
Response of Bletilla striata to Drought: Effects on Biochemical and Physiological Parameter Also with Electric Measurements

Abstract
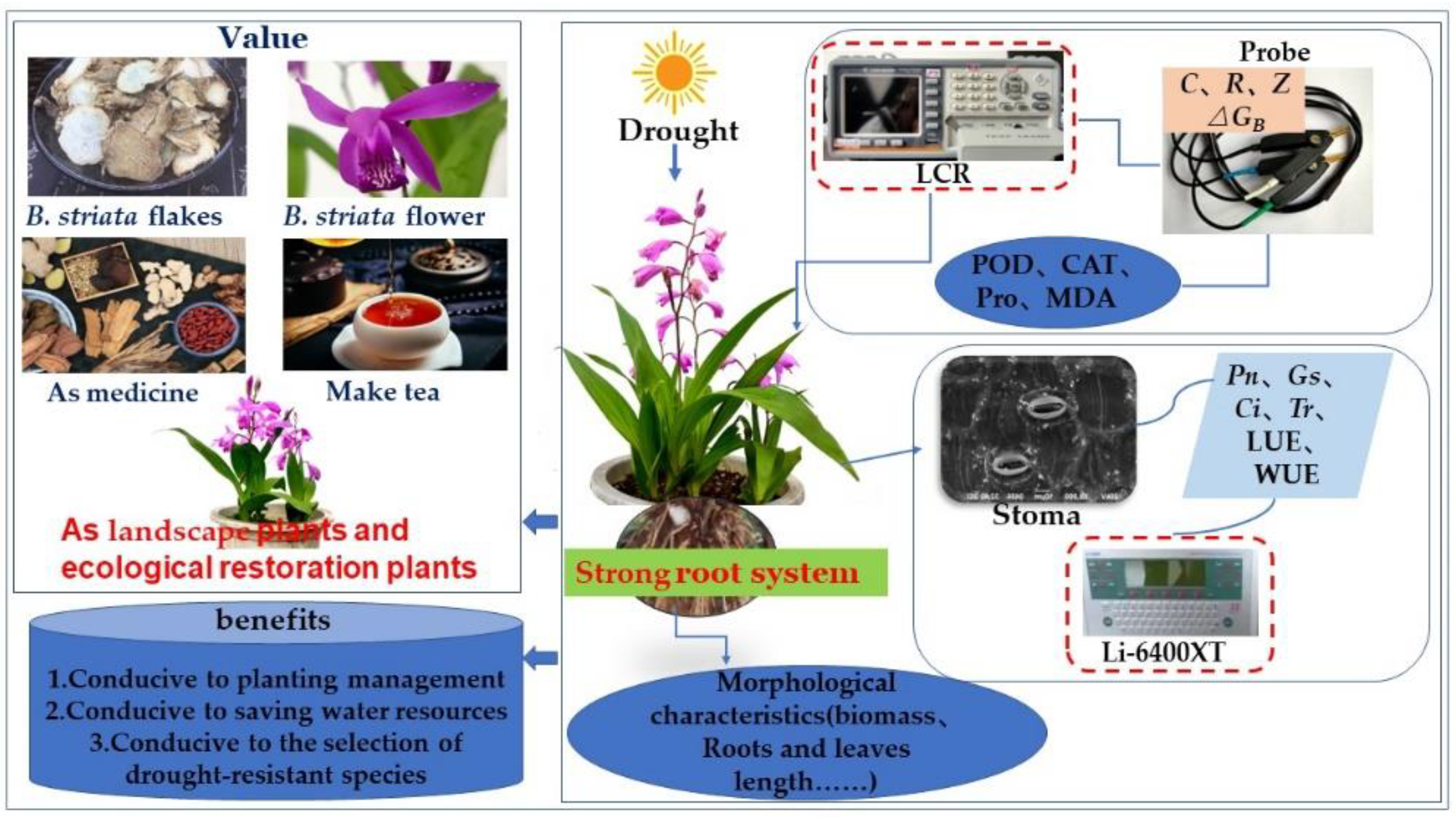
:1. Introduction
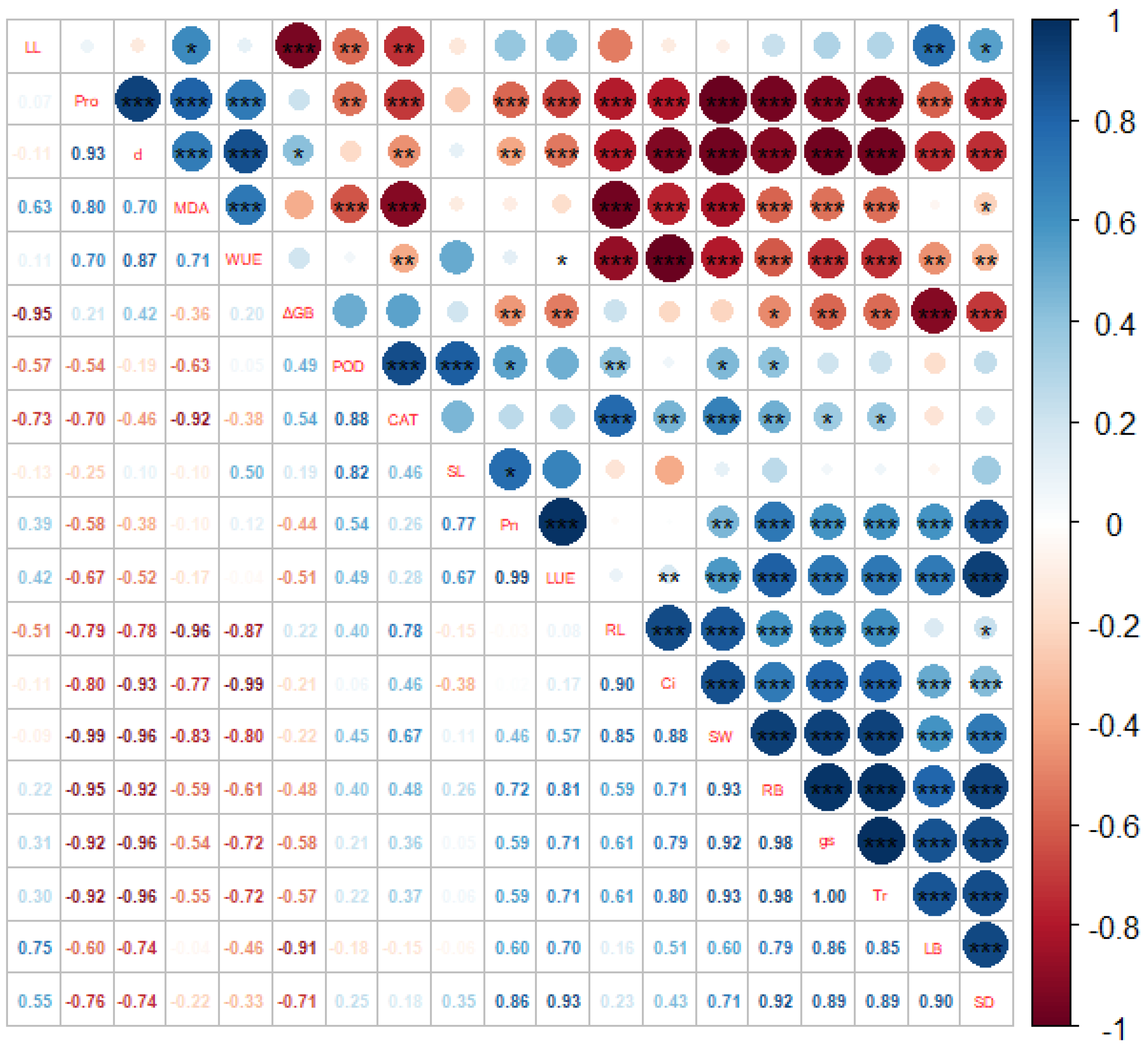
2. Results
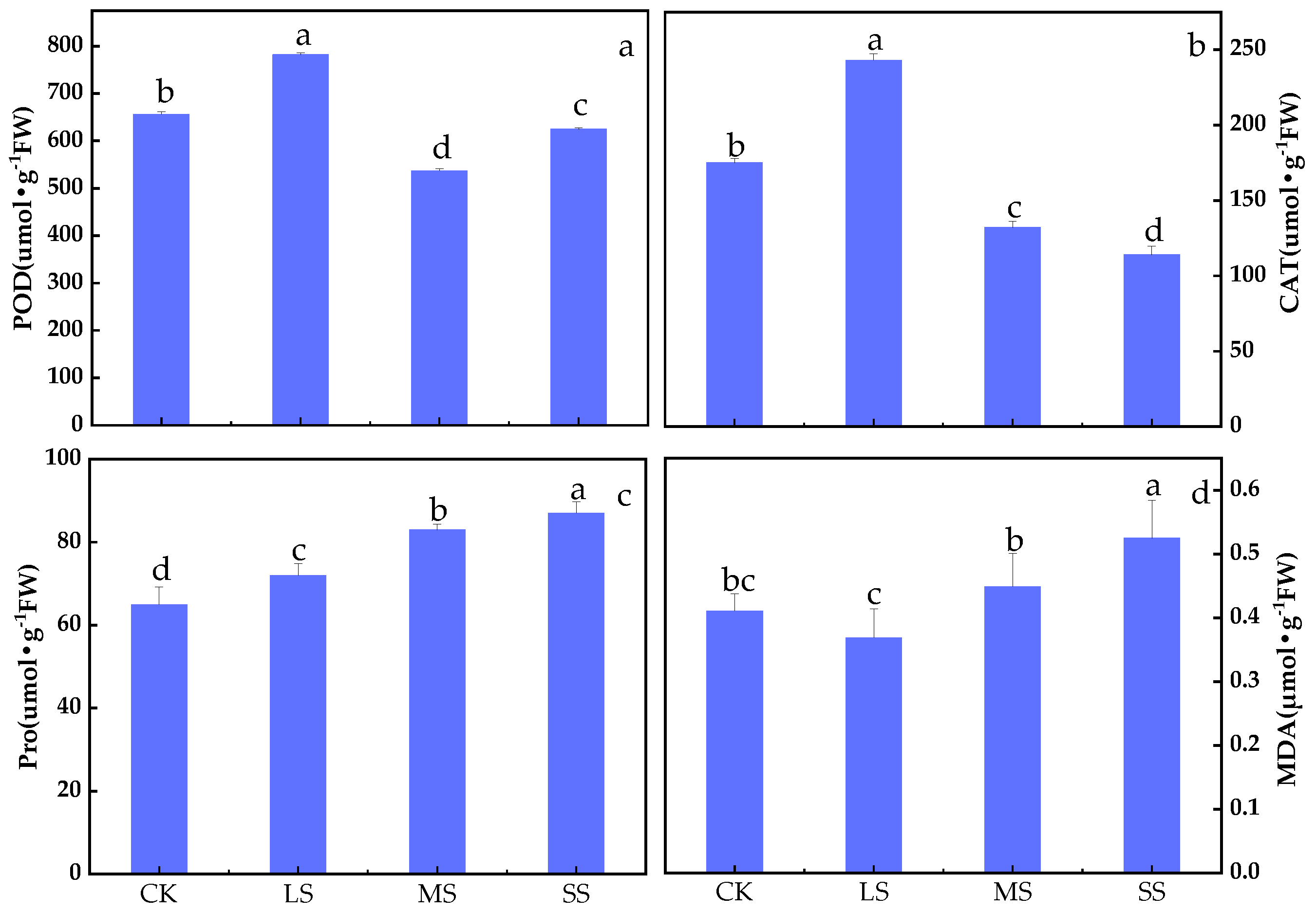
2.1. B. striata Enzyme Activity Analysis
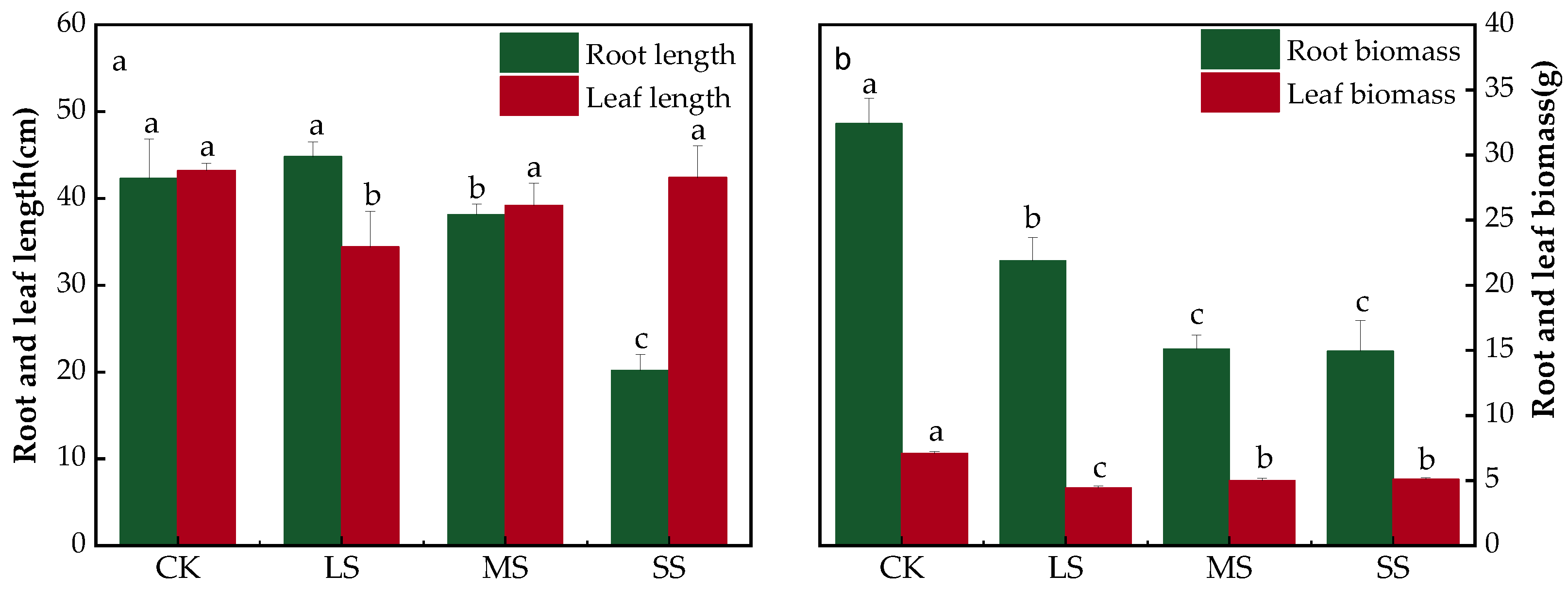
2.2. B. striata Root Lengths and Biomass Analysis
2.3. Analysis of B. striata Photosynthetic Characteristics
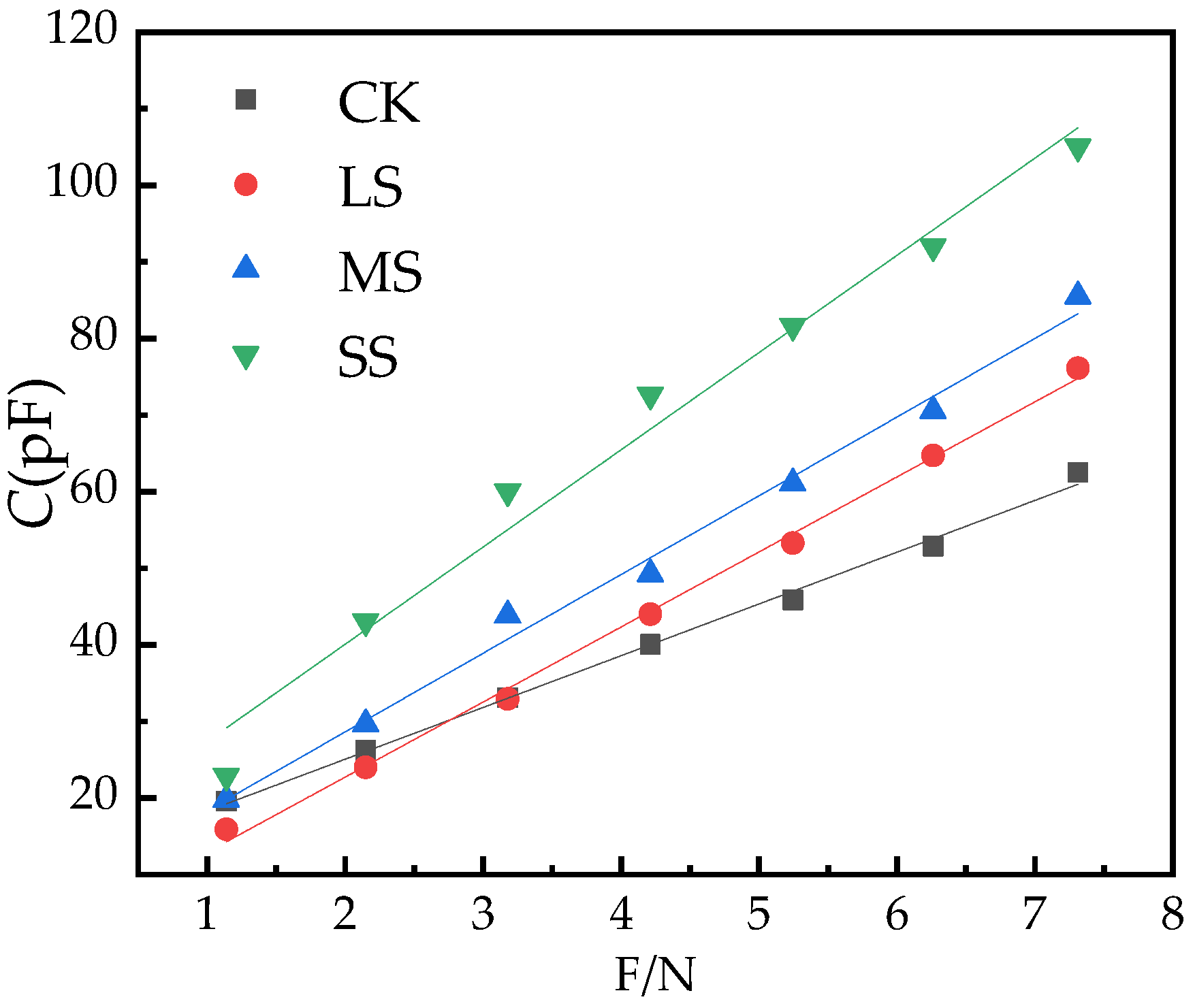
2.4. B. striata Physiological Capacitance Model
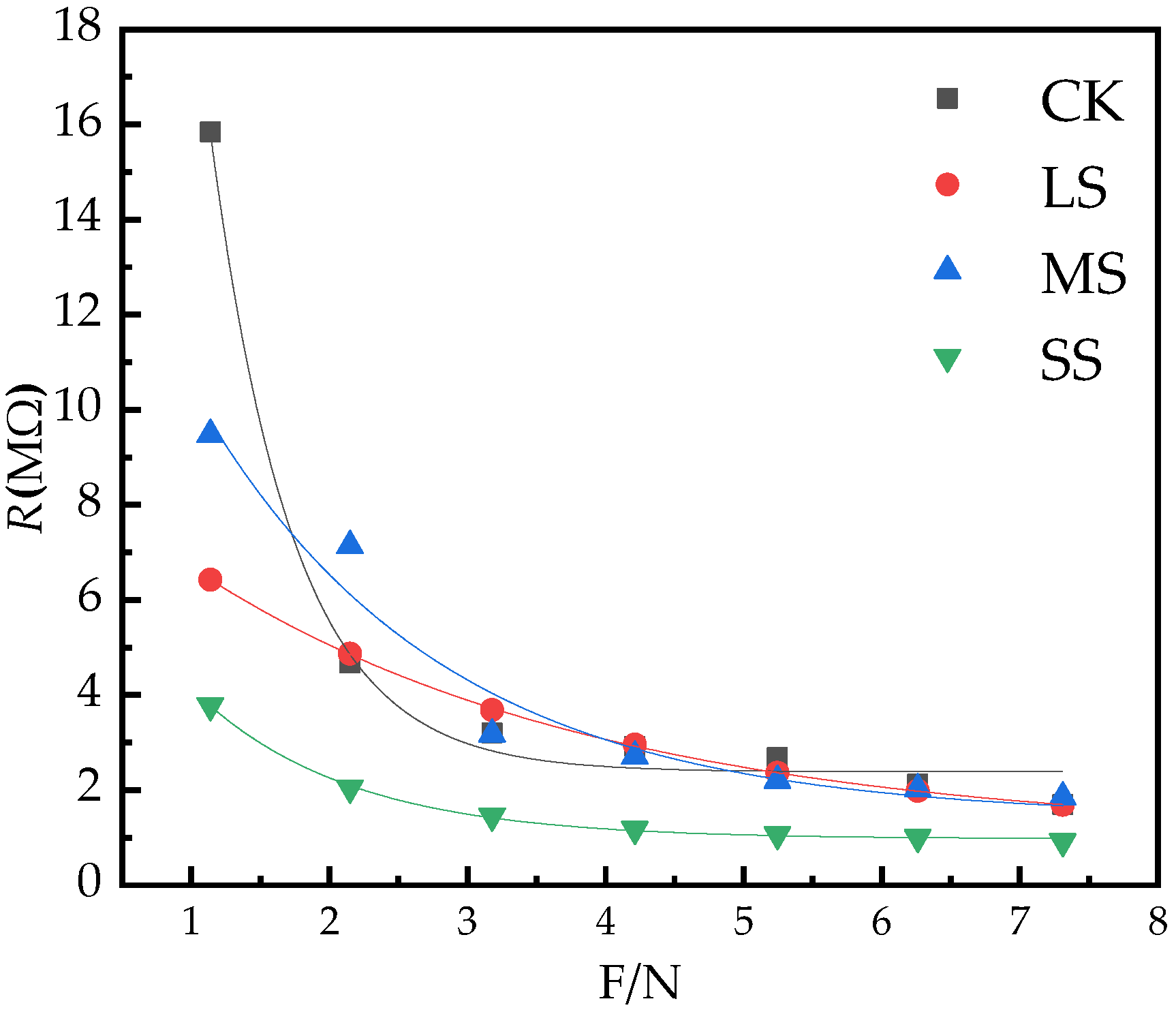
2.5. B. striata Physiological Resistance Model
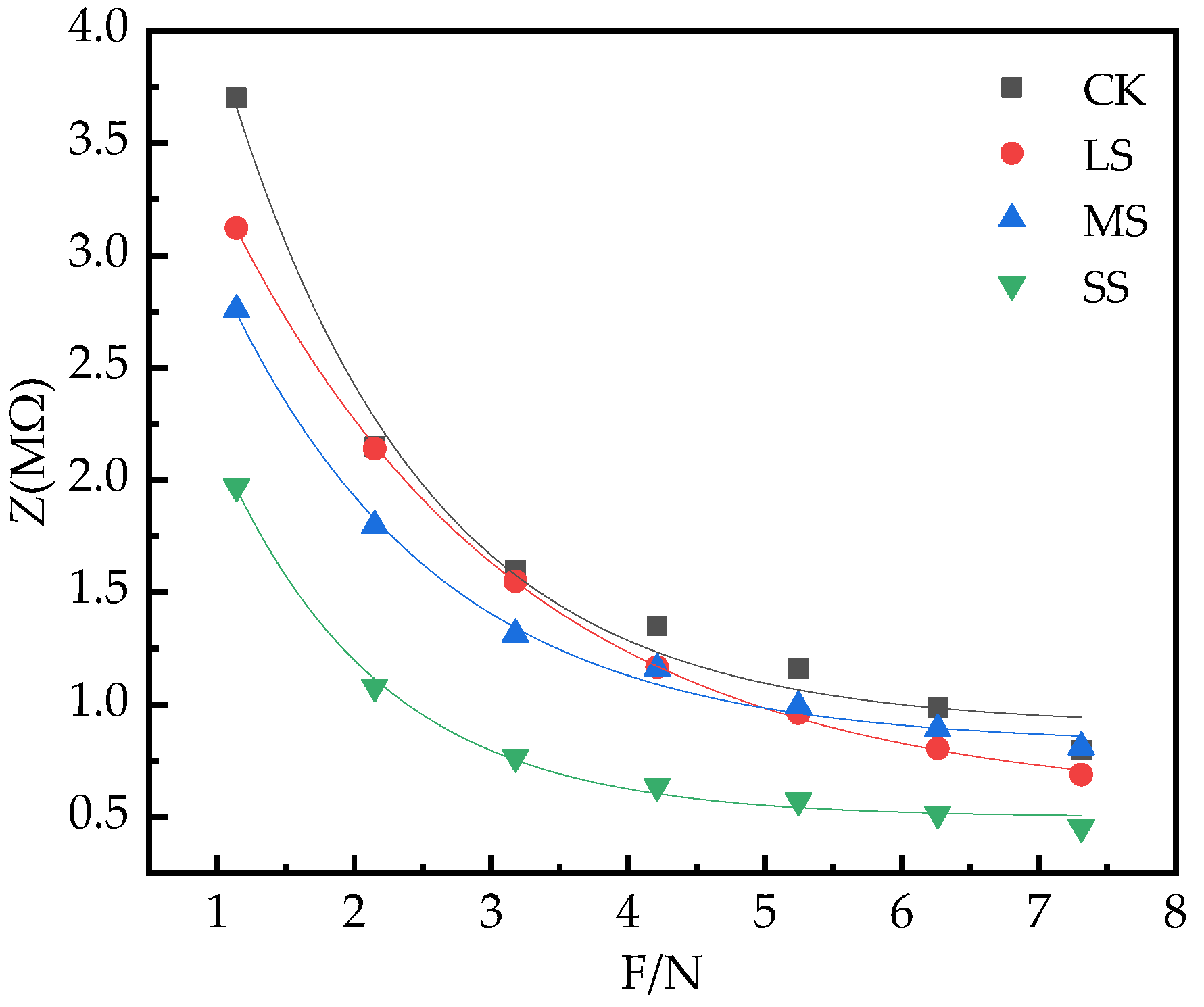
2.6. B. striata Physiological Impedance (Z) Model
2.7. Cell Metabolic Energy Analysis
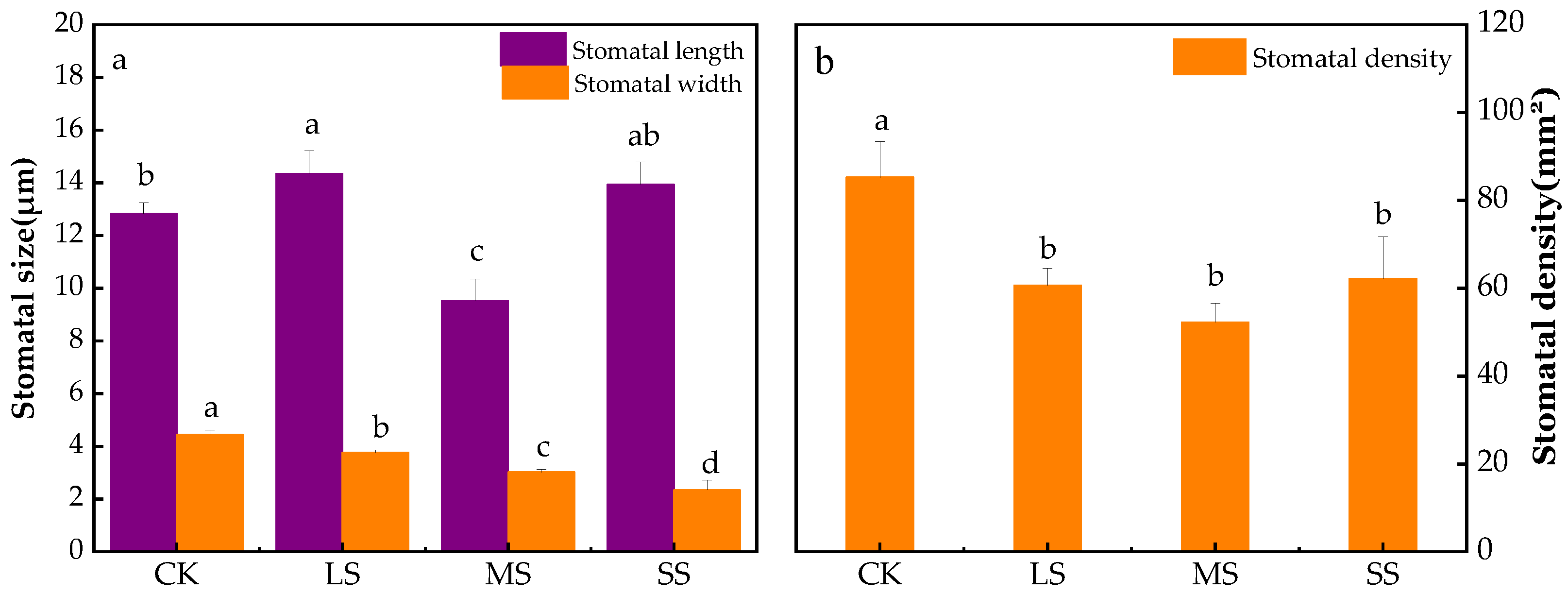
2.8. B. striata Stomatal Feature Analysis
3. Discussion
3.1. Response of B. striata Enzyme Activities to Drought Stress
3.2. Morphological Characteristics of B. striata under Drought Environments
3.3. Response of B. striata Photosynthetic Characteristics to Drought Stress
3.4. Response of B. striata Electrophysiology to Drought Stress
4. Materials and Methods
4.1. Experimental Materials and Experimental Design
4.2. POD Activity Assay
4.3. CAT Activity Assay
4.4. Pro Content Assay
4.5. MDA Content Assay
4.6. B. striata Biomass and Growth Parameters Assay
4.7. B. striata Photosynthetic Parameters Assay
4.8. Establishment of Electrophysiological Model and Acquisition of Cell Metabolic Energy
4.9. Physiological Capacitance
4.10. Physiological Resistance
4.11. Physiological Impedance
4.12. B. striata Stomatal Scanning
4.13. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gorai, M.; Aloui, E.W.; Yang, X.; Neffati, M. Toward understanding the ecological role of mucilage in seed germination of a desert shrub Henophyton deserti: Interactive effects of temperature, salinity and osmotic stress. Plant Soil 2014, 374, 727–738. [Google Scholar] [CrossRef]
- Ha, C.V.; Leyva-González, M.A.; Osakabe, Y.; Tran, U.T.; Nishiyama, R.; Watanabe, Y.; Tanaka, M.; Seki, M.; Yamaguchi, S.; Dong, N.V. Positive regulatory role of strigolactone in plant responses to drought and salt stress. Proc. Natl. Acad. Sci. USA. 2014, 111, 851–856. [Google Scholar] [CrossRef]
- Omprakash; Gobu, R.; Bisen, P.; Baghel, M.; Chourasia, K.N. Resistance/Tolerance Mechanism under Water Deficit (Drought) Condition in Plants. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 66–78. [Google Scholar]
- Ashraf, M. Inducing drought tolerance in plants: Recent advances. Biotechnol. Adv. 2010, 28, 169–183. [Google Scholar] [CrossRef] [PubMed]
- Xing, D.K.; Wu, Y.Y.; Wang, R.; Fu, W.G.; Zhou, Y.C.; Javed, Q. Effects of drought stress on photosynthesis and glucose-6-phosphate dehydrogenase activity of two biomass energy plants (Jatrophacurcas L. and Verniciafordii H.). J. Anim. Plant Sci. 2015, 25, 172–179. [Google Scholar]
- Fang, Y.; Xiong, L. General mechanisms of drought response and their application in drought resistance improvement in plants. Cell Mol. Life Sci. 2015, 72, 673–689. [Google Scholar] [CrossRef] [PubMed]
- Bhusal, N.; Lee, M.; Han, A.R.; Han, A.; Kim, H.S. Responses to drought stress in Prunus sargentii and Larix kaempferi seedlings using morphological and physiological parameters. Forest Ecol. Manag. 2020, 465, 118099. [Google Scholar] [CrossRef]
- Yang, X.L.; Li, Y.Y.; Chen, H.B.; Huang, J.; Zhang, Y.M.; Qi, M.F.; Li, Y.F.; Li, T.L. Photosynthetic response mechanism of soil salinity-induced cross-tolerance to subsequent drought stress in tomato plants. Plants 2020, 9, 363. [Google Scholar] [CrossRef]
- Gao, S.S.; Wang, Y.L.; Yu, S.; Huang, Y.Q.; Liu, H.C.; Chen, W.; He, X.Y. Effects of drought stress on growth, physiology and secondary metabolites of two Adonis species in northeast China. Sci. Hortic. 2020, 259, 108795. [Google Scholar] [CrossRef]
- Mcdowell, N.G. Mechanisms Linking Drought, Hydraulics, Carbon Metabolism, and Vegetation Mortality. Plant Physiol. 2011, 155, 1051–1059. [Google Scholar] [CrossRef]
- Chaves, M.M.; Oliveira, M.M. Mechanisms underlying plant re-silience to water deficits: Prospects for water-saving agriculture. J. Exp. Bot. 2004, 55, 2365–2384. [Google Scholar] [CrossRef]
- Bhusal, N.; Lee, M.; Lee, H.; Adhikari, A.; Han, A.R.; Han, A.; Kim, H.S. Evaluation of morphological, physiological, and biochemical traits for assessing drought resistance in eleven tree species. Sci. Total Environ. 2021, 779, 146466. [Google Scholar] [CrossRef] [PubMed]
- Schuppler, U.; He, P.H.; John, P.C.L.; Munns, R. Effects of water stress on cell division and cell-division-cycle-2-like cell-cycle ki-nase activity in wheat leaves. Plant Physiol. 1998, 117, 667–678. [Google Scholar] [CrossRef] [PubMed]
- Lipiec, J.; Doussan, C.; Nosalewicz, A.; Kondracka, K. Effect of drought and heat stresses on plant growth and yield: A review. Int Agrophys. 2013, 27, 463–477. [Google Scholar] [CrossRef]
- Kavar, T.; Maras, M.; Kidric, M.; Šuštar-Vozlič, J.; Meglič, V. Identification of genes involved in the response of leaves of Phaseolus vulgaris to drought stress. Mol. Breed. 2007, 21, 159–172. [Google Scholar] [CrossRef]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress: Effects, mechanisms and management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar] [CrossRef]
- Lundholm, J.; Tran, S.; Gebert, L. Plant functional traits predict green roof ecosystem services. Environ. Sci. Technol. 2015, 49, 2366–2374. [Google Scholar] [CrossRef]
- Yin, C.; Peng, Y.; Zang, R.; Zhu, Y.; Li, C. Adaptive responses of Populus kangdingensis to drought stress. Physiol. Plantarum. 2005, 123, 445–451. [Google Scholar] [CrossRef]
- Arndt, S.K.; Clifford, S.C.; Wanek, W.; Jones, H.G.; Popp, M. Physiological and morphological adaptations of the fruit tree Ziziphus rotundifolia in response to progressive drought stress. Tree Physiol. 2001, 21, 705–715. [Google Scholar] [CrossRef]
- He, X.R.; Wang, X.X.; Fang, J.C.; Zhao, Z.F.; Huang, L.H.; Guo, H.; Zheng, X.H. Bletilla striata: Medicinal uses, phytochemistry and pharmacological activities. J. Ethnopharmacol. 2016, 195, 20–38. [Google Scholar] [CrossRef]
- Kong, L.; Yu, L.; Feng, T.; Yin, X.J.; Liu, T.J.; Dong, L. Physicochemical characterization of the polysaccharide from Bletilla striata: Effect of drying method. Carbohydr. Polym. 2015, 125, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, W.M.; Shi, J.S.; Sun, D.F.; Zhu, C.L. Research progress of Bletila striata polysaccharide gum in medicine. Polym. Bull. 2010, 31, 52–57. [Google Scholar]
- Yu, R.; Wu, Y.; Xing, D. Can electrophysiological parameters substitute for growth, and photosynthetic parameters to characterize the response of mulberry and paper mulberry to drought. Plants 2021, 10, 1772. [Google Scholar] [CrossRef] [PubMed]
- Burdon, S.J. Note on the electrical phenomena which accompany irritation of the leaf of Dionaea muscipula. Proc. R. Soc. Lond. 1873, 21, 495–496. [Google Scholar]
- Li, M.Q.; Li, J.Y.; Mao, H.P.; Wu, Y.Y. Diagnosis and detection of phosphorus nutrition level for Solanum lycopersicum based on electrical impedance spectroscopy. Biosyst. Eng. 2016, 143, 108–118. [Google Scholar]
- Javed, Q.; Wu, Y.Y.; Xing, D.K.; Azeem, A.; Ullah, I.; Zaman, M. Re-watering: An effective measure to recover growth and photosynthetic characteristics in salt-stressed Brassica napus L. Chil. J. Agric. Res. 2017, 77, 78–86. [Google Scholar] [CrossRef]
- Zebelo, S.A.; Maffei, M.E. Plant Electrophysiology: Early Stages of the Plant Response to Chemical Signals//Deciphering Chemical Language of Plant Communication; Springer: Cham, Switzerland, 2016; pp. 285–303. [Google Scholar]
- Javed, Q.; Wu, Y.Y.; Azeem, A.; Ullah, I. Evaluation of irrigation effects using diluted salted water based on electrophysiological properties of plants. J. Plant Interact. 2017, 12, 219–227. [Google Scholar] [CrossRef]
- Fromm, J.; Lautner, S. Electrical signals and their physiological significance in plants. Plant Cell Environ. 2007, 30, 249–257. [Google Scholar] [CrossRef]
- Saraiva, G.F.R.; Ferreira, A.S.; Souza, G.M. Osmotic stress decreases complexity underlying the electrophysiological dynamic in soybean. Plant Biol. 2017, 19, 1031–1037. [Google Scholar] [CrossRef]
- Canales, J.; Henriquez-Valencia, C.; Brauchi, S. The integration of electrical signals originating in the root of vascular plants. Front. Plant Sci. 2018, 8, 2173. [Google Scholar] [CrossRef]
- Saglio, P.H.; Raymond, P.; Pradet, A. metabolic activity and energy charge of excised maize root tips under anoxia. Plant Physiol. 1980, 66, 1053–1057. [Google Scholar] [CrossRef] [PubMed]
- Vanhercke, T.; Tahchy, A.E.; Liu, Q.; Zhou, X.R.; Shrestha, P.; Divi, U.K.; Ral, J.P.; Mansour, M.P. Metabolic engineering of biomass for high energy density: Oilseed-like triacylglycerol yields from plant leaves. Plant Biotechnol. J. 2014, 12, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Shu, S.M.; Duan, X.; Zhao, Y.Y.; Xiong, H.Q. A model based on time, space, energy and iterative mechanism for woody plant metabolic rates and biomass. J. Biobased Mater. Bioenergy 2016, 10, 184–194. [Google Scholar] [CrossRef]
- Yan, X.F.; Wang, Z.Y.; Huang, L.; Wang, C.; Hou, R.F.; Xu, Z.L.; Qiao, X.J. Research progress on electrical signals in higher plants. Prog. Nat. Sci. 2009, 19, 531–541. [Google Scholar] [CrossRef]
- Oyarce, P.; Gurovich, L. Evidence for the transmission of information through electric potentials in injured avocado trees. J. Plant Physiol. 2001, 168, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.M.; Wu, Y.Y.; Xing, D.K.; Zhao, K.; Yu, R. Rapid measurement of drought resistance in plants based on electrophysiological properties. Trans. ASABE. 2015, 58, 1441–1446. [Google Scholar]
- Liu, C.C.; Liu, Y.G.; Fan, D.Y.; Guo, K. Plant drought tolerance assessment for re-vegetation in heterogeneous karst landscapes of southwestern China. Flora Morphol. Distrib. Funct. Ecol. Plants 2012, 207, 30–38. [Google Scholar] [CrossRef]
- Castaeda, V.; González, E.M. Strategies to Apply Water-Deficit Stress: Similarities and Disparities at the Whole Plant Metabolism Level in Medicago truncatula. Int. J. Mol. Sci. 2021, 22, 2813. [Google Scholar] [CrossRef]
- Bhargava, S.; Sawant, K. Drought stress adaptation: Metabolic adjustment and regulation of gene expression. Plant Breed. 2013, 132, 21–32. [Google Scholar] [CrossRef]
- Lourkistia, R.; Oustrica, J.; Quilichinib, Y.; Froelicherc, Y.; Herbetted, S.; Morillone, R.; Bertia, L.; Santinia, J. Improved response of triploid citrus varieties to water deficit is related to anatomical and cytological properties. Plant Physiol. Biochem. 2021, 162, 762–775. [Google Scholar] [CrossRef]
- Li, Y.; Zhao, H.X.; Duan, B.L.; Korpelainen, H.; Li, C.Y. Effect of drought and ABA on growth, photosynthesis and antioxidant system of Cotinus coggygria seedlings under two different light conditions. Environ. Exp Bot. 2011, 71, 107–113. [Google Scholar] [CrossRef]
- Koffler, B.E.; Luschin-Ebengreuth, N.; Stabentheiner, E.; Müller, M.; Zechmann, B. Compartment specific response of antioxidants to drought stress in Arabidopsis. Plant Sci. 2014, 227, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, F.K.; Rivero, R.M.; Blumwald, E.; Mittler, R. Reactive oxygen species, abiotic stress and stress combination. Plant J. 2017, 90, 856–867. [Google Scholar] [CrossRef] [PubMed]
- Srivalli, B.; Sharma, G.; Khanna-chopra, R. Antioxidative defense system in an upland rice cultivar subjected to increasing intensity of water stress followed by recovery. Physiol. Plant. 2003, 119, 503–512. [Google Scholar] [CrossRef]
- Kadioglu, A.; Saruhan, N.; Sağlam, A.; Terzi, R.; Acet, T. Exogenous salicylic acid alleviates effects of long term drought stress and delays leaf rolling by inducing antioxidant system. Plant Growth Regul. 2011, 64, 27–37. [Google Scholar] [CrossRef]
- Lawson, T.; Oxborough, K.; Morison, J.I.L.; Baker, N.R. The responses of guard and mesophyll cell photosynthesis to CO2, O2, light, and water stress in a range of species are similar. J. Exp. Bot. 2003, 54, 1743–1752. [Google Scholar] [CrossRef] [Green Version]
- Redondo, F.J.; Peña, T.C.D.L.; Morcillo, C.N.; Lucas, M.M.; Pueyo, J.J. Overexpression of flavodoxin in bacteroids induces changes in antioxidant metabolism leading to delayed senescence and starch accumulation in alfalfa root nodules. Plant Physiol. 2009, 149, 1166–1178. [Google Scholar] [CrossRef]
- Vieira, E.A.; Silva, M.D.G.; Moro, C.F.; Laura, V.A. Physiological and biochemical changes attenuate the effects of drought on the Cerrado species Vatairea macrocarpa (Benth.) Ducke. Plant Physiol. Biochem. 2017, 115, 472–483. [Google Scholar] [CrossRef]
- Taylor, S.H.; Franks, P.J.; Hulme, S.P.; Spriggs, E.; Christin, P.A.; Edwards, E.J.; Woodward, F.I.; Osborne, C.P. Photosynthetic pathway and ecological adaptation explain stomatal trait diversity amongst grasses. New Phytol. 2012, 193, 387–396. [Google Scholar] [CrossRef]
- Zhou, Y.; Huang, L.H.; Wei, X.L.; Zhou, H.Y.; Chen, X. Physiological, morphological, and anatomical changes in Rhododendron agastum in response to shading. Plant Growth Regul. 2017, 81, 23–30. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Ding, J.Q.; Wang, H.; Su, L.; Zhao, C.C. Biochar addition alleviate the negative effects of drought and salinity stress on soybean productivity and water use efficiency. BMC Plant Biol. 2020, 20, 288. [Google Scholar] [CrossRef] [PubMed]
- Durand, M.; Brendel, O.; Buré, C.; Thiec, D.L. Changes in irradiance and vapour pressure deficit under drought induce distinct stomatal dynamics between glasshouse and field-grown poplars. New Phytol. 2020, 227, 392–406. [Google Scholar] [CrossRef] [PubMed]
- Bhusal, N.; Kim, H.S.; Han, S.G.; Yoon, T.M. Photosynthetic traits and plant–water relations of two apple cultivars grown as bi-leader trees under long-term waterlogging conditions. Environ. Exp. Bot. 2020, 176, 104111. [Google Scholar] [CrossRef]
- Cowan, I.R.; Farquhar, G.D. Stomatal function in relation to leaf metabolism and environment: Stomatal function in the regulation of gas exchange. Symp. Soc. Exp. Biol. 1977, 31, 471–505. [Google Scholar]
- Egilla, J.N.; Davies, F.T.; Boutton, T.W. Drought stress influences leaf water content, photosynthesis, and water-use efficiency of Hibiscus rosa-sinensis at three potassium concentrations. Photosynthetica 2005, 43, 135–140. [Google Scholar] [CrossRef]
- Wu, Y.Y.; Wu, Y.S.; Fang, L.; Wu, M.K.; Wang, R.; Su, Y.; Wang, S.J.; Liu, C.Q. A Method for Measuring Metabolic Energy of Plant Leaf Cells. China Patent CN109060886A, 21 December 2018. [Google Scholar]
- Thongsook, T.; Barrett, D.M. Heat inactivation and reactivation of broccoli peroxidase. J. Agric. Food Chem. 2005, 53, 3215–3222. [Google Scholar] [CrossRef]
- Mou, D.L.; Yao, Y.N.; Yang, Y.Q.; Zhang, Y.M.; Tian, C.Y.; Achal, V. Plant high tolerance to excess manganese related with root growth, manganese distribution and antioxidative enzyme activity in three grape cultivars. Ecotoxicol. Environ. Saf. 2011, 74, 776–786. [Google Scholar] [CrossRef] [PubMed]
- Güneş, A.; Kordali, Ş.; Turan, M.; Bozhüyük, A.U. Determination of antioxidant enzyme activity and phenolic contents of some species of the Asteraceae family from medicanal plants. Ind. Crops Prod. 2019, 137, 208–213. [Google Scholar] [CrossRef]
- Ábrahám, E.; Hourton-Cabassa, C.; Erdei, L.; Szabados, L. Methods for determination of proline in plants. Plant Stress Toler. 2010, 639, 317–331. [Google Scholar]
- Loreto, F.; Velikova, V. Isoprene produced by leaves protects the photosynthetic apparatus against ozone damage, quenches ozone products, and reduces lipid peroxidation of cellular membranes. Plant Physiol. 2001, 127, 1781–1787. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated. Chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
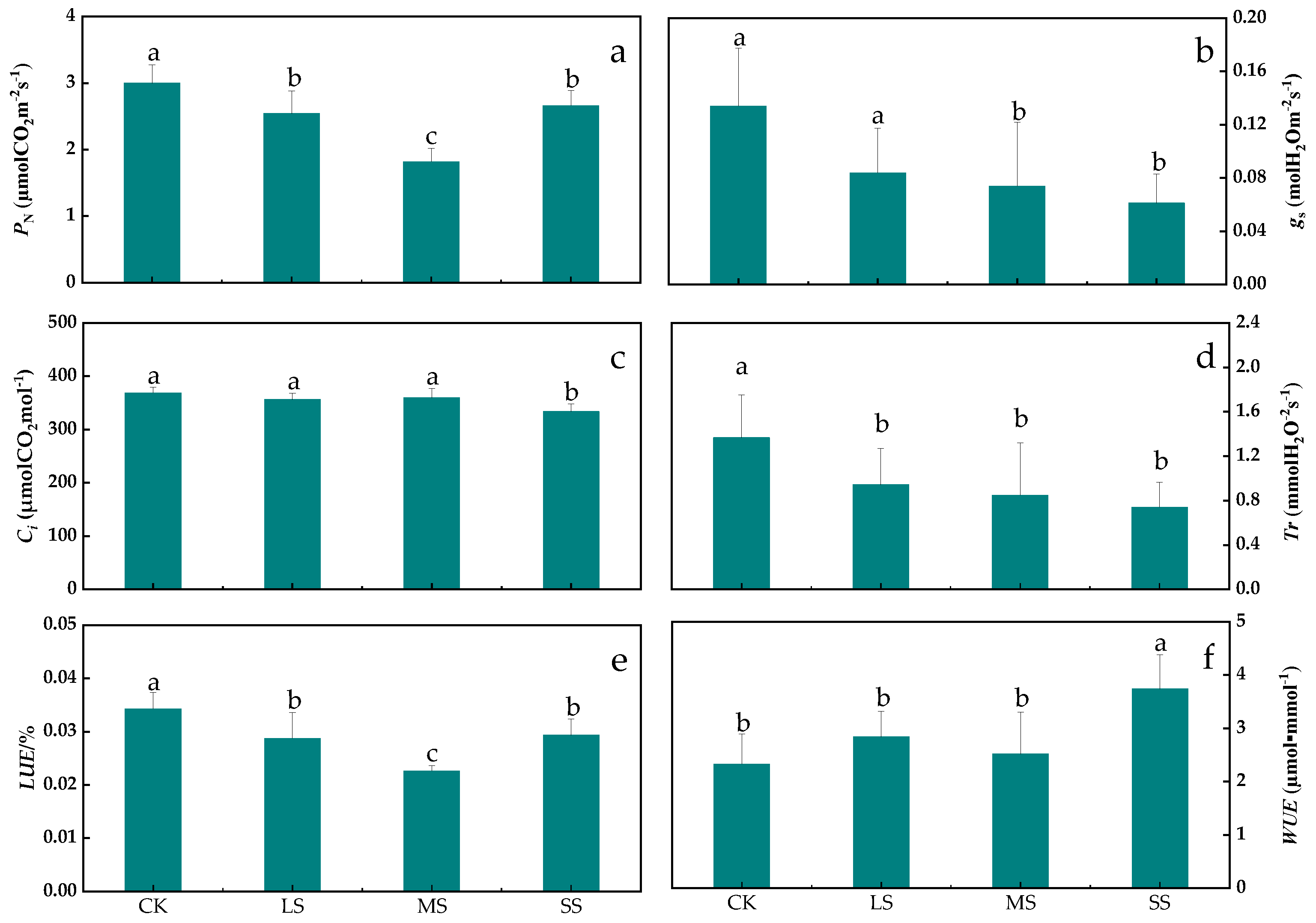{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | x0 | h | Equation |
|---|---|---|---|
| CK | 11.54 | 6.70 | C = 11.54 + 6.70F R2 = 0.99, p < 0.05, n = 76 |
| LS | 3.12 | 9.80 | C = 3.12 + 9.80F R2 = 0.99, p < 0.05, n = 76 |
| MS | 8.06 | 10.28 | C = 8.06 + 10.28F R2 = 0.99, p < 0.05, n = 76 |
| SS | 14.68 | 12.70 | C = 14.68 + 12.70F R2 = 0.98, p < 0.05, n = 76 |
| Treatments | y0 | k1 | b1 | Equation |
|---|---|---|---|---|
| CK | 2.39 | 90.82 | 1.68 | R = 2.39 + 90.82e−1.68F R2 = 0.99, p < 0.05, n = 76 |
| LS | 1.05 | 7.94 | 0.34 | R = 1.05 + 7.94e−0.34F R2 = 0.99, p < 0.05, n = 76 |
| MS | 1.42 | 15.94 | 0.57 | R = 1.42 + 15.94e−0.57F R2 = 0.95, p < 0.05, n = 76 |
| SS | 0.97 | 7.82 | 0.90 | R = 0.97 + 7.82e−0.90F R2 = 0.99, p < 0.05, n = 76 |
| Treatments | p0 | k2 | b2 | Equation |
|---|---|---|---|---|
| CK | 0.91 | 6.06 | 0.69 | Z = 0.91 + 6.06e−0.69F R2 = 0.98, p < 0.05, n = 76 |
| LS | 0.57 | 4.34 | 0.47 | Z = 0.57 + 4.34e−0.47F R2 = 0.99, p < 0.05, n = 76 |
| MS | 0.82 | 3.99 | 0.64 | Z = 0.82 + 3.99e−0.64F R2 = 0.99, p < 0.05, n = 76 |
| SS | 0.50 | 3.89 | 0.86 | Z = 0.50 + 3.89e−0.86F R2 = 0.99, p < 0.05, n = 76 |
| Treatments | d/(10−12m) | ΔGR-E/(1012Jm−1) | ΔGZ-E/(1012Jm−1) | ΔGR/(J) | ΔGZ/(J) | ΔGB/(J) |
|---|---|---|---|---|---|---|
| CK | 7.54 | 2.17 | 2.75 | 16.32 | 20.71 | 18.52 |
| LS | 11.03 | 5.95 | 4.32 | 65.60 | 47.62 | 56.61 |
| MS | 11.57 | 4.24 | 2.47 | 49.06 | 28.59 | 38.83 |
| SS | 14.29 | 2.32 | 2.39 | 33.13 | 34.08 | 33.61 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, Y.; Cai, C.; Yang, Q.; Quan, W.; Li, C.; Wu, Y. Response of Bletilla striata to Drought: Effects on Biochemical and Physiological Parameter Also with Electric Measurements. Plants 2022, 11, 2313. https://doi.org/10.3390/plants11172313
Gao Y, Cai C, Yang Q, Quan W, Li C, Wu Y. Response of Bletilla striata to Drought: Effects on Biochemical and Physiological Parameter Also with Electric Measurements. Plants. 2022; 11(17):2313. https://doi.org/10.3390/plants11172313
Chicago/Turabian StyleGao, Yongdao, Chang Cai, Qiaoan Yang, Wenxuan Quan, Chaochan Li, and Yanyou Wu. 2022. "Response of Bletilla striata to Drought: Effects on Biochemical and Physiological Parameter Also with Electric Measurements" Plants 11, no. 17: 2313. https://doi.org/10.3390/plants11172313