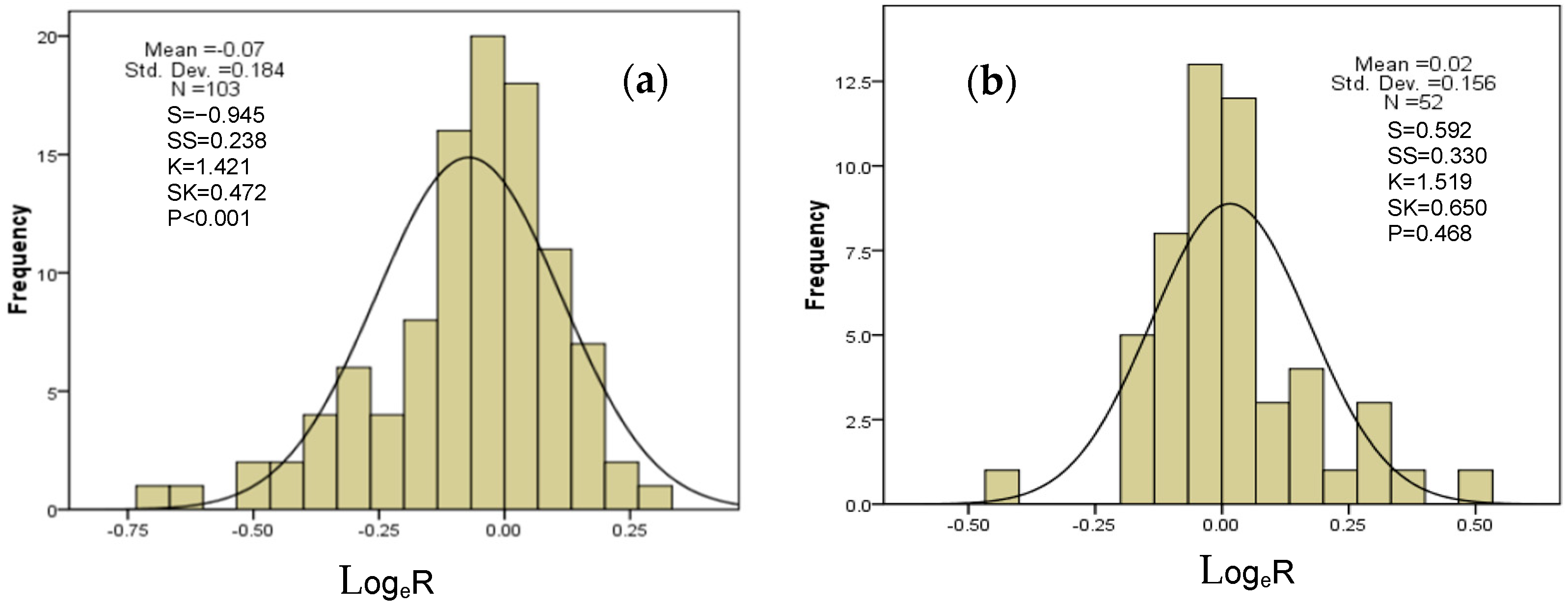
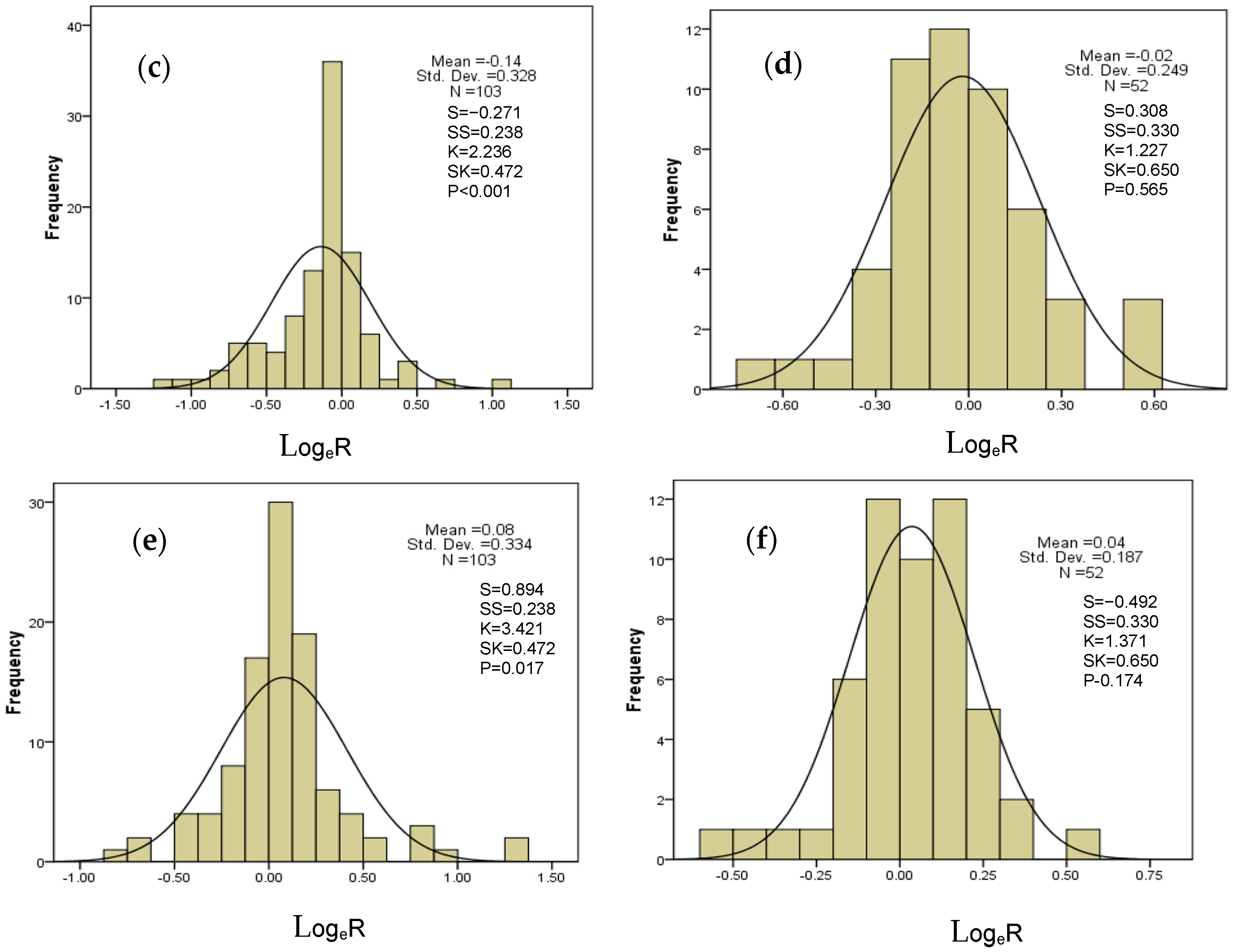
2.6. Overall Difference and Distribution Range of N, P, and N:P Ratio Effects of Drought on AM and ECM Plants
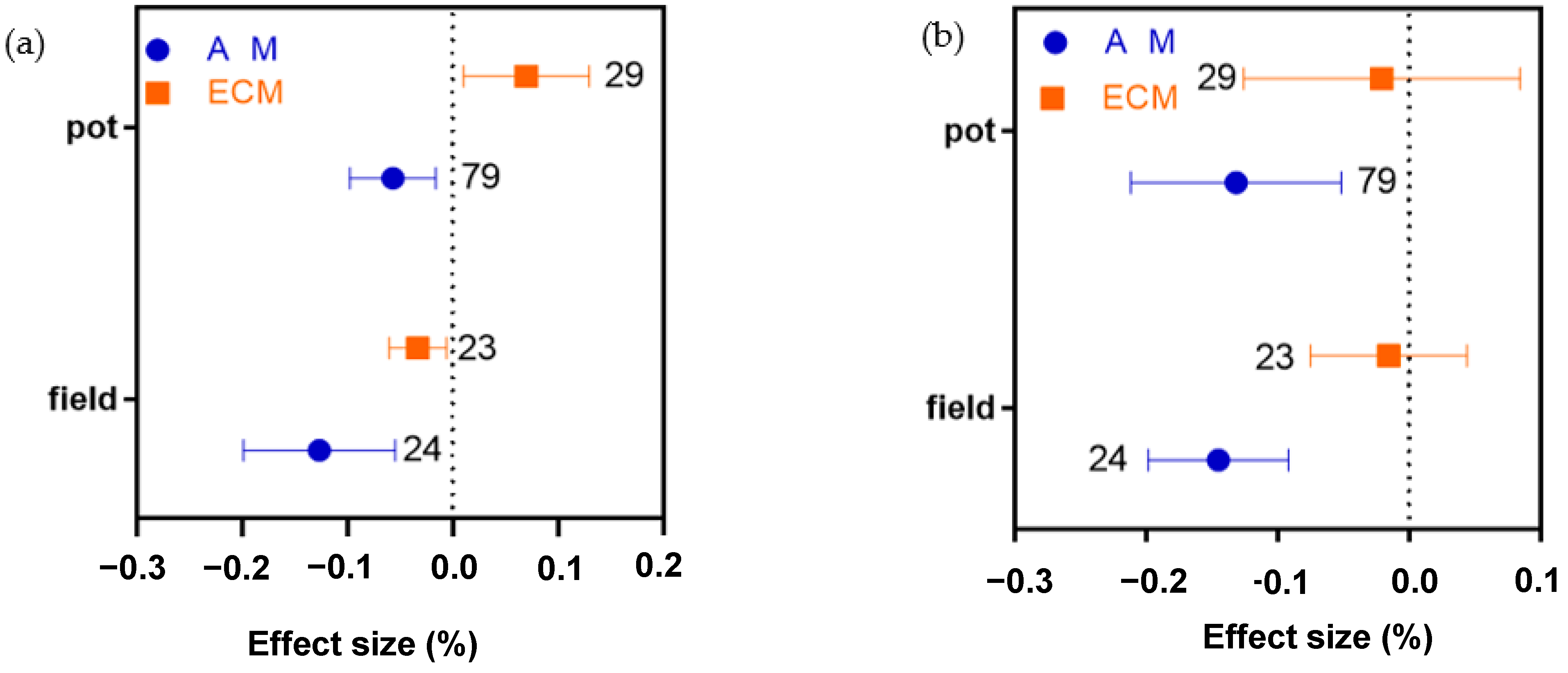
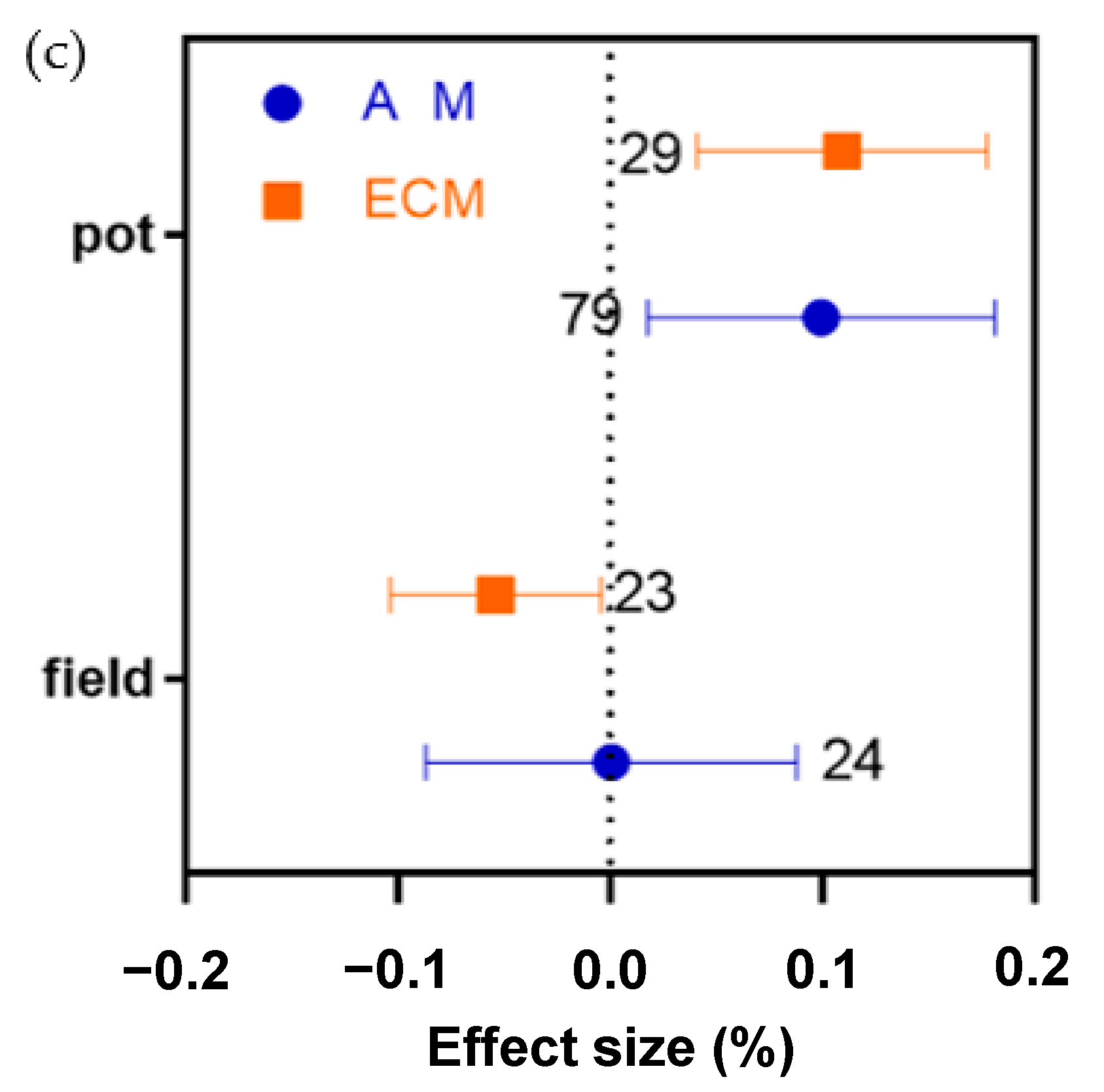
2.7. Different Effects of Pot or Field Treatment on AM and ECM Plant N, P, and N:P Ratio
2.8. Effects of Other Factors on N, P, and N:P Ratio of AM and ECM Plants
3. Materials and Methods
3.1. Data Compilation
3.2. Data Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: London, UK, 2008. [Google Scholar]
- Genre, A.; Lanfranco, L.; Perotto, S.; Bonfante, P. Unique and common traits in mycorrhizal symbioses. Nat. Rev. Microbiol. 2020, 18, 649–660. [Google Scholar] [CrossRef] [PubMed]
- Van der Heijden, M.G.A.; Martin, F.M.; Selosse, M.A.; Sanders, I.R. Mycorrhizal ecology and evolution: The past, the present, and the future. New Phytol. 2015, 205, 1406–1423. [Google Scholar] [CrossRef]
- Qiu, Q.; Bender, S.; Franz, M.A.S.; Hu, Y.L. Arbuscular mycorrhizal fungi mitigate soil nitrogen and phosphorus losses: A meta-analysis. Sci. Total Environ. 2022, 807, 150–857. [Google Scholar] [CrossRef] [PubMed]
- Parniske, M. Arbuscular mycorrhiza: The mother of plant root endosymbiosis. Nat. Rev. Microbiol. 2008, 6, 763–775. [Google Scholar] [CrossRef]
- Hodge, A.; Campbell, C.D.; Fitter, A.H. An arbuscular mycorrhizal fungus accelerates decomposition and acquires nitrogen directly from organic material. Nature 2001, 413, 297–299. [Google Scholar] [CrossRef]
- Wu, Q.S.; Cao, M.Q.; Zou, Y.N.; Wu, C.; He, X.H. Mycorrhizal colonization represents functional equilibrium on root morphology and carbon distribution of trifoliate orange grown in a split-root system. Sci. Hortic. 2016, 199, 95–102. [Google Scholar]
- Chen, L.; Cai, Y.X.; Lei, H.M.; Qi, X.X.; Lin, J.J.; Liao, W.; Huang, Z.X. Comparison of soil nitrogen cycling between arbuscular mycorrhizal and ectomycorrhizal forests in a subtropical region. Chin. J. Ecol. 2022, 41, 218–226. [Google Scholar]
- Liu, X.Y.; Hu, Y.K. Carbon, nitrogen and phosphorus stoichiometry in leaves and fine roots of typical forest swamps in the Greater Khingan Mountains. Chin. J. Appl. Ecol. 2020, 31, 3385–3394. [Google Scholar]
- Shi, Z.Y.; Xu, S.X.; Yang, M.; Zhang, M.G.; Lu, S.H.; Chang, H.Q.; Wang, X.G.; Chen, X.N. Leaf nitrogen and phosphorus stoichiometry are closely linked with mycorrhizal type traits of legume species. Legume Res. Int. J. 2021, 44, 81–87. [Google Scholar] [CrossRef]
- Cheeke, T.E.; Phillips, R.P.; Brzostek, E.R.; Rosling, A.; Bever, J.D.; Fransson, P. Dominant mycorrhizal association of trees alters carbon and nutrient cycling by selecting for microbial groups with distinct enzyme function. New Phytol. 2017, 214, 432–442. [Google Scholar] [CrossRef]
- Wang, J.L.; Chen, G.F.; Zou, G.Y.; Song, X.F.; Liu, F.X. Comparative on plant stoichiometry response to agricultural non-point source pollution in different types of ecological ditches. Environ. Sci. Pollut. Res. 2019, 26, 647–658. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.F.; Li, Q.; Lei, Z.F.; Zhang, J.B.; Song, X.Z.; Song, X.Z. Ecological stoichiometry of nitrogen and phosphorus in Moso bamboo (Phyllostachys edulis) during the explosive growth period of new emergent shoots. J. Plant Res. 2019, 132, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Lebauer, D.S.; Treseder, K.K. Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed. Ecology 2008, 89, 371–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, Y.; Yi, S.; Ren, S.; Li, N.; Chen, J. Responses of typical grasslands in a semi-arid basin on the Qinghai-Tibetan Plateau to climate change and disturbances. Environ. Earth Sci. 2014, 71, 1421–1431. [Google Scholar] [CrossRef]
- Sistla, S.A.; Schimel, J.P. Stoichiometric flexibility as a regulator of carbon and nutrient cycling in terrestrial ecosystems under change. New Phytol. 2012, 196, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Sterner, R.W.; Elser, J.J. Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere; Princeton University Press: Princeton, NJ, USA, 2002. [Google Scholar]
- Reich, P.B.; Oleksyn, J. Global patterns of plant leaf N and P in relation to temperature and latitude. Proc. Natl. Acad. Sci. USA 2004, 101, 11001–11006. [Google Scholar] [CrossRef]
- Guo, L.L.; Chen, Y.; Zhang, Z.; Fukushima, T. N: P stoichiometry in a forested runoff during storm events: Comparisons with regions and vegetation types. Sci. World J. 2012, 2012, 257392. [Google Scholar] [CrossRef] [PubMed]
- Asrar, A.A.; Abdel-Fattah, G.M.; Elhindi, K.M. Improving growth, flower yield, and water relations of snapdragon (Antirhinum majus L.) plants grown under well-watered and water-stress conditions using arbuscular mycorrhizal fungi. Photosynthetica 2012, 50, 305–316. [Google Scholar] [CrossRef]
- Manoharan, P.T.; Shanmugaiah, V.; Balasubramanian, N.; Gomathinayagam, S.; Muthuchelian, K. Influence of AM fungi on the growth and physiological status of Erythrina variegata Linn. grown under different water stress conditions. Eur. J. Soil Biol. 2010, 46, 151–156. [Google Scholar] [CrossRef]
- Farooq, M.; Hussain, M.; Wahid, A.; Siddique, K.H.M. Drought stress in plants: An overview. In Plant Responses to Drought Stress; Aroca, R., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 1–33. [Google Scholar]
- Zeng, D.P.; Jiang, L.L.; Zeng, C.S. Reviews on the ecological stoichiometry characteristics and its applications. Acta Ecol. Sin. 2013, 33, 54–84. [Google Scholar]
- He, M.Z.; Dijkstra, F.A. Drought effect on plant nitrogen and phosphorus: A meta-analysis. New Phytol. 2014, 204, 924–931. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.C.; Song, F.B.; Liu, S.Q.; Liu, T.D.; Zhou, X. Arbuscular mycorrhizae improves photosynthesis and water status of Zea mays L. under drought stress. Plant Soil Environ. 2012, 64, 186–191. [Google Scholar] [CrossRef]
- Augé, R.M. Arbuscular mycorrhizae and soil/plant water relations. Can. J. Soil Sci. 2004, 84, 373–381. [Google Scholar] [CrossRef]
- Lehto, T.; Zwiazek, J.J. Ectomycorrhizas and water relations of trees: A review. Mycorrhiza 2011, 21, 71–90. [Google Scholar] [CrossRef]
- Xu, H.; Minna, K.; Walid, E.K.; Lee, S.H.; Pardo, A.G.; Cooke, J.E.K.; Zwiazek, J.J. Overexpression of Laccaria bicolor aquaporin JQ585595 alters root water transport properties in ectomycorrhizal white spruce (Picea glauca) seedlings. New Phytol. 2015, 205, 757–770. [Google Scholar] [CrossRef]
- Mónica, S.; Bernardes, A.D.S.; Rita, M.A.; André, A.; Susana, S.; Rui, M. Ectomycorrhizal inoculation with Pisolithus tinctorius reduces stress induced by drought in cork oak. Mycorrhiza 2018, 28, 247–258. [Google Scholar]
- Lin, S.S.; Sun, X.W.; Wang, X.J.; Dou, C.Y.; Li, Y.Y.; Luo, Q.Y.; Sun, L.; Jin, L. Progress and application prospect of mycorrhizal studies in China. Acta Prataculturae Sin. 2013, 22, 310–325. [Google Scholar]
- Hempel, S.; Götzenberger, L.; Kühn, I.; Michalski Stefan, G.; Rillig Matthias, C.; Zobel, M.; Moora, M. Mycorrhizas in the central European flora: Relationships with plant life history traits and ecology. Ecology 2013, 94, 1389–1399. [Google Scholar] [CrossRef]
- Wang, B.; Qiu, Y.L. Phylogenetic distribution and evolution of mycorrhizas in land plants. Mycorrhiza 2006, 16, 299–363. [Google Scholar] [CrossRef]
- Harley, J.L.; Harley, E.L. A check-list of mycorrhiza in the British. New Phytol. 1987, 107, 741–749. [Google Scholar] [CrossRef]
- Averill, C.; Bhatnagar, J.M.; Dietze, M.C.; Pearse, W.D.; Kivlin, S.N. Global imprint of mycorrhizal fungi on whole-plant nutrient economics. Proc. Natl. Acad. Sci. USA 2019, 116, 23163–23168. [Google Scholar] [CrossRef]
- Zhang, H.Y.; Lu, X.T.; Hartmann, H.; Kelle, A.; Han, X.G.; Trumbore, S.; Phillips, R. Foliar nutrient resorption differs between arbuscular mycorrhizal and ectomycorrhizal trees at local and global scales. Glob. Ecol. Biogeogr. 2018, 27, 875–885. [Google Scholar] [CrossRef]
- Cornelissen, J.H.C.; Aerts, R.; Cerabolini, B.; Werger, M.; van der Heijden, M. Carbon cycling traits of plant species are linked with mycorrhizal strategy. Oecologia 2001, 129, 611–619. [Google Scholar] [CrossRef] [PubMed]
- Read, D.J. Mycorrhizas in ecosystems. Experientia 1991, 47, 376–391. [Google Scholar] [CrossRef]
- Schulze, E.-D.; Chapin, F.S.; Gebauer, G. Nitrogen nutrition and isotope differences among life forms at the northern tree line of Alaska. Oecologia 1994, 100, 406–412. [Google Scholar] [CrossRef] [PubMed]
- Chapin, F.S. New cog in the nitrogen cycle. Nature 1995, 377, 199–200. [Google Scholar] [CrossRef]
- Michelsen, A.; Quarmby, C.; Sleep, D.; Jonasson, S. Vascular plant 15N natural abundance in heath and forest tundra ecosystems is closely correlated with presence and type of mycorrhizal fungi in roots. Oecologia 1998, 115, 406–418. [Google Scholar] [CrossRef]
- Näsholm, T.; Ekblad, A.; Nordin, A.; Giesler, R.; Högberg, M.; Högberg, P. Boreal forest plants take up organic nitrogen. Nature 1998, 392, 914–916. [Google Scholar] [CrossRef]
- Yan, G.Y.; Lu, J.; Qiu, L.Y.; Huang, M.T.; Xing, Y.J.; Wang, Q.G. Mycorrhizal characteristics of AM and ECM and their response to environmental changes. J. Qufu Normal Univ. 2022, 48, 106–112. [Google Scholar] [CrossRef]
- Jackson, R.B.; Canadell, J.; Ehleringer, J.R.; Mooney, H.A.; Sala, O.E.; Schulze, E.D. A global analysis of root distributions for terrestrial biomes. Oecologia 1996, 108, 389–411. [Google Scholar] [CrossRef]
- Deng, L.; Peng, C.H.; Kim, D.G.; Li, J.W.; Liu, Y.l.; Hai, X.Y.; Liu, Q.Y.; Huang, C.B.; Shangguan, Z.P.; Kuzyakov, Y.K. Drought effects on soil carbon and nitrogen dynamics in global natural ecosystems: A meta-analysis. Earth-Sci. Rev. 2020, 214, 103501. [Google Scholar] [CrossRef]
- Tian, D.; Yan, Z.B.; Fang, J.Y. Plant ecological stoichiometry and its main hypotheses. Chin. J. Plant Ecol. 2021, 45, 682–713. [Google Scholar] [CrossRef]
- Fu, W.; Chen, B.D.; Rillig, M.C.; Jansa, J.; Ma, W.; Xu, C.; Luo, W.T.; Wu, H.H.; Hao, Z.P.; Wu, H.; et al. Community response of arbuscular mycorrhizal fungi to extreme drought in a cold-temperate grassland. New Phytol. 2021, 234, 2003–2017. [Google Scholar] [CrossRef] [PubMed]
- Kilpelainen, J.; Vestberg, M.; Repo, T.; Lehto, T. Arbuscular and ectomycorrhizal root colonisation and plant nutrition in soils exposed to freezing temperatures. Soil Biol. Biochem. 2016, 99, 85–93. [Google Scholar] [CrossRef]
- Kilpelainen, J.; Barbero-Lopez, A.; Vestberg, M.; Heiskanen, J.; Lehto, T. Does severe soil drought have after-effects on arbuscular and ectomycorrhizal root colonisation and plant nutrition? Plant Soil. 2017, 418, 377–386. [Google Scholar] [CrossRef]
- Fernández, R.; Bertrand, A.; Casares, A.; García, R.; González, A.; Tamés, R.S. Cadmium accumulation and its effect on the in vitro growth of woody fleabane and mycorrhized white birch. Environ pollu. 2008, 152. [Google Scholar] [CrossRef]
- Baar, J.; Bastiaans, T.; van de Coevering, M.A.; Roelofs, J.G.M. Ectomycorrhizal root development in wet Alder carr forests in response to desiccation and eutrophication. Mycorrhiza 2002, 12. [Google Scholar]
- Zhang, Y.M.; Ma, K.M.; Qu, L.Y. Effects of AM fungus inoculation on the root system of small saddle sheep hoof beetle seedlings under drought conditions. Acta Ecologica Sinica. 2017, 37, 2611–2619. [Google Scholar]
- Chaiane, S.; Juarês José, A.; Sidney, L.S. Efficiency of the on-farm Mycorrhizal inoculant and phonolite rock on growth and nutrition of schinus terebinthifolius and eucalyptus saligna. Rev Bras Cienc. 2017, 40. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Planting Conditions (PC) | Species Distribution (SD) | PC × SD | ||||
|---|---|---|---|---|---|---|
| F | P | F | P | F | P | |
| N | 6.754 | 0.01 | 3.143 | 0.078 | 1.366 | 0.244 |
| P | 0 | 0.997 | 0.733 | 0.394 | 0.065 | 0.799 |
| N:P | 2.375 | 0.126 | 0.134 | 0.716 | 0.662 | 0.422 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jing, M.; Shi, Z.; Zhang, M.; Zhang, M.; Wang, X. Nitrogen and Phosphorus of Plants Associated with Arbuscular and Ectomycorrhizas Are Differentially Influenced by Drought. Plants 2022, 11, 2429. https://doi.org/10.3390/plants11182429
Jing M, Shi Z, Zhang M, Zhang M, Wang X. Nitrogen and Phosphorus of Plants Associated with Arbuscular and Ectomycorrhizas Are Differentially Influenced by Drought. Plants. 2022; 11(18):2429. https://doi.org/10.3390/plants11182429
Chicago/Turabian StyleJing, Manman, Zhaoyong Shi, Mengge Zhang, Menghan Zhang, and Xiaohui Wang. 2022. "Nitrogen and Phosphorus of Plants Associated with Arbuscular and Ectomycorrhizas Are Differentially Influenced by Drought" Plants 11, no. 18: 2429. https://doi.org/10.3390/plants11182429