
Comparative Metabolomics Analysis of Stigmas and Petals in Chinese Saffron (Crocus sativus) by Widely Targeted Metabolomics


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Widely Targeted Metabolomics Analysis in Stigmas and Petals
2.2. Multivariate Analysis of Differential Metabolites among Stigmas and Petals
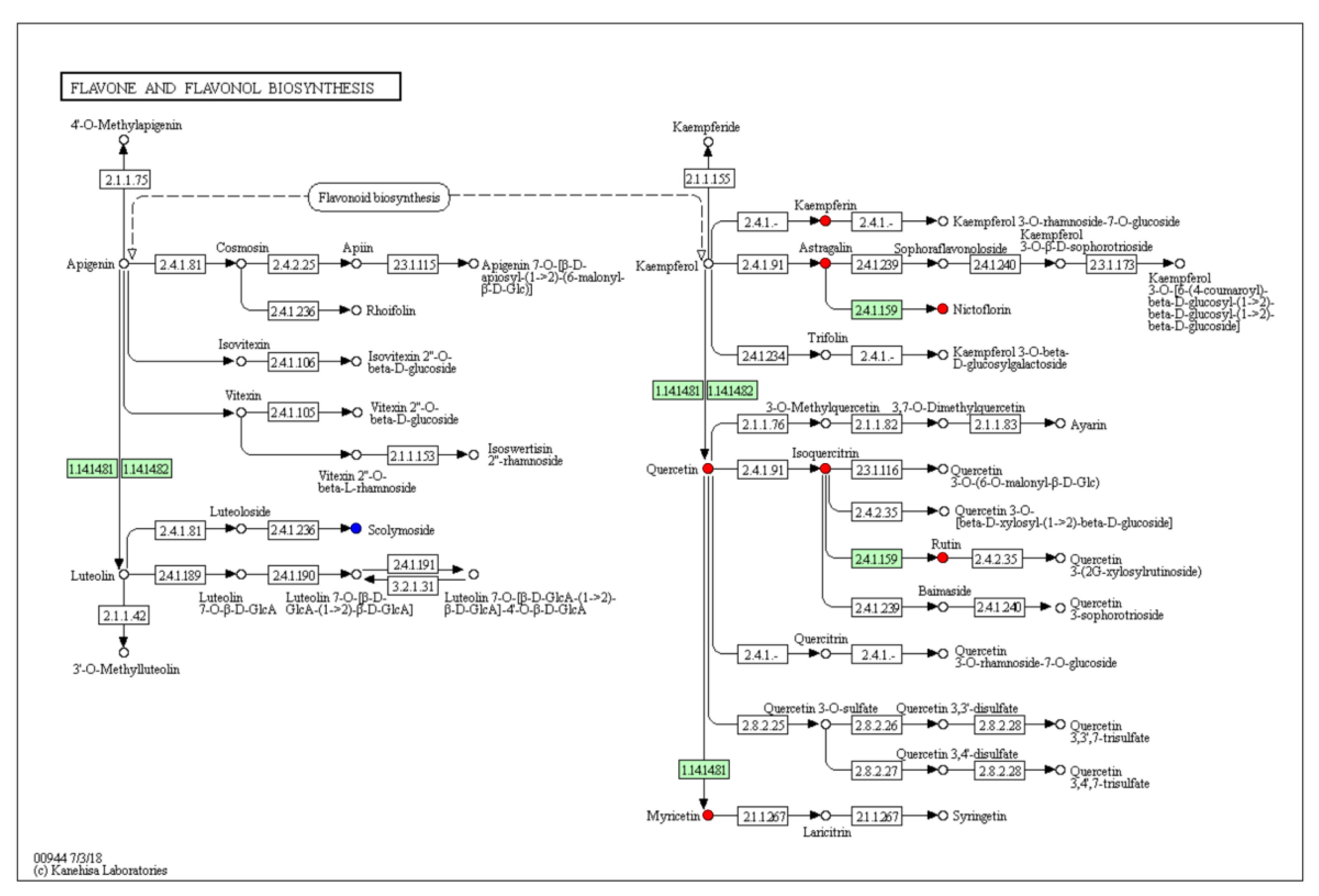
2.3. Metabolic Pathway Analysis of Differential Metabolites
3. Discussion
4. Materials and Methods
4.1. Site Description
4.2. Sample Preparation and Metabolite Extraction
4.3. UHPLC-MS Analysis
4.4. Data Preprocessing and Annotation
4.5. Statistical Analysis
4.6. KEGG Function Annotation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ahrazem, O.; Rubio-Moraga, A.; Nebauer, S.G.; Molina, R.V.; Gómez-Gómez, L. Saffron: Its phytochemistry, developmental processes, and biotechnological prospects. J. Agric. Food Chem. 2015, 63, 8751–8764. [Google Scholar] [CrossRef] [PubMed]
- Gresta, F.; Lombardo, G.M.; Siracusa, L.; Ruberto, G. Saffron, an alternative crop for sustainable agricultural systems. A review. Agron. Sustain. Dev. 2008, 28, 95–112. [Google Scholar] [CrossRef]
- Zeka, K.; Marrazzo, P.; Micucci, M.; Ruparelia, K.C.; Arroo, R.R.J.; Macciarelli, G.; Nottola, S.A.; Continenza, M.A.; Chiarini, A.; Angeloni, C.; et al. Activity of antioxidants from Crocus sativus L. petals: Potential preventive effects towards cardiovascular system. Antioxidants 2020, 9, 1102. [Google Scholar] [CrossRef] [PubMed]
- Yao, C.; Qian, X.D.; Zhou, G.F.; Zhang, S.W.; Li, L.Q.; Guo, Q.S. A comprehensive analysis and comparison between vacuum and electric oven drying methods on Chinese saffron (Crocus sativus L.). Food Sci. Biotechnol. 2019, 28, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Lambrianidou, A.; Koutsougianni, F.; Papapostolou, I.; Dimas, K. Recent advances on the anticancer properties of saffron (Crocus sativus L.) and its major constituents. Molecules 2021, 26, 86. [Google Scholar] [CrossRef] [PubMed]
- Ganaie, S.A.; Dwivedi, S.; Bhat, A.; Khanday, M. A study on economics of saffron (Crocus sativus L.) cultivation in Jammu and Kashmir, India. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 1600–1616. [Google Scholar]
- Shahnoushi, N.; Abolhassani, L.; Kavakebi, V.; Reed, M.; Saghaian, S. Chapter 21—Economic analysis of saffron production. In Woodhead Publishing Series in Food Science, Technology and Nutrition, Saffron; Woodhead Publishing: Cambridge, UK, 2020; pp. 337–356. [Google Scholar]
- Hashemi Gahruie, H.; Parastouei, K.; Mokhtarian, M.; Rostami, H.; Niakousari, M.; Mohsenpour, Z. Application of innovative processing methods for the extraction of bioactive compounds from saffron (Crocus sativus) petals. J. Appl. Res. Med. Aromat. Plants 2020, 19, 100264. [Google Scholar] [CrossRef]
- Mahaqiq, Z.; Moossavi, M.; Hemmati, M.; Kazemi, T.; Mehrpour, O. Antioxidant properties of saffron stigma and petals: A potential therapeutic approach for insulin resistance through an insulin-sensitizing adipocytokine in high-calorie diet rats. Int. J. Prev. Med. 2020, 11, 184. [Google Scholar]
- Moshfegh, F.; Balanejad, S.Z.; Shahrokhabady, K.; Attaranzadeh, A. Crocus sativus (saffron) petals extract and its active ingredient, anthocyanin improves ovarian dysfunction, regulation of inflammatory genes and antioxidant factors in testosterone-induced PCOS mice. J. Ethnopharmacol. 2021, 282, 114594. [Google Scholar] [CrossRef]
- Mottaghipisheh, J.; Sourestani, M.M.; Kiss, T.; Horváth, A.; Tóth, B.; Ayanmanesh, M.; Khamushi, A.; Csupor, D. Comprehensive chemotaxonomic analysis of saffron crocus tepal and stamen samples, as raw materials with potential antidepressant activity. J. Pharm. Biomed. Anal. 2020, 184, 113183. [Google Scholar] [CrossRef]
- Xu, S.Y.; Ge, X.J.; Li, S.M.; Guo, X.Y.; Dai, D.D.; Yang, T.G. Discrimination of Different Parts of Saffron by Metabolomic-Based Ultra-Performance Liquid Chromatography Coupled with High-Definition Mass Spectrometry. Chem. Biodivers. 2019, 16, e1900363. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, R.; Ahmed, H.; Dixit, R.K.; Dharamveer; Saraf, S.A. Crocus sativus L.: A comprehensive review. Pharmacogn. Rev. 2010, 4, 200–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vahedi, M.; Kabiri, M.; Salami, S.A.; Rezadoost, H.; Kanani, M.R. Quantitative HPLC-based metabolomics of some Iranian saffron (Crocus sativus L.) accessions. Ind. Crops Prod. 2018, 118, 26–29. [Google Scholar] [CrossRef]
- Fang, Q.W.; Fu, W.W.; Yang, J.L.; Lu, Y.; Chen, J.C.; Wu, P.Z.; Zhang, X.; Xu, H.X. New monoterpenoids from the stigmas of Crocus sativus. J. Nat. Med. 2022, 76, 102–109. [Google Scholar] [CrossRef]
- Goli, S.A.H.; Mokhtari, F.; Rahimmalek, M. Phenolic compounds and antioxidant activity from saffron (Crocus sativus L.) petal. J. Agric. Sci. 2012, 4, 175–181. [Google Scholar]
- Gajula, S.N.R.; Nanjappan, S. Metabolomics: A recent advanced omics technology in herbal medicine research. In Medicinal and Aromatic Plants; Academic Press: New York, NY, USA, 2021; pp. 97–117. [Google Scholar]
- Li, F.M.; Lu, X.M.; Liu, H.P.; Liu, M.; Xiong, Z.L. A pharmaco-metabonomic study on the therapeutic basis and metabolic effects of Epimedium brevicornum Maxim. on hydrocortisone-induced rat using UPLC-MS. Biomed. Chromatogr. 2007, 21, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Hou, C.C.; Chen, C.H.; Yang, N.S.; Chen, Y.P.; Lo, C.P.; Wang, S.Y.; Tien, Y.J.; Tsai, P.W.; Shyur, L.F. Comparative metabolomics approach coupled with cell- and gene-based assays for species classification and anti-inflammatory bioactivity validation of Echinacea plants. J. Nutr. Biochem. 2010, 21, 1045–1059. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Bai, X.; Yang, F.Q.; Zhang, X.J.; Hu, Y.J.; Li, P.; Wan, J.B. Discriminating from species of Curcumae Radix (Yujin) by a UHPLC/Q-TOFMS-based metabolomics approach. Chin. Med. 2016, 11, 21. [Google Scholar] [CrossRef]
- Wu, J.; Yang, W.Z.; Pan, H.Q.; Yao, S.; Wu, W.Y.; Guo, D.A. Geographic impact evaluation of the quality of Alismatis Rhizoma by untargeted metabolomics and quantitative assay. J. Sep. Sci. 2017, 41, 839–846. [Google Scholar] [CrossRef]
- Yu, C.N.; Luo, X.J.; Zhan, X.R.; Hao, J.; Zhang, L.; Song, Y.B.L.; Shen, C.J.; Dong, M. Comparative metabolomics reveals the metabolic variations between two endangered Taxus species (T. fuana and T. yunnanensis) in the Himalayas. BMC Plant Biol. 2018, 18, 197. [Google Scholar] [CrossRef]
- Shang, X.H.; Huang, D.; Wang, Y.; Xiao, L.; Ming, R.H.; Zeng, W.D.; Cao, S.; Lu, L.Y.; Wu, Z.D.; Yan, H.B. Identification of nutritional ingredients and medicinal components of Pueraria lobata and its varieties using UPLC-MS/MS-based metabolomics. Molecules 2021, 26, 6587. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, A.; Razavi, B.M.; Hosseinzadeh, H. Saffron (Crocus sativus) petal as a new pharmacological target: A review. Iran. J. Basic Med. Sci. 2018, 21, 1091–1099. [Google Scholar] [PubMed]
- Qin, X.X.; Lu, Y.F.; Zhen, P.; Fan, S.X.; Yao, Y.C. Systematic chemical analysis approach reveals superior antioxidant capacity via the synergistic effect of flavonoid compounds in red vegetative tissues. Front. Chem. 2018, 6, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brito, S.A.; Rodrigues, F.F.G.; Campos, A.R.; da Costa, J.G.M. Evaluation of the antifungal activity and modulation between Cajanus cajan (L.) Millsp. leaves and roots ethanolic extracts and conventional antifungals. Pharmacogn. Mag. 2012, 8, 103–106. [Google Scholar] [PubMed]
- Karak, P. Biological activities of flavonoids: An overview. Int. J. Pharm. Sci. Res. 2019, 10, 1567–1574. [Google Scholar]
- Yuan, Z.H.; Liang, Z.E.; Wu, J.; Yi, J.E.; Chen, X.J.; Sun, Z.L. A potential mechanism for the anti-apoptotic property of koumine involving mitochondrial pathway in LPS-mediated RAW 264.7 macrophages. Molecules 2016, 21, 1317. [Google Scholar] [CrossRef]
- Deryabin, D.; Inchagova, K.; Rusakova, E.; Duskaev, G. Coumarin’s anti-quorum sensing activity can be enhanced when combined with other plant-derived small molecules. Molecules 2021, 26, 208. [Google Scholar] [CrossRef]
- You, M.K.; Lee, Y.J.; Kim, J.K.; Baek, S.A.; Jeon, Y.A.; Lim, S.H.; Ha, S.H. The organ-specific differential roles of rice DXS and DXR, the first two enzymes of the MEP pathway, in carotenoid metabolism in Oryza sativa leaves and seeds. BMC Plant Biol. 2020, 20, 167. [Google Scholar] [CrossRef]
- Soliman, S.M.; Sheta, N.M.; Ibrahim, B.M.M.; El-Shawwa, M.M.; El-Halim, S.M.A. Novel intranasal drug delivery: Geraniol charged polymeric mixed micelles for targeting cerebral insult as a result of ischaemia/reperfusion. Pharmaceutics 2020, 12, 76. [Google Scholar] [CrossRef]
- Chu, J.P.; Zhang, T.T.; He, K.X. Cariogenicity features of Streptococcus mutans in presence of rubusoside. BMC Oral Health 2016, 16, 54. [Google Scholar] [CrossRef]
- Kashchenko, N.I.; Olennikov, D.N. Phenolome of asian agrimony tea (Agrimonia asiatica Juz., Rosaceae): LC-MS profile, α-glucosidase inhibitory potential and stability. Foods 2020, 9, 1348. [Google Scholar] [CrossRef] [PubMed]
- Park, C.H.; Chae, S.C.; Park, S.Y.; Kim, J.K.; Kim, Y.J.; Chung, S.O.; Arasu, M.V.; Al-Dhabi, N.A.; Park, S.U. Anthocyanin and carotenoid contents in different cultivars of chrysanthemum (Dendranthema grandiflorum Ramat.) flower. Molecules 2015, 20, 11090–11102. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekara, A.; Daugelaite, J.; Shahidi, F. DNA scission and LDL cholesterol oxidation inhibition and antioxidant activities of Bael (Aegle marmelos) flower extracts. J. Tradit. Complement. Med. 2018, 8, 428–435. [Google Scholar] [CrossRef]
- Yoon, E.K.; Ku, S.K.; Lee, W.; Kwak, S.; Kang, H.; Jung, B.; Bae, J.S. Antitcoagulant and antiplatelet activities of scolymoside. BMB Rep. 2015, 48, 577–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, K.; Liu, Y.J.; Yang, X.Y.; Chen, X.; Ye, G. A Ginseng Saffron Liquid Beverage as well as Preparation Method. China Patent Application CN109619333A, 16 April 2019. [Google Scholar]
- Li, K.; Shang, X.H.; Zhao, Y.H.; An, H.T.; Ye, G.; Liu, Y.J. Plant Beverage Containing Saffron as well as Preparation Method and Application of Plant Beverage. China Patent Application CN106923115A, 10 December 2018. [Google Scholar]
- Wang, H.; He, S.X.; Xiong, F.; Hu, X.L.; Wang, R. A Flower Tea with Antidepressant Effect as well as Preparation Method. China Patent Application CN112438334A, 5 March 2021. [Google Scholar]
- Maeda, J.; Roybal, E.J.; Brents, C.A.; Uesaka, M.; Aizawa, Y.; Kato, T.A. Natural and glucosyl flavonoids inhibit poly(ADP-ribose) polymerase activity and induce synthetic lethality in BRCA mutant cells. Oncol. Rep. 2014, 31, 551–556. [Google Scholar] [CrossRef]
- Fatehi, M.; Rashidabady, T.; Hassanabad, Z.F.; Saffron, E. Effects of Crocus sativus petals’ extract on rat blood pressure and on responses induced by electrical field stimulation in the rat isolated vas deferens and guinea-pig ileum. J. Ethnopharmacol. 2003, 84, 199–203. [Google Scholar] [CrossRef]
- Kuhl, C.; Tautenhahn, R.; Bottcher, C.; Larson, T.R.; Neumann, S. CAMERA: An integrated strategy for compound spectra extraction and annotation of liquid chromatography/mass spectrometry data sets. Anal. Chem. 2012, 84, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.M.; Xia, T.; Peng, Y.; Pan, M.; Zhang, M.J.; Lu, H.M.; Chen, X.Q.; Liang, Y.Z. Multiscale peak detection in wavelet space. Analyst 2015, 140, 7955–7964. [Google Scholar] [CrossRef]
- Wiklund, S.; Johansson, E.; Sjostrom, L.; Mellerowicz, E.J.; Edlund, U.; Shockcor, J.P.; Gottfries, J.; Moritz, T.; Trygg, J. Visualization of GC/TOF-MS-Based metabolomics data for identification of biochemically interesting compounds using OPLS class models. Anal. Chem. 2008, 80, 115–122. [Google Scholar] [CrossRef]
- Bai, Y.L.; Zhang, F.; Zhang, H.Y.; Xu, C.; Wu, L.; Xia, C. Follicular fluid metabolite changes in dairy cows with inactive ovary identified using untargeted metabolomics. BioMed Res. Int. 2020, 2020, 1–10. [Google Scholar] [CrossRef]
- Kolde, R. Pheatmap: Pretty Heatmaps, Version 1.0.12; 2021. Available online: https://CRAN.R-project.org/package=pheatmap (accessed on 1 June 2022).
- Ogata, H.; Goto, S.; Sato, K.; Fujibuchi, W.; Bono, H.; Kanehisa, M. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 1999, 27, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.G.; Sinelnikov, I.V.; Han, B.; Wishart, D.S. MetaboAnalyst 3.0—Making metabolomics more meaningful. Nucleic Acids Res. 2015, 43, W251–W257. [Google Scholar] [CrossRef] [PubMed] [Green Version]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, L.; Cai, Y.; Yang, L.; Zou, Z.; Zhu, J.; Zhang, Y. Comparative Metabolomics Analysis of Stigmas and Petals in Chinese Saffron (Crocus sativus) by Widely Targeted Metabolomics. Plants 2022, 11, 2427. https://doi.org/10.3390/plants11182427
Zhou L, Cai Y, Yang L, Zou Z, Zhu J, Zhang Y. Comparative Metabolomics Analysis of Stigmas and Petals in Chinese Saffron (Crocus sativus) by Widely Targeted Metabolomics. Plants. 2022; 11(18):2427. https://doi.org/10.3390/plants11182427
Chicago/Turabian StyleZhou, Lin, Youming Cai, Liuyan Yang, Zhongwei Zou, Jiao Zhu, and Yongchun Zhang. 2022. "Comparative Metabolomics Analysis of Stigmas and Petals in Chinese Saffron (Crocus sativus) by Widely Targeted Metabolomics" Plants 11, no. 18: 2427. https://doi.org/10.3390/plants11182427