
Physiological and Structural Responses of Olive Leaves Related to Tolerance/Susceptibility to Verticillium dahliae

Abstract
:1. Introduction
2. Results
2.1. Evaluation of Verticillium Wilt of Olive Symptoms
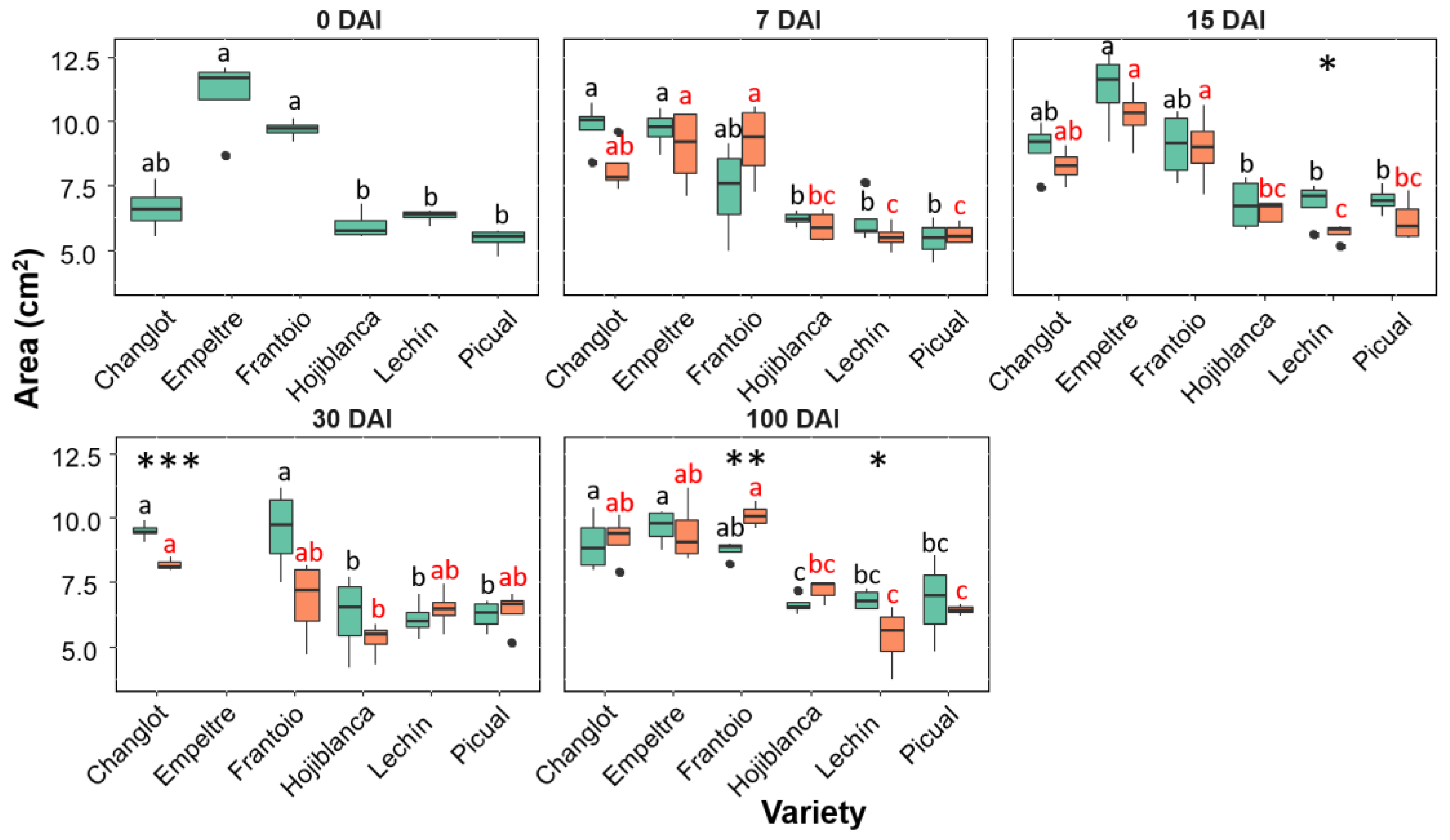
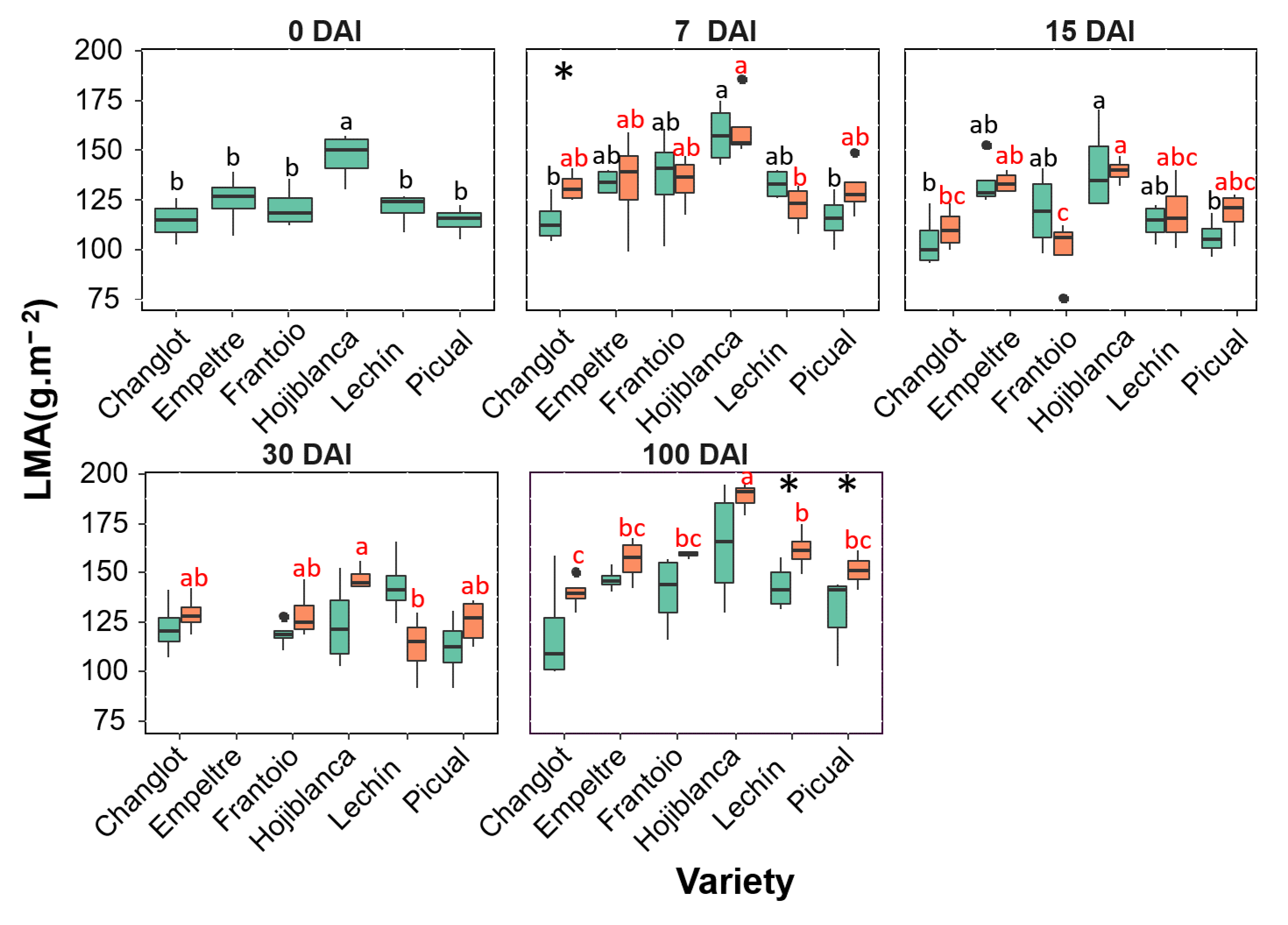
2.2. Variability in Structural Traits between VWO-Susceptible and VWO-Tolerant Cultivars
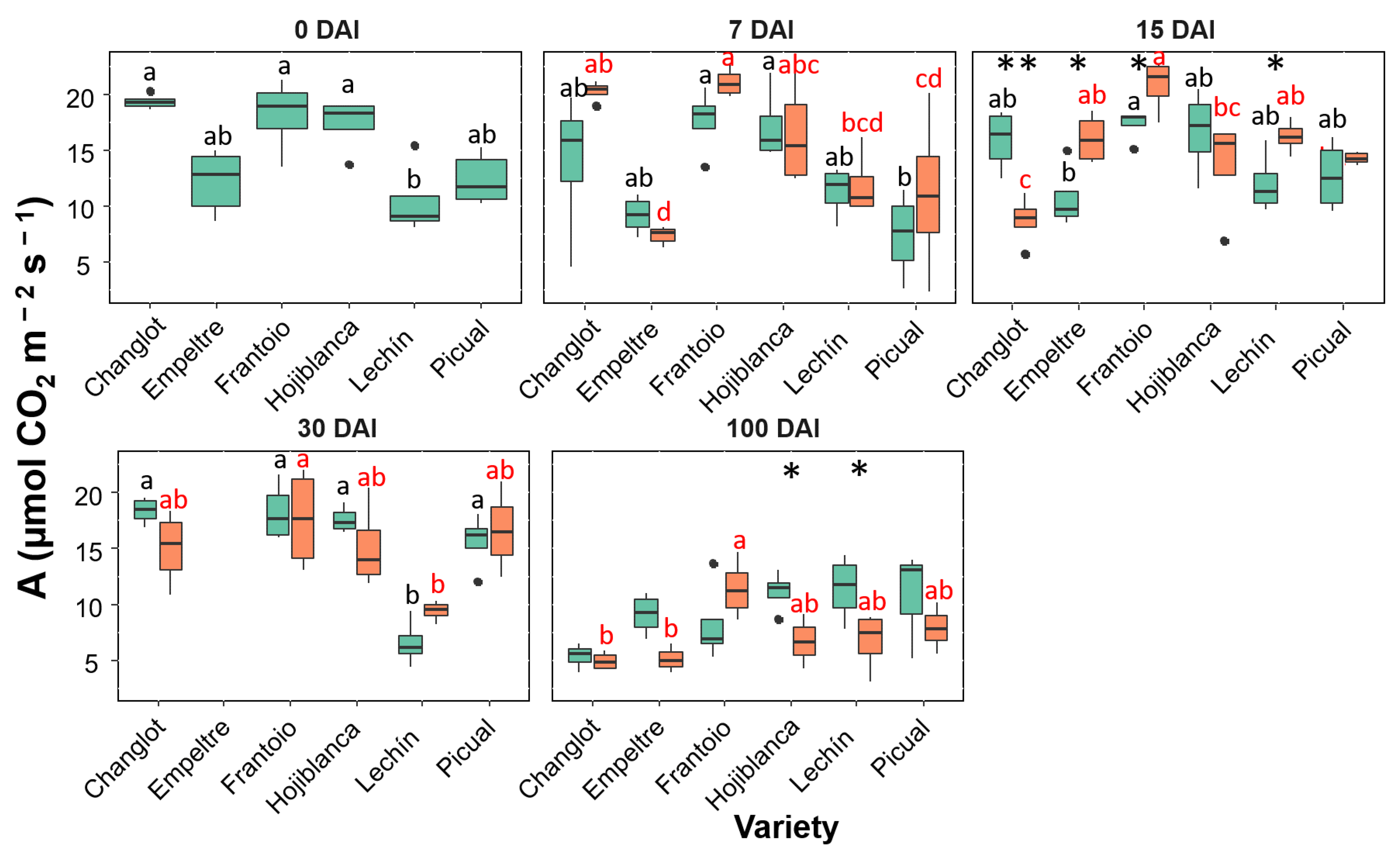
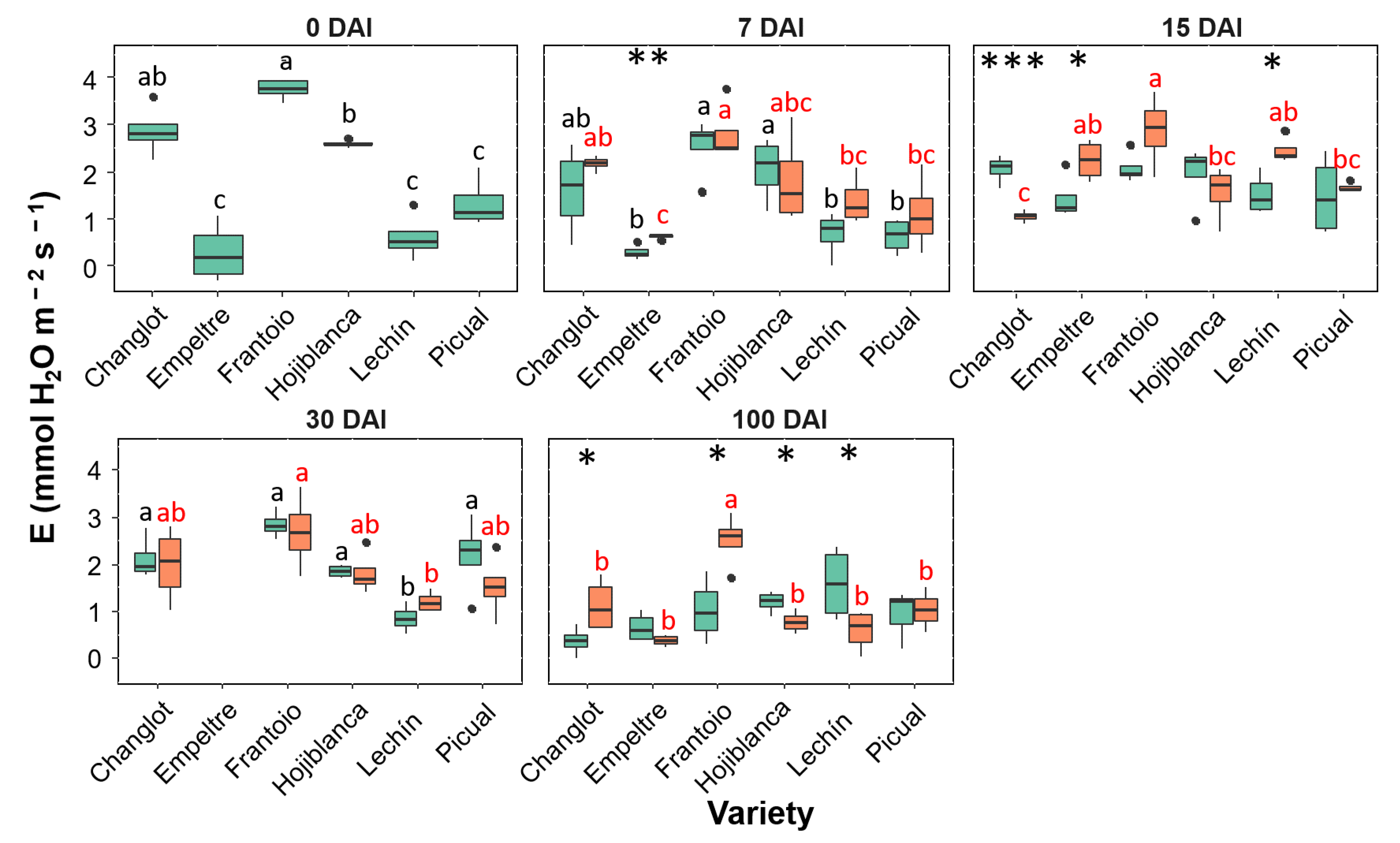
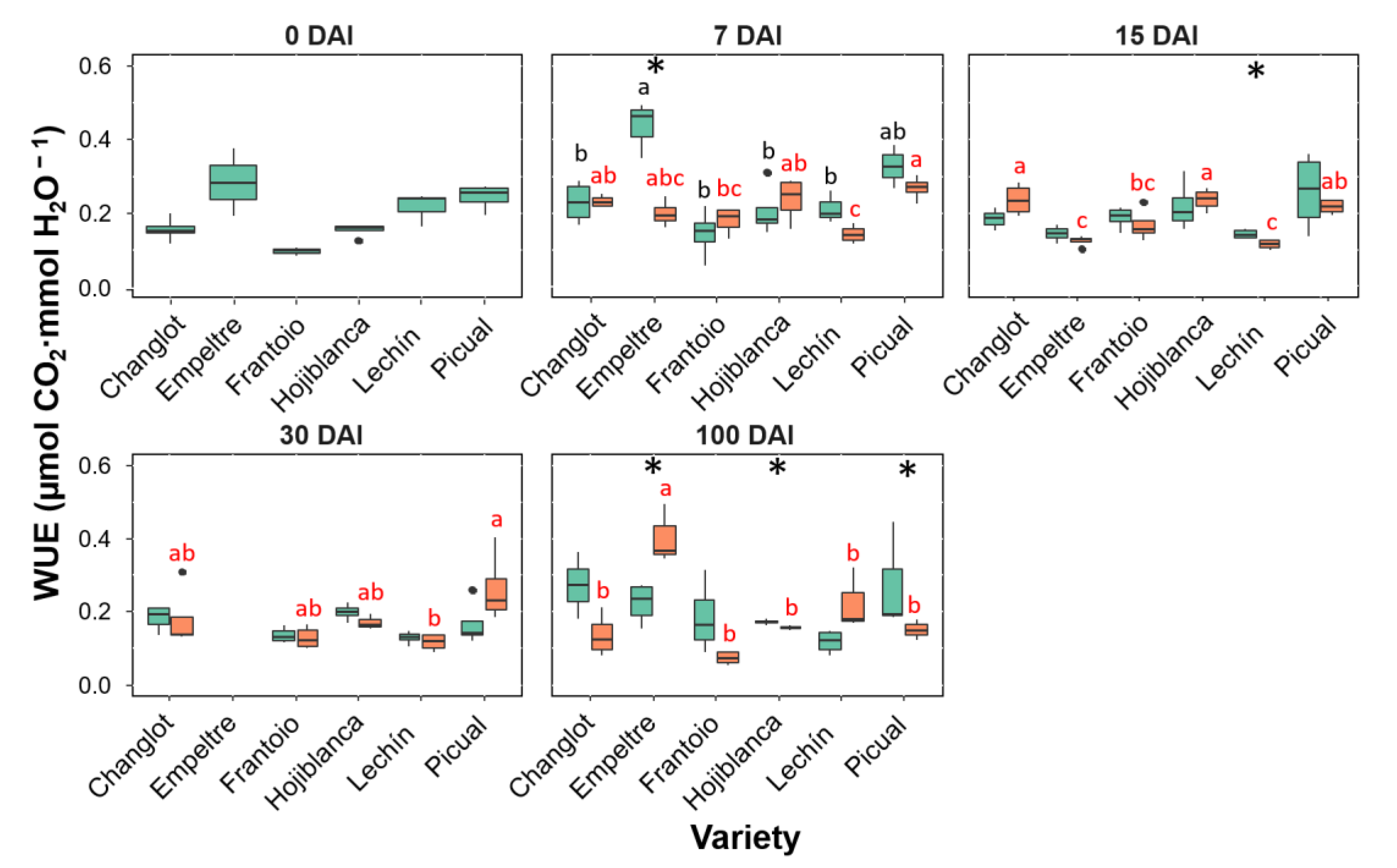
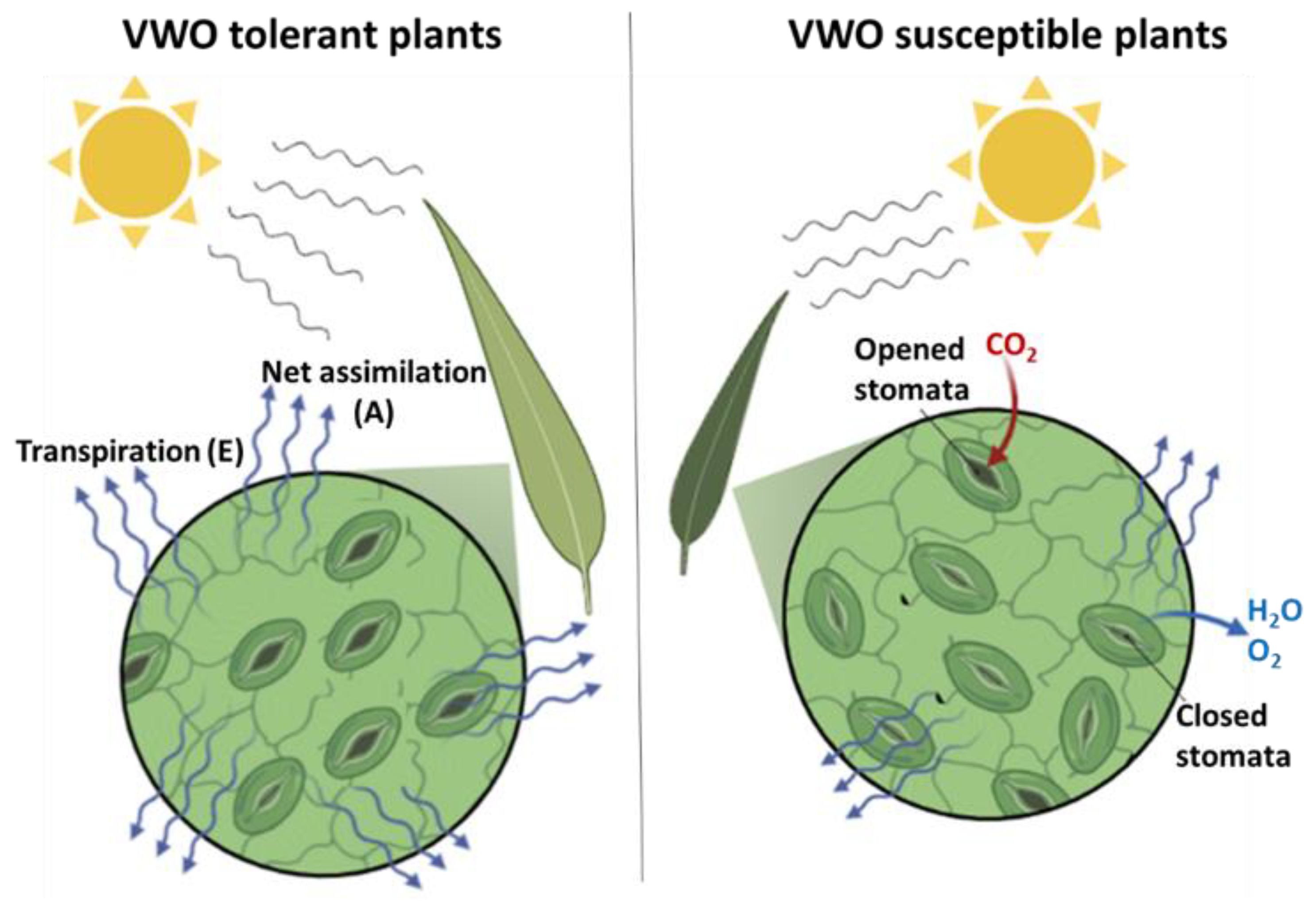
2.3. Variability in Physiological Traits between VWO-Susceptible and VWO-Tolerant Cultivars
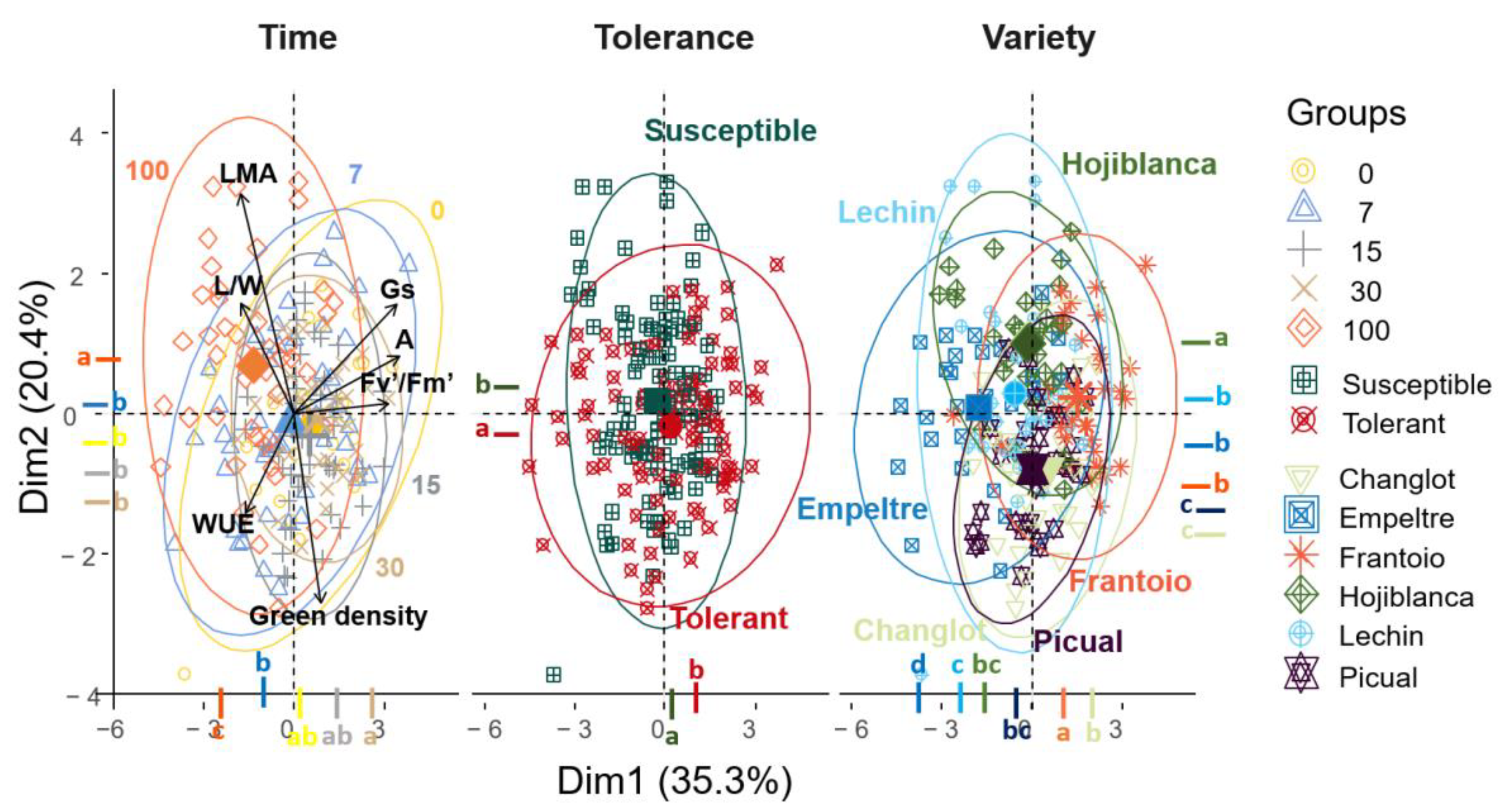
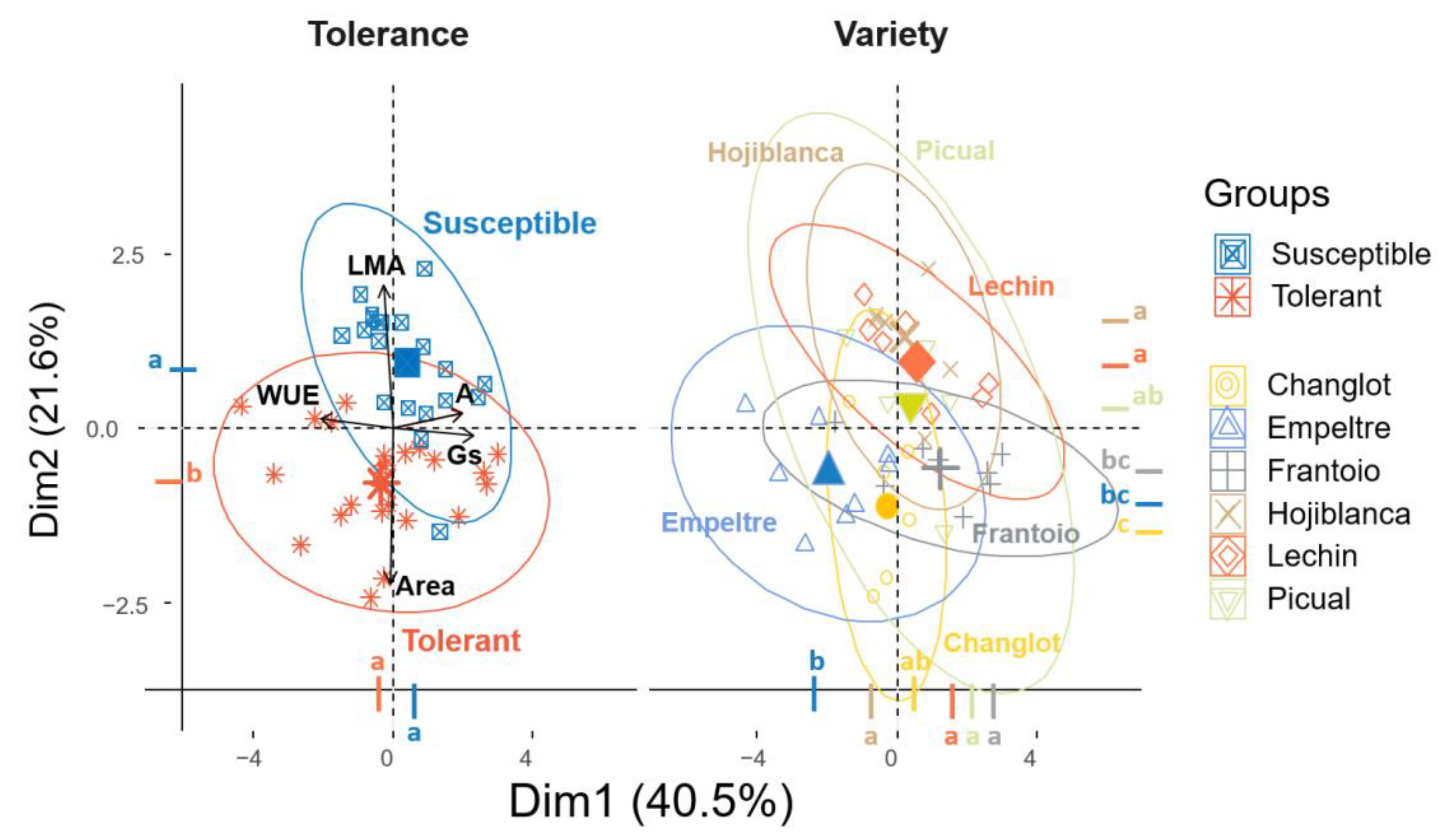
2.4. Contribution of Structural and Physiological Traits to Explain Differences between VWO-Susceptible and VWO-Tolerant Cultivars and among Olive Varieties
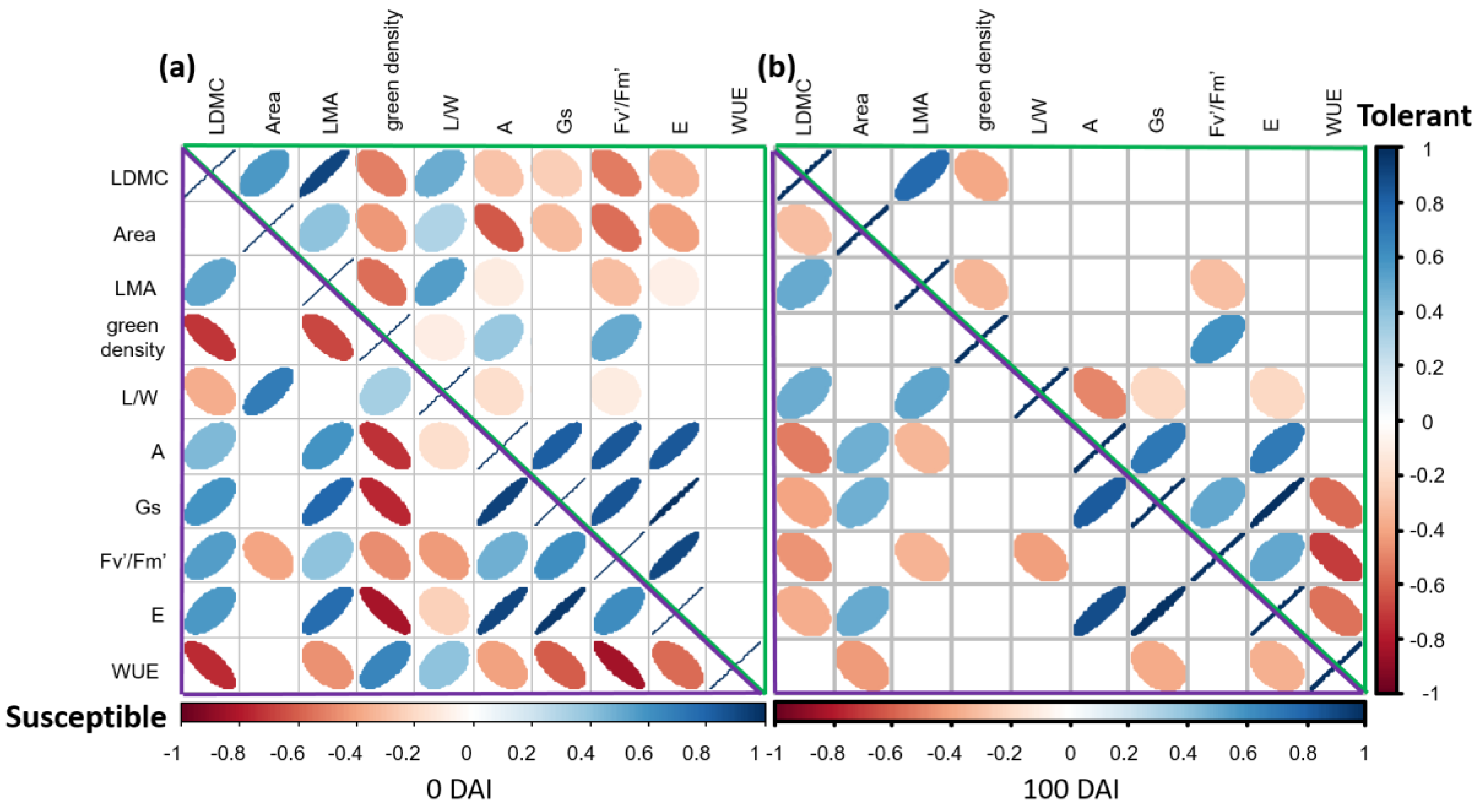
2.5. Differences in the Relationships between Structural and Physiological Traits of Susceptible versus Tolerant Cultivars before and after Inoculation with V. dahliae
3. Discussion
4. Materials and Methods
4.1. Olive Plant Material, Inoculation with V. dahliae and Aboveground Tissue Sampling
4.2. Physiological Functional Traits
4.3. Structural Functional Traits
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pegg, G.F.; Brady, B.L. Verticillium Wilts; CABI: Wallingford, UK, 2002; ISBN 1845933222. [Google Scholar]
- Fradin, E.F.; Thomma, B.P.H.J. Physiology and molecular aspects of Verticillium wilt diseases caused by V. dahliae and V. albo-atrum. Mol. Plant Pathol. 2006, 7, 71–86. [Google Scholar] [CrossRef] [PubMed]
- Montes-Osuna, N.; Mercado-Blanco, J. Verticillium wilt of olive and its control: What did we learn during the last decade? Plants 2020, 9, 735. [Google Scholar] [CrossRef] [PubMed]
- López-Escudero, F.J.; Del Río, C.; Caballero, J.M.; Blanco-López, M.A. Evaluation of olive cultivars for resistance to Verticillium dahliae. Eur. J. Plant Pathol. 2004, 110, 79–85. [Google Scholar] [CrossRef]
- Nigro, A.F.; Gallone, P.; Romanazzi, G.; Schena, L.; Ippolito, A.; Salerno, M.G. Incidence of verticillium wilt on olive in Apulia and genetic diversity of Verticillium dahliae isolates from infected trees. J. Plant Pathol. 2005, 87, 13–23. [Google Scholar]
- Blanco-López, M.A.; Jiménez-Díaz, R.M.; Caballero, J.M. Symptomatology, incidence and distribution of Verticillium wilt of olive trees in Andalucía. Phytopathol. Mediterr. 1984, 23, 1–8. [Google Scholar]
- López-Escudero, F.J.; Mercado-Blanco, J. Verticillium wilt of olive: A case study to implement an integrated strategy to control a soil-borne pathogen. Plant Soil 2011, 344, 1–50. [Google Scholar] [CrossRef]
- Scholz, S.S.; Schmidt-Heck, W.; Guthke, R.; Furch, A.C.U.; Reichelt, M.; Gershenzon, J.; Oelmüller, R. Verticillium dahliae-Arabidopsis interaction causes changes in gene expression profiles and jasmonate levels on different time scales. Front. Microbiol. 2018, 9, 1–19. [Google Scholar] [CrossRef]
- Sancho-Adamson, M.; Trillas, M.I.; Bort, J.; Fernández-Gallego, J.A.; Romanyà, J. Use of rgb vegetation indexes in assessing early effects of verticillium wilt of olive in asymptomatic plants in high and low fertility scenarios. Remote Sens. 2019, 11, 607. [Google Scholar] [CrossRef]
- Dervis, S.; Mercado-Blanco, J.; Erten, L.; Valverde-Corredor, A.; Pérez-Artés, E. Verticillium wilt of olive in Turkey: A survey on disease importance, pathogen diversity and susceptibility of relevant olive cultivars. Eur. J. Plant Pathol. 2010, 127, 287–301. [Google Scholar] [CrossRef]
- Bowden, R.L.; Rouse, D.I.; Sharkey, T.D. Mechanism of photosynthesis decrease by Verticillium dahliae in potato. Plant Physiol. 1990, 94, 1048–1055. [Google Scholar] [CrossRef]
- Sadras, V.O.; Quiroz, F.; Echarte, L.; Escande, A.; Pereyra, V.R. Effect of Verticillium dahliae on photosynthesis, leaf expansion and senescence of field-grown sunflower. Ann. Bot. 2000, 86, 1007–1015. [Google Scholar] [CrossRef]
- Pascual, I.; Azcona, I.; Morales, F.; Aguirreolea, J.; Sánchez-Díaz, M. Photosynthetic response of pepper plants to wilt induced by Verticillium dahliae and soil water deficit. J. Plant Physiol. 2010, 167, 701–708. [Google Scholar] [CrossRef] [PubMed]
- Ayele, A.G.; Wheeler, T.A.; Dever, J.K. Impacts of verticillium wilt on photosynthesis rate, lint production, and fiber quality of greenhouse-grown cotton (Gossypium hirsutum). Plants 2020, 9, 857. [Google Scholar] [CrossRef]
- Zarco-Tejada, P.J.; Poblete, T.; Camino, C.; González-Dugo, V.; Calderón, R.; Hornero, A.; Hernández-Clemente, R.; Román-Écija, M.; Velasco-Amo, M.P.; Landa, B.B.; et al. Divergent abiotic spectral pathways unravel pathogen stress signals across species. Nat. Commun. 2021, 12, 6088. [Google Scholar] [CrossRef]
- Bowden, R.L. Effects of Verticillium dahliae on gas exchange of potato. Phytopathology 1991, 81, 293–301. [Google Scholar] [CrossRef]
- Giorio, P.; Sorrentino, G.; D’Andria, R. Stomatal behaviour, leaf water status and photosynthetic response in field-grown olive trees under water deficit. Environ. Exp. Bot. 1999, 42, 95–104. [Google Scholar] [CrossRef]
- Bruno, G.L.; Sermani, S.; Triozzi, M.; Tommasi, F. Physiological response of two olive cultivars to secondary metabolites of Verticillium dahliae Kleb. Plant Physiol. Biochem. 2020, 151, 292–298. [Google Scholar] [CrossRef] [PubMed]
- Petridis, A.; Therios, I.; Samouris, G.; Koundouras, S.; Giannakoula, A. Effect of water deficit on leaf phenolic composition, gas exchange, oxidative damage and antioxidant activity of four Greek olive (Olea europaea L.) cultivars. Plant Physiol. Biochem. 2012, 60, 1–11. [Google Scholar] [CrossRef]
- Montes-Osuna, N.; Gómez-Lama Cabanás, C.; Valverde-Corredor, A.; Legarda, G.; Prieto, P.; Mercado-Blanco, J. Evaluation of indigenous olive biocontrol rhizobacteria as protectants against drought and salt stress. Microorganisms 2021, 9, 1209. [Google Scholar] [CrossRef]
- Ibaraki, Y.; Murakami, J. Distribution of chlorophyll fluorescence parameter Fv/Fm within individual plants under various stress conditions. Acta Hortic. 2007, 761, 255–260. [Google Scholar] [CrossRef]
- Inamullah; Isoda, A. Adaptive responses of soybean and cotton to water stress II. Changes in CO2 assimilation rate, chlorophyll fluorescence and photochemical reflectance index in relation to leaf temperature. Plant Prod. Sci. 2005, 8, 131–138. [Google Scholar] [CrossRef]
- Hampton, R.E.; Wullschleger, S.D.; Oosterhuis, D.M. Impact of verticillium wilt on net photosynthesis, respiration and photorespiration in field-grown cotton (Gossypium hirsutum L.). Physiol. Mol. Plant Pathol. 1990, 37, 271–280. [Google Scholar] [CrossRef]
- Saeed, I.A.M.; MacGuidwin, A.E.; Rouse, D.I.; Sharkey, T.D. Limitation to photosynthesis in Pratylenchus penetrans- and Verticillium dahliae-infected potato. Crop Sci. 1999, 39, 1340–1346. [Google Scholar] [CrossRef]
- Bosabalidis, A.M.; Kofidis, G. Comparative effects of drought stress on leaf anatomy of two olive cultivars. Plant Sci. 2002, 163, 375–379. [Google Scholar] [CrossRef]
- García-Ruiz, G.M.; Trapero, C.; Del Río, C.; López-Escudero, F.J. Evaluation of resistance of Spanish olive cultivars to Verticillium dahliae in inoculations conducted in greenhouse. Phytoparasitica 2014, 42, 205–212. [Google Scholar] [CrossRef]
- Arias-Calderón, R.; Rodríguez-Jurado, D.; Bejarano-Alcázar, J.; Belaj, A.; de la Rosa, R.; León, L. Evaluation of Verticillium wilt resistance in selections from olive breeding crosses. Euphytica 2015, 206, 619–629. [Google Scholar] [CrossRef]
- Valverde, P.; Trapero, C.; Arquero, O.; Serrano, N.; Barranco, D.; Muñoz Díez, C.; López-Escudero, F.J. Highly infested soils undermine the use of resistant olive rootstocks as a control method of verticillium wilt. Plant Pathol. 2021, 70, 144–153. [Google Scholar] [CrossRef]
- Trapero, C.; Serrano, N.; Arquero, O.; Del Río, C.; Trapero, A.; López-Escudero, F.J. Field resistance to Verticillium wilt in selected olive cultivars grown in two naturally infested soils. Plant Dis. 2012, 97, 668–674. [Google Scholar] [CrossRef]
- Gómez-Lama Cabanás, C.; Schilirò, E.; Valverde-Corredor, A.; Mercado-Blanco, J. Systemic responses in a tolerant olive (Olea europaea L.) cultivar upon root colonization by the vascular pathogen Verticillium dahliae. Front. Microbiol. 2015, 6, 928. [Google Scholar] [CrossRef]
- Leyva-Pérez, M.O.; Jiménez-Ruiz, J.; Gómez-Lama Cabanás, C.; Valverde-Corredor, A.; Barroso, J.B.; Luque, F.; Mercado-Blanco, J. Tolerance of olive (Olea europaea) cv Frantoio to Verticillium dahliae relies on both basal and pathogen-induced differential transcriptomic responses. New Phytol. 2018, 217, 671–686. [Google Scholar]
- Robb, J. Verticillium tolerance: Resistance, susceptibility, or mutualism? Can. J. Bot. 2007, 85, 903–910. [Google Scholar] [CrossRef]
- Prieto, P.; Navarro-Raya, C.; Valverde-Corredor, A.; Amyotte, S.G.; Dobinson, K.F.; Mercado-Blanco, J. Colonization process of olive tissues by Verticillium dahliae and its in planta interaction with the biocontrol root endophyte Pseudomonas fluorescens PICF7. Microb. Biotechnol. 2009, 2, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Ruiz, J.; Ramírez-Tejero, J.A.; Fernández-Pozo, N.; de la O Leyva-Pérez, M.; Yan, H.; de la Rosa, R.; Belaj, A.; Montes, E.; Rodríguez-Ariza, M.O.; Navarro, F.; et al. Transposon activation is a major driver in the genome evolution of cultivated olive trees (Olea europaea L.). Plant Genome 2020, 13, e20010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández-González, A.J.; Cardoni, M.; Gómez-Lama Cabanás, C.; Valverde-Corredor, A.; Villadas, P.J.; Fernández-López, M.; Mercado-Blanco, J. Linking belowground microbial network changes to different tolerance level towards Verticillium wilt of olive. Microbiome 2020, 8, 11. [Google Scholar] [CrossRef]
- Navas-Cortés, J.A.; Landa, B.B.; Mercado-Blanco, J.; Trapero-Casas, J.L.; Rodríguez-Jurado, D.; Jiménez-Díaz, R.M. Spatiotemporal analysis of spread of infections by Verticillium dahliae pathotypes within a high tree density olive orchard in Southern Spain. Phytopathology 2008, 98, 167–180. [Google Scholar] [CrossRef]
- Calderón, R.; Navas-Cortés, J.A.; Lucena, C.; Zarco-Tejada, P.J. High-resolution airborne hyperspectral and thermal imagery for early detection of Verticillium wilt of olive using fluorescence, temperature and narrow-band spectral indices. Remote Sens. Environ. 2013, 139, 231–245. [Google Scholar] [CrossRef]
- Cardoni, M.; Mercado-Blanco, J.; Villar, R. Functional traits of olive varieties and their relationship with the tolerance level towards verticillium wilt. Plants 2021, 10, 1079. [Google Scholar] [CrossRef]
- Díaz-Espejo, A.; Walcroft, A.S.; Fernández, J.E.; Hafidi, B.; Palomo, M.J.; Girón, I.F. Modeling photosynthesis in olive leaves under drought conditions. Tree Physiol. 2006, 26, 1445–1456. [Google Scholar] [CrossRef]
- Cardoni, M.; Gómez-Lama Cabanás, C.; Valverde-Corredor, A.; Villar, R.; Mercado-Blanco, J. Unveiling differences in root defense mechanisms between tolerant and susceptible olive cultivars to Verticillium dahliae. Front. Plant Sci. 2022, 13, 863055. [Google Scholar] [CrossRef]
- Markakis, E.A.; Tjamos, S.E.; Antoniou, P.P.; Roussos, P.A.; Paplomatas, E.J.; Tjamos, E.C. Phenolic responses of resistant and susceptible olive cultivars induced by defoliating and nondefoliating Verticillium dahliae pathotypes. Plant Dis. 2010, 94, 1156–1162. [Google Scholar] [CrossRef]
- Pandey, S.K.; Singh, H. A simple, cost-effective method for leaf area estimation. J. Bot. 2011, 2011, 658240. [Google Scholar] [CrossRef]
- Johnson, K.B. Agrisexport. Phytopathology 1988, 78, 1198–1205. [Google Scholar] [CrossRef]
- Nogués, S.; Cotxarrera, L.; Alegre, L.; Trillas, M.I. Limitations to photosynthesis in tomato leaves induced by Fusarium wilt. New Phytol. 2002, 154, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Westoby, M.; Falster, D.S.; Moles, A.T.; Vesk, P.A.; Wright, I.J. Plant ecological strategies: Some leading dimensions of variation between species. Annu. Rev. Ecol. Syst. 2002, 33, 125–159. [Google Scholar] [CrossRef]
- Wright, I.J.; Westoby, M. Leaves at low versus high rainfall: Coordination of structure, lifespan and physiology. New Phytol. 2002, 155, 403–416. [Google Scholar] [CrossRef]
- Poorter, H.; Niinemets, Ü.; Poorter, L.; Wright, I.J.; Villar, R. Causes and consequences of variation in leaf mass per area (LMA): A meta-analysis. New Phytol. 2009, 182, 565–588. [Google Scholar] [CrossRef]
- John, G.P.; Scoffoni, C.; Buckley, T.N.; Villar, R.; Poorter, H.; Sack, L. The anatomical and compositional basis of leaf mass per area. Ecol. Lett. 2017, 20, 412–425. [Google Scholar] [CrossRef] [PubMed]
- Shipley, B.; Vu, T.T. Dry matter content as a measure of dry matter concentration in plants and their parts. New Phytol. 2002, 153, 359–364. [Google Scholar] [CrossRef]
- Murchie, E.H.; Lawson, T. Chlorophyll fluorescence analysis: A guide to good practice and understanding some new applications. J. Exp. Bot. 2013, 64, 3983–3998. [Google Scholar] [CrossRef]
- Nogués, S.; Baker, N.R. Effects of drought on photosynthesis in Mediterranean plants grown under enhanced UV-B radiation. J. Exp. Bot. 2000, 51, 1309–1317. [Google Scholar] [CrossRef]
- Scharte, J.; Schön, H.; Weis, E. Photosynthesis and carbohydrate metabolism in tobacco leaves during an incompatible interaction with Phytophthora nicotianae. Plant Cell Environ. 2005, 28, 1421–1435. [Google Scholar] [CrossRef]
- Grimmer, M.K.; Foulkes, M.J.; Pavaley, N.D. Foliar pathogenesis and plant water relations: A review. J. Exp. Bot. 2012, 63, 4321–4331. [Google Scholar] [CrossRef] [PubMed]
- Monsi, M. Uber den Lichtfaktor in den Pflanzen-gesellschaften und seine Bedeutung fur die Stoffproduktion. Jpn. J. Bot. 1953, 14, 22–52. [Google Scholar]
- Lakso, A.N.; Prat, C.; Pearson, R.C.; Pool, R.M.; Seem, R.C.; Welser, M.J. Photosynthesis, transpiration, and water use efficiency of mature grape laves infected with Uncinula necator (Powdery Mildew). Phytopathology 1982, 72, 232–236. [Google Scholar] [CrossRef]
- Andersen, P.C. Impact of pecan leaf blotch on gas exchange of pecan leaves. Plant Dis. 1990, 74, 203–207. [Google Scholar] [CrossRef]
- Junior, W.C.J.; Vale, F.X.R.; Martinez, C.A.; Coelho, R.R.; Costa, L.C.; Hau, B.; Zambolim, L. Effects of angular leaf spot and rust on leaf gas exchange and yield of common bean (Phaseolus vulgaris). Photosynthetica 2001, 39, 603–606. [Google Scholar] [CrossRef]
- Bacon, M. Water Use Efficiency in Plant Biology; John Wiley & Sons: Hoboken, NJ, USA, 2009; ISBN 1-4051-1434-7. [Google Scholar]
- Gremer, J.R.; Kimball, S.; Keck, K.R.; Huxman, T.E.; Angert, A.L.; Venable, D.L. Water-use efficiency and relative growth rate mediate competitive interactions in Sonoran Desert winter annual plants. Am. J. Bot. 2013, 100, 2009–2015. [Google Scholar] [CrossRef]
- Ribeiro, R.V.; Machado, E.C.; Oliveira, R.F. Early photosynthetic responses of sweet orange plants infected with Xylella fastidiosa. Physiol. Mol. Plant Pathol. 2003, 62, 167–173. [Google Scholar] [CrossRef]
- Tomáškováa, I. Water use efficiency of Norway spruce with bud blight disease. Acta Hortic. 2018, 1222, 133–136. [Google Scholar] [CrossRef]
- De Mendiburu, F.; Simon, R. Agricolae—Ten years of an open source statistical tool for experiments in breeding, agriculture and biology. PeerJ 2015, 3, e1404v1. [Google Scholar]
- Kassambara, A. Practical guide to principal component methods in R: PCA, M (CA), FAMD, MFA, HCPC, factoextra. Sthda 2017, 2. Available online: http://www.sthda.com/english/wiki/practical-guide-to-principal-component-methods-in-r (accessed on 5 July 2022).
- Wei, T.; Simko, V.; Levy, M.; Xie, Y.; Jin, Y.; Zemla, J. R package “corrplot”: Visualization of a Correlation Matrix. Statistician 2017, 56, 316–324. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Traits | Factors | Interactions | R2 | |||||
|---|---|---|---|---|---|---|---|---|
| Structural | Variety | Treatment | Time | Variety × Treatment | Time × Variety | Time × Treatment | Time × Variety × Treatment | (×100) |
| Area | 66.29 *** | 0.94 * | 1.67 ** | 0.46 | 7.74 *** | 0.91 | 3.59 ** | 81.59 |
| LMA | 25.85 *** | 1.73 ** | 24.57 *** | 2.12 | 5.03 | 3.01 ** | 3.78 | 68.74 |
| L/W | 33.68 *** | 0.02 | 10.72 *** | 0.82 | 8.27 | 0.91 | 1.46 | 55.89 |
| LDMC | 3.35 * | 0.06 | 15.73 *** | 2.43 | 16.65 ** | 1.45 | 4.33 | 53.57 |
| Physiological | ||||||||
| A | 22.37 *** | 0.00 | 24.02 *** | 1.62 * | 17.77 *** | 1.92 ** | 6.34 ** | 78.62 |
| Gs | 15.19 *** | 0.31 | 27.69 *** | 0.86 | 25.70 *** | 0.18 | 8.84 *** | 78.46 |
| Fv′/Fm′ | 22.07 *** | 0.67 | 7.41 *** | 0.75 | 10.56 ** | 0.53 | 4.89 | 46.89 |
| E | 37.69 *** | 0.46 * | 11.73 *** | 1.32 * | 20.91 *** | 0.52 | 6.15 *** | 78.78 |
| WUE | 4.13 *** | 0.03 | 73.22 *** | 0.29 | 16.18 *** | 0.09 | 1.16 *** | 95.10 |
| Traits | Factors | Interactions | Coefficients of Determination | ||||
|---|---|---|---|---|---|---|---|
| Structural | Variety | Treatment | Tolerance | Variety × Treatment | Treatment × Tolerance | R2 (×100) 1 | R2 (×100) 2 |
| Area | 72.64 *** | 0.02 | 69.89 *** | 6.85 | 1.99 | 79.5 | 71.8 |
| LMA | 40.25 *** | 18.94 *** (+) | 6.3 | 1.53 | 0.54 | 60.72 | 25.8 |
| L/W | 48.96 *** | 2.75 | 31.03 *** | 2.82 | 0.02 | 54.53 | 33.8 |
| LDMC | 11.78 | 11.23 * (+) | 5.91 | 7.32 | 0.78 | 30.33 | 17.9 |
| Physiological | |||||||
| A | 25.31 ** | 13.20 ** (−) | 7.72 * | 19.51 *** | 8.80 | 58.02 | 29.7 |
| Gs | 29.86 *** | 0.08 | 0 | 39.24 *** | 15.22 | 69.18 | 15.3 |
| Fv′/Fm′ | 70.02 *** | 2.44 | 11.10 * | 5.49 | 0.38 | 77.95 | 13.9 |
| E | 30.13 *** | 0.39 | 0.02 | 34 *** | 16.3 | 64.53 | 16.7 |
| WUE | 33.94 *** | 0.004 | 0.28 | 25.14 ** | 3.33 | 59.08 | 3.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cardoni, M.; Quero, J.L.; Villar, R.; Mercado-Blanco, J. Physiological and Structural Responses of Olive Leaves Related to Tolerance/Susceptibility to Verticillium dahliae. Plants 2022, 11, 2302. https://doi.org/10.3390/plants11172302
Cardoni M, Quero JL, Villar R, Mercado-Blanco J. Physiological and Structural Responses of Olive Leaves Related to Tolerance/Susceptibility to Verticillium dahliae. Plants. 2022; 11(17):2302. https://doi.org/10.3390/plants11172302
Chicago/Turabian StyleCardoni, Martina, José Luis Quero, Rafael Villar, and Jesús Mercado-Blanco. 2022. "Physiological and Structural Responses of Olive Leaves Related to Tolerance/Susceptibility to Verticillium dahliae" Plants 11, no. 17: 2302. https://doi.org/10.3390/plants11172302