
Metal Complexation of Bis-Chalcone Derivatives Enhances Their Efficacy against Fusarium Wilt Disease, Caused by Fusarium equiseti, via Induction of Antioxidant Defense Machinery
 ,
,  ,
, 
Abstract
:1. Introduction
2. Results
2.1. Metal Complexation of DMAPAPP Boosts Its Antioxidant Activity
2.2. Pathogenicity Test of Fusarium Isolates on Pepper Plants
2.3. Morphological Characterization
2.4. Molecular Identification of Fusarium Strain
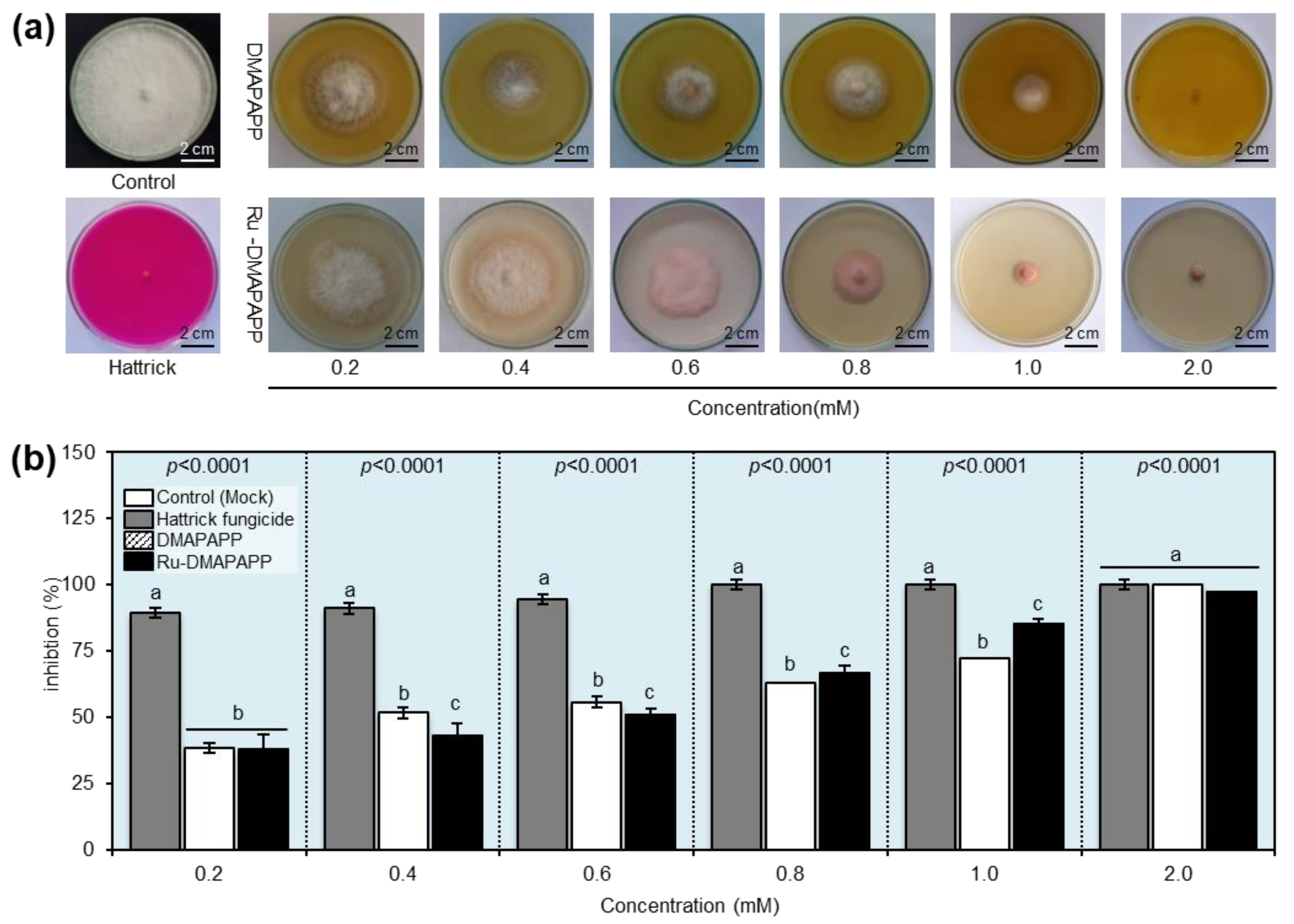
2.5. Antifungal Activity of DMAPAPP and Its Ru-DMAPAPP
2.6. Effects of DMAPAPP and Ru-DMAPAPP on the Development of Pepper Fusarium Wilt Disease under Greenhouse Conditions
2.7. DMAPAPP and Ru-DMAPAPP Complex Alleviate the Oxidative Stress of F. equiseti-Infected Leaves
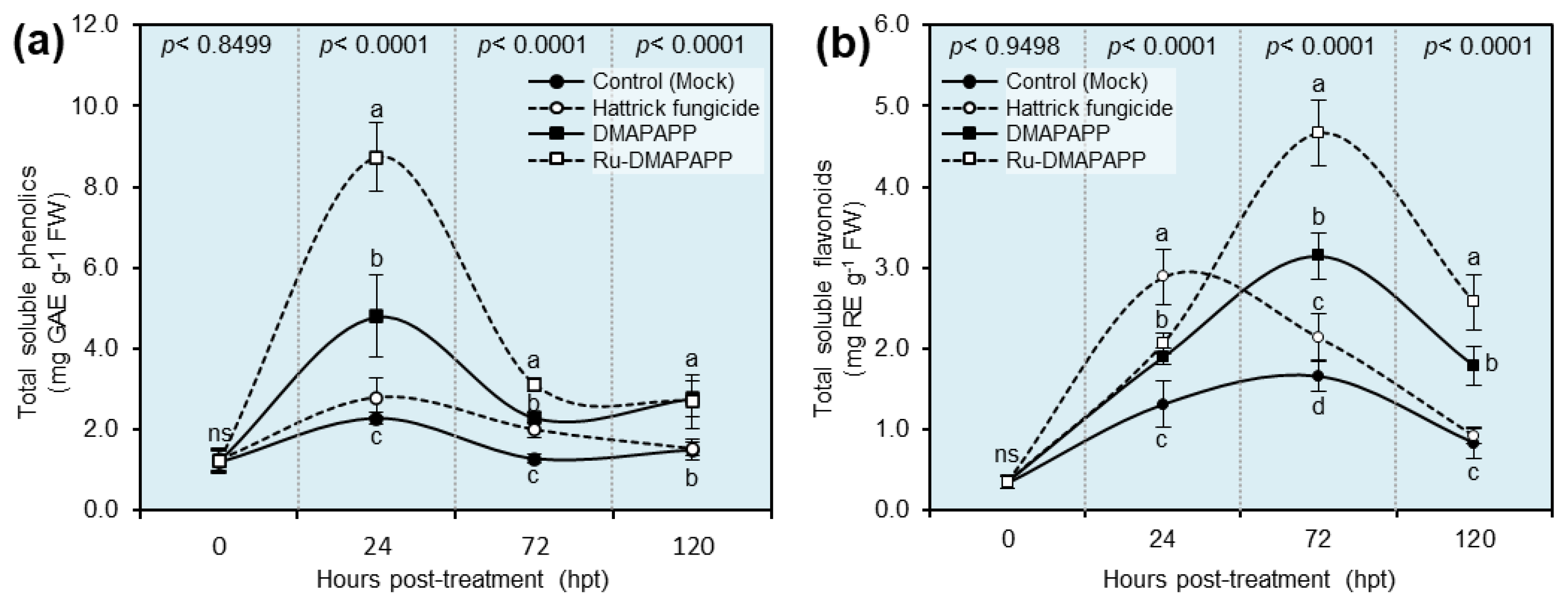
2.8. DMAPAPP and Ru-DMAPAPP Complex Enhanced the Profile of Total Soluble Phenolics and Flavonoids of F. equiseti-Infected Pepper Plants
2.9. Effects of DMAPAPP and Ru-DMAPAPP Complex on the Activity of Antioxidant Defense-Related Enzymes
2.10. DMAPAPP and Ru-DMAPAPP Stimulate the Expression of Antioxidant Enzymes
2.11. Effect of DMAPAPP and Ru-DMAPAPP on the Growth Parameters and Total Chlorophyll in F. equiseti-Infected Pepper Plants
3. Discussion
4. Materials and Methods
4.1. Tested Compounds
4.2. Antioxidant and Radical Scavenging Assay
4.3. Isolation, Morphological Characterization, Molecular Identification of Fusarium Isolates
4.3.1. Pathogen Isolation
4.3.2. Morphological and Cultural Characterization of Fusarium sp. Isolates
4.3.3. Molecular Identification of Fusarium sp. Isolates
4.3.4. Phylogenetic Analysis
4.4. Pathogenicity Test
4.5. Antifungal Activity
4.6. Greenhouse Experiment, Disease Assessment, and Growth Parameters
4.7. In Situ Histochemical Localization of Hydrogen Peroxide (H2O2) and Superoxide Anion (O2•−)
4.8. Total Soluble Phenolic and Total Flavonoid Compounds
4.9. Antioxidant Enzymatic Activity
4.10. Gene Expression Analysis
4.11. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Walsh, B.M.; Hoot, S.B. Phylogenetic Relationships of Capsicum (Solanaceae) Using DNA Sequences from Two Noncoding Regions: The Chloroplast atpB-rbcL Spacer Region and Nuclear waxy Introns. Int. J. Plant Sci. 2015, 162, 1409–1418. [Google Scholar] [CrossRef]
- Heiser, C.B.; Pickersgill, B. Names for the cultivated Capsicum species (solanaceae). Taxon 1969, 18, 277–283. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations (FAO). FAOSTAT. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 2 September 2022).
- Black, L.L.; Green, S.K.; Hartman, G.L.; Poulos, M. Pepper Diseases: A Field Guide; CTA: Wageningen, The Netherlands, 1993; ISBN 9290580. [Google Scholar]
- Roberts, P.D.; Adkins, S.; Pernezny, K.; Jones, J.B. Diseases of Pepper and Their Management. In Diseases of Fruits and Vegetables: Volume II; Springer: Dordrecht, The Netherlands, 2004; pp. 333–387. [Google Scholar] [CrossRef]
- Sanogo, S. Chile Pepper and The Threat of Wilt Diseases. Plant Health Prog. 2003, 4, 1–5. [Google Scholar] [CrossRef]
- Poddar, R.K.; Singh, D.V.; Dubey, S.C. Management of Chickpea Wilt through Combination of Fungicides and Bioagents. Indian Phytopathol. 2004, 57, 39–43. [Google Scholar]
- Klein, E.; Katan, J.; Gamliel, A. Soil Suppressiveness by Organic Amendment to Fusarium Disease in Cucumber: Effect on Pathogen and Host. Phytoparasitica 2016, 44, 239–249. [Google Scholar] [CrossRef]
- Wang, B.; Li, R.; Ruan, Y.; Ou, Y.; Zhao, Y.; Shen, Q. Soil Biology & Biochemistry Pineapple e Banana Rotation Reduced the Amount of Fusarium oxysporum More than Maize e Banana Rotation Mainly through Modulating Fungal Communities. Soil Biol. Biochem. 2015, 86, 77–86. [Google Scholar] [CrossRef]
- Gabrekiristos, E.; Demiyo, T. Hot Pepper Fusarium Wilt (Fusarium oxysporum f. sp. capsici): Epidemics, Characteristic Features and Management Options. J. Agric. Sci. 2020, 12, 347–360. [Google Scholar] [CrossRef]
- Hewedy, O.A. Genetic Diversity and Biocontrol Efficacy of Indigenous Trichoderma Isolates against Fusarium Wilt of Pepper. J. Basic Microbiol. 2020, 60, 126–135. [Google Scholar] [CrossRef]
- Lomas-Cano, T.; Palmero-Llamas, D.; de Cara, M.; García-Rodríguez, C.; Boix-Ruiz, A.; Camacho-Ferre, F.; Tello-Marquina, J.C. First Report of Fusarium oxysporum on Sweet Pepper Seedlings in Almería, Spain. Plant Dis. 2014, 98, 1435. [Google Scholar] [CrossRef]
- Naik, M. Wilt of Chilli with Special Reference to Cultural, Morphological, Molecular Characterization and Pathogenic Variability of Fusarium Isolates of India. In Proceedings of the Midterm Review Meeting of the Project, Held at Indian Institute of Vegetable Research, Varanasi, India, 23 July 2006. [Google Scholar]
- Tembhurne, B.; Belabadevi, B.; Kisan, B.; Tilak, I.; Ashwathanarayana, D.; Naik, M. Molecular Characterization and Screening for Fusarium (Fusarium solani) Resistance in Chilli (Capsicum annuum L.) Genotypes. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 1585–1597. [Google Scholar] [CrossRef] [Green Version]
- Hami, A.; Rasool, R.S.; Khan, N.A.; Mir, M.A.; Ahmed, N.; Masoodi, K.Z. Morpho-Molecular Identification and First Report of Fusarium Equiseti in Causing Chilli Wilt from Kashmir (Northern Himalayas). Sci. Rep. 2021, 11, 3610. [Google Scholar] [CrossRef] [PubMed]
- López-Seijas, J.; García-Fraga, B.; da Silva, A.F.; Sieiro, C. Wine Lactic Acid Bacteria with Antimicrobial Activity as Potential Biocontrol Agents against Fusarium oxysporum f. sp. lycopersici. Agronomy 2020, 10, 31. [Google Scholar] [CrossRef]
- Lamichhane, J.R.; Dürr, C.; Schwanck, A.A.; Robin, M.H.; Sarthou, J.P.; Cellier, V.; Messéan, A.; Aubertot, J.N. Integrated Management of Damping-off Diseases. A Review. Agron. Sustain. Dev. 2017, 37, 10–35. [Google Scholar] [CrossRef]
- Dhar, D.N. The Chemistry of Chalcones and Related Compounds; John Wiley & Sons: Hoboken, NJ, USA, 1981. [Google Scholar]
- López, S.N.; Castelli, M.V.; Zacchino, S.A.; Domínguez, J.N.; Lobo, G.; Charris-Charris, J.; Cortés, J.C.G.; Ribas, J.C.; Devia, C.; Rodríguez, A.M.B.; et al. In Vitro Antifungal Evaluation and Structure Activity Relationships of a New Series of Chalcone Derivatives and Synthetic Analogues, with Inhibitory Properties against Polymers of the Fungal Cell Wall. Bioorg. Med. Chem. 2001, 9, 1999–2013. [Google Scholar] [CrossRef]
- Elias, D.W.; Beazely, M.A.; Kandepu, N.M. Bioactivities of Chalcones. Curr. Med. Chem. 1999, 6, 1125–1149. [Google Scholar]
- Go, M.L.; Wu, X.; Liu, X.L. Chalcones: An Update on Cytotoxic and Chemoprotective Properties. Curr. Med. Chem. 2005, 12, 483–499. [Google Scholar] [CrossRef]
- Singh, P.; Anand, A.; Kumar, V. Recent Developments in Biological Activities of Chalcones: A Mini Review. Eur. J. Med. Chem. 2014, 85, 758–777. [Google Scholar] [CrossRef]
- Sökmen, M.; Akram Khan, M. The Antioxidant Activity of Some Curcuminoids and Chalcones. Inflammopharmacology 2016, 24, 81–86. [Google Scholar] [CrossRef]
- Zheng, Y.; Wang, X.; Gao, S.; Ma, M.; Ren, G.; Liu, H.; Chen, X. Synthesis and Antifungal Activity of Chalcone Derivatives. Nat. Prod. Res. 2015, 29, 1804–1810. [Google Scholar] [CrossRef]
- López, G.; Mellado, M.; Werner, E.; Said, B.; Godoy, P.; Caro, N.; Besoain, X.; Montenegro, I.; Madrid, A. Sonochemical Synthesis of 2′-Hydroxy-Chalcone Derivatives with Potential Anti-Oomycete Activity. Antibiotics 2020, 9, 576. [Google Scholar] [CrossRef]
- Zhou, Q.; Tang, X.; Chen, S.; Zhan, W.; Hu, D.; Zhou, R.; Sun, N.; Wu, Y.; Xue, W. Design, Synthesis, and Antifungal Activity of Novel Chalcone Derivatives Containing a Piperazine Fragment. J. Agric. Food Chem. 2022, 70, 1029–1036. [Google Scholar] [CrossRef] [PubMed]
- Gaber, M.; El-Sayed, Y.S.; Diab, H. Spectral Behavior Study of 3-(4-Dimethylamino-Phenyl)-1-{6-[3-(4-Dimethylamino-Phenyl)-Acryloyl]-Pyridin-2-Yl}-Propanone. Opt. Laser Technol. 2011, 43, 592–598. [Google Scholar] [CrossRef]
- El-Sayed, Y.S.; Gaber, M.; El-Wakiel, N. Design of Mn(II), Fe(III) and Ru(III) Chalcone Complexes: Structural Elucidation, Spectral, Thermal and Catalytic Activity Studies. J. Mol. Struct. 2021, 1224, 129283. [Google Scholar] [CrossRef]
- Abada, K.A.; Ahmed, M.A. Management Fusarium Wilt of Sweet Pepper by Bacillus Strains. Am. J. Life Sci. 2014, 2, 19–25. [Google Scholar] [CrossRef]
- Ammar, M.M.; Khalifa, E.Z.; Kamel, S.M.H. Integrated Control of Root Rot and Wilt Diseases of Pepper. Menoufia J. Plant Prot. 2020, 5, 185–186. [Google Scholar] [CrossRef]
- Gupta, D.; Jain, D.K. Chalcone Derivatives as Potential Antifungal Agents: Synthesis, and Antifungal Activity. J. Adv. Pharm. Technol. Res. 2015, 6, 114–117. [Google Scholar] [CrossRef]
- Boeck, P.; Leal, P.C.; Yunes, R.A.; Filho, V.C.; López, S.; Sortino, M.; Escalante, A.; Furlán, R.L.E.; Zacchino, S. Antifungal Activity and Studies on Mode of Action of Novel Xanthoxyline-Derived Chalcones. Arch. Pharm. Int. J. Pharm. Med. Chem. 2005, 338, 87–95. [Google Scholar] [CrossRef]
- Reddy, N.P.; Aparoy, P.; Reddy, T.C.M.; Achari, C.; Sridhar, P.R.; Reddanna, P. Design, Synthesis, and Biological Evaluation of Prenylated Chalcones as 5-LOX Inhibitors. Bioorg. Med. Chem. 2010, 18, 5807–5815. [Google Scholar] [CrossRef]
- Lahtchev, K.L.; Batovska, D.I.; Parushev, S.P.; Ubiyvovk, V.M.; Sibirny, A.A. Antifungal Activity of Chalcones: A Mechanistic Study Using Various Yeast Strains. Eur. J. Med. Chem. 2008, 43, 2220–2228. [Google Scholar] [CrossRef]
- Tweedy, B.G. Plant Extracts with Metal Ions as Potential Antimicrobial Agents. Phytopathology 1964, 55, 910–914. [Google Scholar]
- Gacche, R.N.; Dhole, N.A.; Kamble, S.G.; Bandgar, B.P. In-Vitro Evaluation of Selected Chalcones for Antioxidant Activity. J. Enzym. Inhib. Med. Chem. 2008, 23, 28–31. [Google Scholar] [CrossRef] [PubMed]
- Vásquez-Martínez, Y.A.; Osorio, M.E.; San Martín, D.A.; Carvajal, M.A.; Vergara, A.P.; Sanchez, E.; Raimondi, M.; Zacchino, S.A.; Mascayano, C.; Torrent, C.; et al. Antimicrobial, Anti-Inflammatory and Antioxidant Activities of Polyoxygenated Chalcones. J. Braz. Chem. Soc. 2019, 30, 286–304. [Google Scholar] [CrossRef]
- Mittal, A.; Vashistha, V.K.; Das, D.K. Recent advances in the antioxidant activity and mechanisms of chalcone derivatives: A computational review. Free Radic. Res. 2022, 1–39. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Tuteja, N. Reactive Oxygen Species and Antioxidant Machinery in Abiotic Stress Tolerance in Crop Plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Racchi, M.L. Antioxidant Defenses in Plants with Attention to Prunus and Citrus spp. Antioxidants 2013, 2, 340–369. [Google Scholar] [CrossRef]
- Dumanović, J.; Nepovimova, E.; Natić, M.; Kuča, K.; Jaćević, V. The Significance of Reactive Oxygen Species and Antioxidant Defense System in Plants: A Concise Overview. Front. Plant Sci. 2021, 11, 2106. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive Oxygen Species: Metabolism, Oxidative Stress, and Signal Transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef]
- Dvořák, P.; Krasylenko, Y.; Zeiner, A.; Šamaj, J.; Takáč, T. Signaling toward reactive oxygen species-scavenging enzymes in plants. Front. Plant Sci. 2021, 11, 2178. [Google Scholar] [CrossRef]
- Bestwick, C.S.; Brown, I.R.; Mansfield, J.W. Localized Changes in Peroxidase Activity Accompany Hydrogen Peroxide Generation during the Development of a Nonhost Hypersensitive Reaction in Lettuce. Plant Physiol. 1998, 118, 1067–1078. [Google Scholar] [CrossRef] [PubMed]
- Noctor, G.; Foyer, C.H. Ascorbate and Glutathione: Keeping Active Oxygen under Control. Annu. Rev. Plant Biol. 1998, 49, 249–279. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Parvin, K.; Bardhan, K.; Nahar, K.; Anee, T.I.; Masud, A.A.C.; Fotopoulos, V. Biostimulants for the regulation of reactive oxygen species metabolism in plants under abiotic stress. Cells 2021, 10, 2537. [Google Scholar] [CrossRef] [PubMed]
- Obianom, C.; Romanazzi, G.; Sivakumar, D. Effects of Chitosan Treatment on Avocado Postharvest Diseases and Expression of Phenylalanine Ammonia-Lyase, Chitinase and Lipoxygenase Genes. Postharvest Biol. Technol. 2019, 147, 214–221. [Google Scholar] [CrossRef]
- Fu, L.; Xu, B.T.; Xu, X.R.; Gan, R.Y.; Zhang, Y.; Xia, E.Q.; Li, H. Bin Antioxidant Capacities and Total Phenolic Contents of 62 Fruits. Food Chem. 2011, 129, 345–350. [Google Scholar] [CrossRef]
- Reddy, A.M.; Reddy, V.S.; Scheffler, B.E.; Wienand, U.; Reddy, A.R. Novel Transgenic Rice Overexpressing Anthocyanidin Synthase Accumulates a Mixture of Flavonoids Leading to an Increased Antioxidant Potential. Metab. Eng. 2007, 9, 95–111. [Google Scholar] [CrossRef]
- Hijaz, F.; Nehela, Y.; Jones, S.E.; Dutt, M.; Grosser, J.W.; Manthey, J.A.; Killiny, N. Metabolically Engineered Anthocyanin-Producing Lime Provides Additional Nutritional Value and Antioxidant Potential to Juice. Plant Biotechnol. Rep. 2018, 12, 329–346. [Google Scholar] [CrossRef]
- Atallah, O.; Yassin, S. Aspergillus spp. Eliminate Sclerotinia Sclerotiorum by Imbalancing the Ambient Oxalic Acid Concentration and Parasitizing Its Sclerotia. Environ. Microbiol. 2020, 22, 5265–5279. [Google Scholar] [CrossRef] [PubMed]
- Atallah, O.O.; Mazrou, Y.S.M.; Atia, M.M.; Nehela, Y.; Abdelrhim, A.S.; Nader, M.M. Polyphasic Characterization of Four Aspergillus Species as Potential Biocontrol Agents for White Mold Disease of Bean. J. Fungi 2022, 8, 626. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Zhou, L.; Yang, C.; Cao, X.; Zhang, L.; Liu, X. Tomato Fusarium Wilt and Its Chemical Control Strategies in a Hydroponic System. Crop Prot. 2004, 23, 243–247. [Google Scholar] [CrossRef]
- Grover, R.; Moore, J.D. Toximetric Studies of Fungicides against the Brown Root Organisms, Sclerotinia Fructicola and S. Laxa. Phytopathology 1962, 52, 876–880. [Google Scholar]
- Romero-Puertas, M.C.; Rodríguez-Serrano, M.; Corpas, F.J.; Gómez, M.; Del Río, L.A.; Sandalio, L.M. Cadmium-Induced Subcellular Accumulation of O2− and H2O2 in Pea Leaves. Plant Cell Environ. 2004, 27, 1122–1134. [Google Scholar] [CrossRef]
- Shi, J.; Fu, X.-Z.; Peng, T.; Huang, X.-S.; Fan, Q.-J.; Liu, J.-H. Spermine Pretreatment Confers Dehydration Tolerance of Citrus In Vitro Plants via Modulation of Antioxidative Capacity and Stomatal Response. Tree Physiol. 2010, 30, 914–922. [Google Scholar] [CrossRef] [PubMed]
- Killiny, N.; Nehela, Y.; Hijaz, F.; Gonzalez-Blanco, P.; Hajeri, S.; Gowda, S. Knock-down of δ-Aminolevulinic Acid Dehydratase via Virus-Induced Gene Silencing Alters the MicroRNA Biogenesis and Causes Stress-Related Reactions in Citrus Plants. Plant Sci. 2020, 299, 110622. [Google Scholar] [CrossRef]
- Nehela, Y.; Taha, N.A.; Elzaawely, A.A.; Xuan, T.D.; Amin, M.A.; Ahmed, M.E.; El-Nagar, A. Benzoic Acid and Its Hydroxylated Derivatives Suppress Early Blight of Tomato (Alternaria solani) via the Induction of Salicylic Acid Biosynthesis and Enzymatic and Nonenzymatic Antioxidant Defense Machinery. J. Fungi 2021, 7, 663. [Google Scholar] [CrossRef] [PubMed]
- Hückelhoven, R.; Fodor, J.; Preis, C.; Kogel, K.H. Hypersensitive Cell Death and Papilla Formation in Barley Attacked by the Powdery Mildew Fungus Are Associated with Hydrogen Peroxide but Not with Salicylic Acid Accumulation. Plant Physiol. 1999, 119, 1251–1260. [Google Scholar] [CrossRef]
- Kähkönen, M.P.; Hopia, A.I.; Vuorela, H.J.; Rauha, J.P.; Pihlaja, K.; Kujala, T.S.; Heinonen, M. Antioxidant Activity of Plant Extracts Containing Phenolic Compounds. J. Agric. Food Chem. 1999, 47, 3954–3962. [Google Scholar] [CrossRef]
- Djeridane, A.; Yousfi, M.; Nadjemi, B.; Boutassouna, D.; Stocker, P.; Vidal, N. Antioxidant Activity of Some Algerian Medicinal Plants Extracts Containing Phenolic Compounds. Food Chem. 2006, 97, 654–660. [Google Scholar] [CrossRef]
- Harrach, B.D.; Fodor, J. Antioxidant, Ethylene and Membrane Leakage Responses to Powdery Mildew Infection of near-Isogenic Barley Lines with Various Types of Resistance. Eur. J. Plant Pathol. 2008, 121, 21–33. [Google Scholar] [CrossRef]
- Malik, C.P.; Singh, M.B. Plant Enzymology and Histo-Enzymology; Kalyani Publishers: New Delhi, India, 1980. [Google Scholar]
- Aebi, H. Catalase in Vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Plant Height (cm plant−1) | Number of Leaves | Shoot Fresh Weight (g plant−1) | Total Leaf Area (cm2) | T. Chlorophyll (Spad) |
|---|---|---|---|---|---|
| Mock control | 26.91 ± 4.77 a | 21.50 ± 4.25 b | 6.207 ± 1.14 c | 07.03 ± 1.20 b | 38.28 ± 13.84 c |
| Hattrick fungicide | 21.75 ± 2.83 b | 20.25 ± 2.22 b | 6.238 ± 1.02 c | 05.38 ± 0.76 b | 65.35 ± 5.06 b |
| DMAPAPP | 27.41 ± 2.02 a | 18.50 ± 4.23 b | 11.57 ± 1.09 b | 05.17 ± 0.62 b | 72.24 ± 7.27 ab |
| Ru-DMAPAPP | 30.58 ± 3.55 a | 29.00 ± 2.21 a | 14.39 ± 3.57 a | 12.38 ± 3.72 a | 81.09 ± 2.67 a |
| Description | Gene | NCBI Accession Number | Primer (Forward and Reverse) | |
| Superoxide dismutase [Cu-Zn] | CaSOD-Cu | NM_001398340.1 | F: R: | TACCACAAATGGCTGCATGT TTTGCTGAGCTCATGTCCAC |
| L-ascorbate peroxidase 1, cytosolic | CaAPX1 | NM_001325037.1 | F: R: | TAGGGAGCAGTTTCCCACAC AACACGTCCCTCAAGTGGTC |
| Glutathione reductase, chloroplastic | CaGR1 | XM_016710630.2 | F: R: | GGTGGAGGGTACATTGCTGT TGCCACCTTCTTCTGCTTTT |
| monodehydroascorbate reductase | CaMDHAR | XM_016687442.2 | F: R: | AGATCGTTGGTGCATTCCTC ATCAACCAGGCACGAAAAAC |
| Actin (Housekeeping gene) | CaACTIN | XM_016722297.2 | F: R: | CCTCGTCACACGGGAGTAAT CACGATTAGCCTTGGGGTTA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Nagar, A.; Elzaawely, A.A.; Xuan, T.D.; Gaber, M.; El-Wakeil, N.; El-Sayed, Y.; Nehela, Y. Metal Complexation of Bis-Chalcone Derivatives Enhances Their Efficacy against Fusarium Wilt Disease, Caused by Fusarium equiseti, via Induction of Antioxidant Defense Machinery. Plants 2022, 11, 2418. https://doi.org/10.3390/plants11182418
El-Nagar A, Elzaawely AA, Xuan TD, Gaber M, El-Wakeil N, El-Sayed Y, Nehela Y. Metal Complexation of Bis-Chalcone Derivatives Enhances Their Efficacy against Fusarium Wilt Disease, Caused by Fusarium equiseti, via Induction of Antioxidant Defense Machinery. Plants. 2022; 11(18):2418. https://doi.org/10.3390/plants11182418
Chicago/Turabian StyleEl-Nagar, Asmaa, Abdelnaser A. Elzaawely, Tran Dang Xuan, Mohamed Gaber, Nadia El-Wakeil, Yusif El-Sayed, and Yasser Nehela. 2022. "Metal Complexation of Bis-Chalcone Derivatives Enhances Their Efficacy against Fusarium Wilt Disease, Caused by Fusarium equiseti, via Induction of Antioxidant Defense Machinery" Plants 11, no. 18: 2418. https://doi.org/10.3390/plants11182418