
The Higher Water Absorption Capacity of Small Root System Improved the Yield and Water Use Efficiency of Maize

 and
and
Abstract
:1. Introduction
2. Results
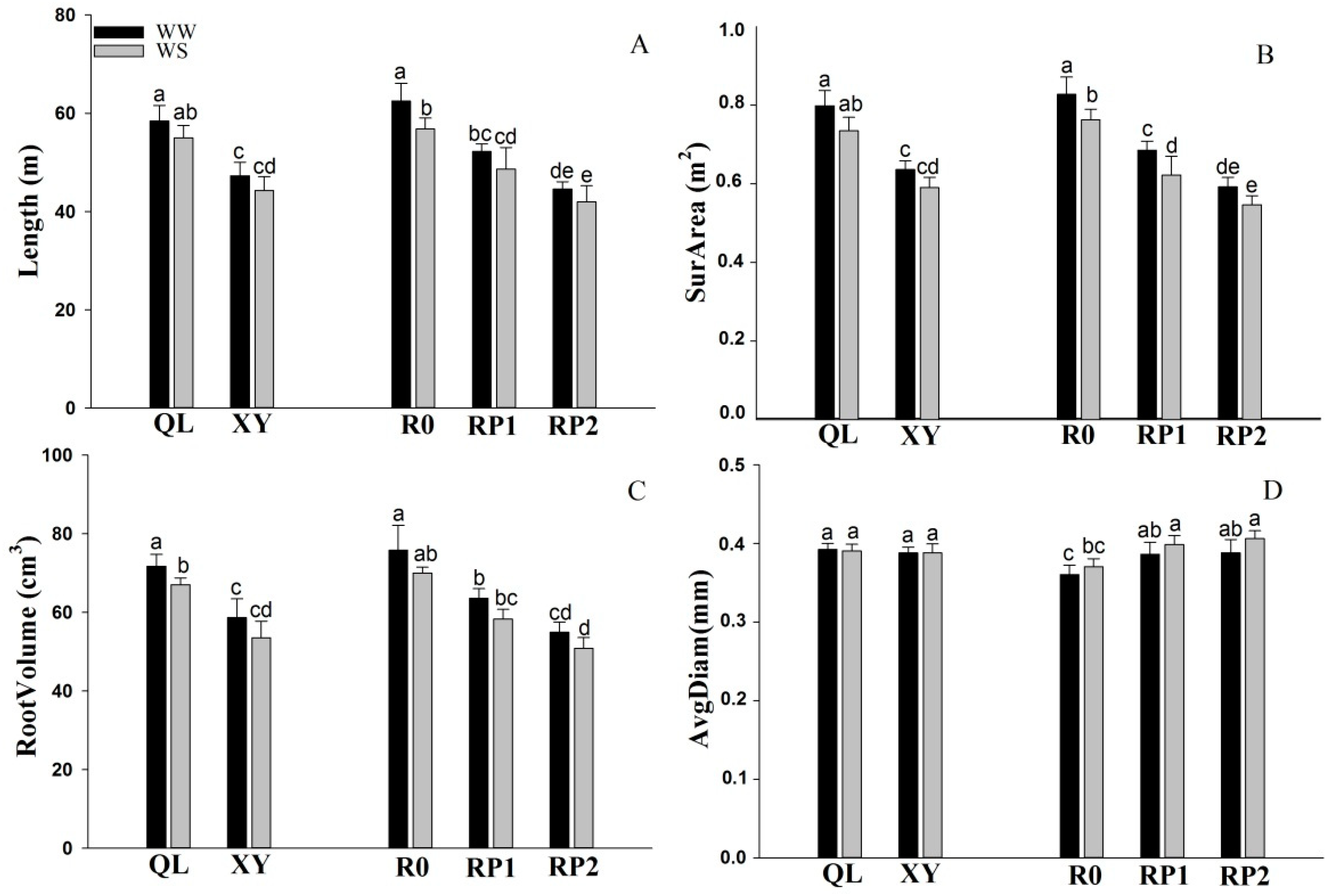
2.1. Root Traits
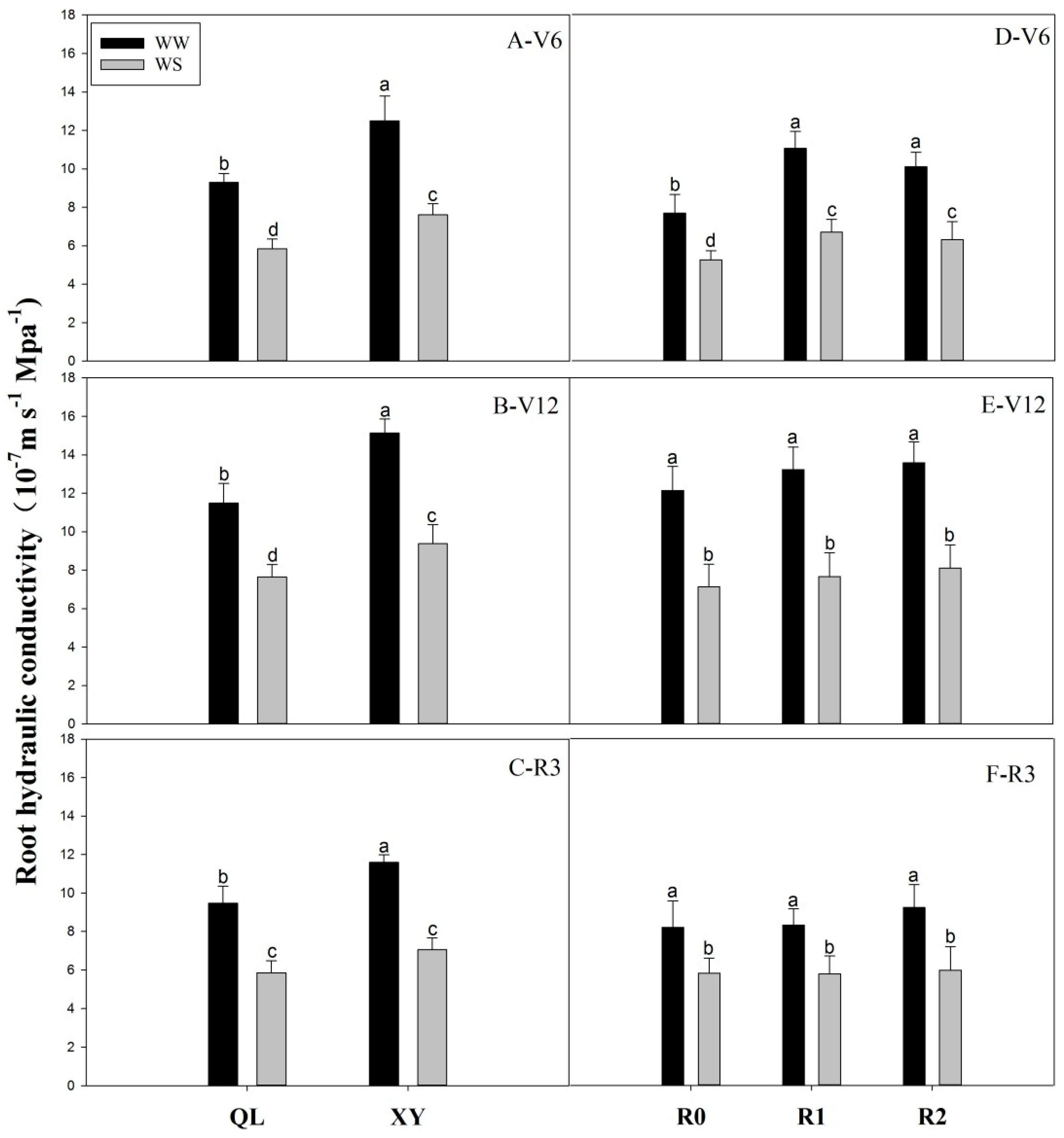
2.2. Root Hydraulic Conductivity
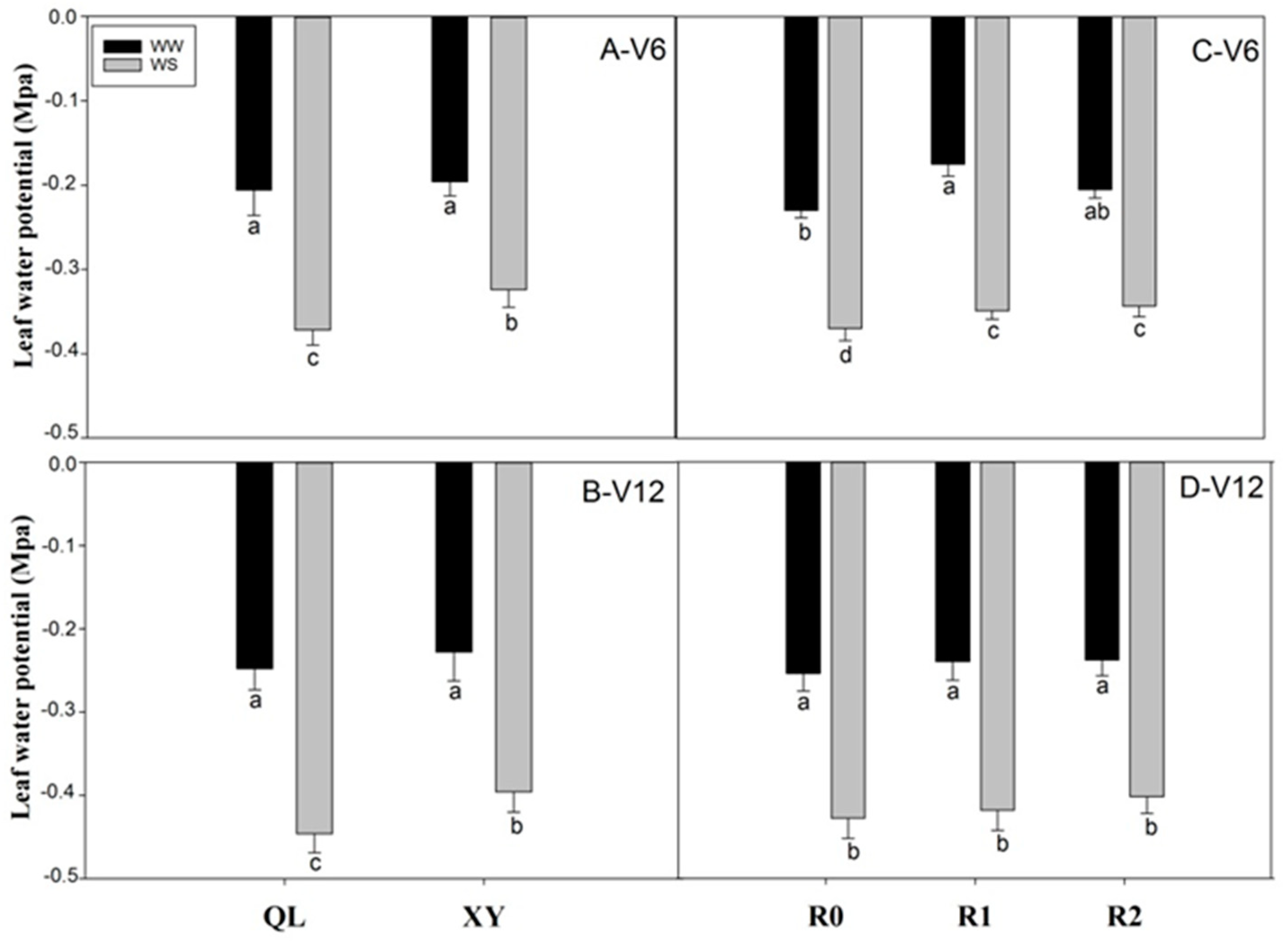
2.3. Leaf Water Potential (Ψleaf)
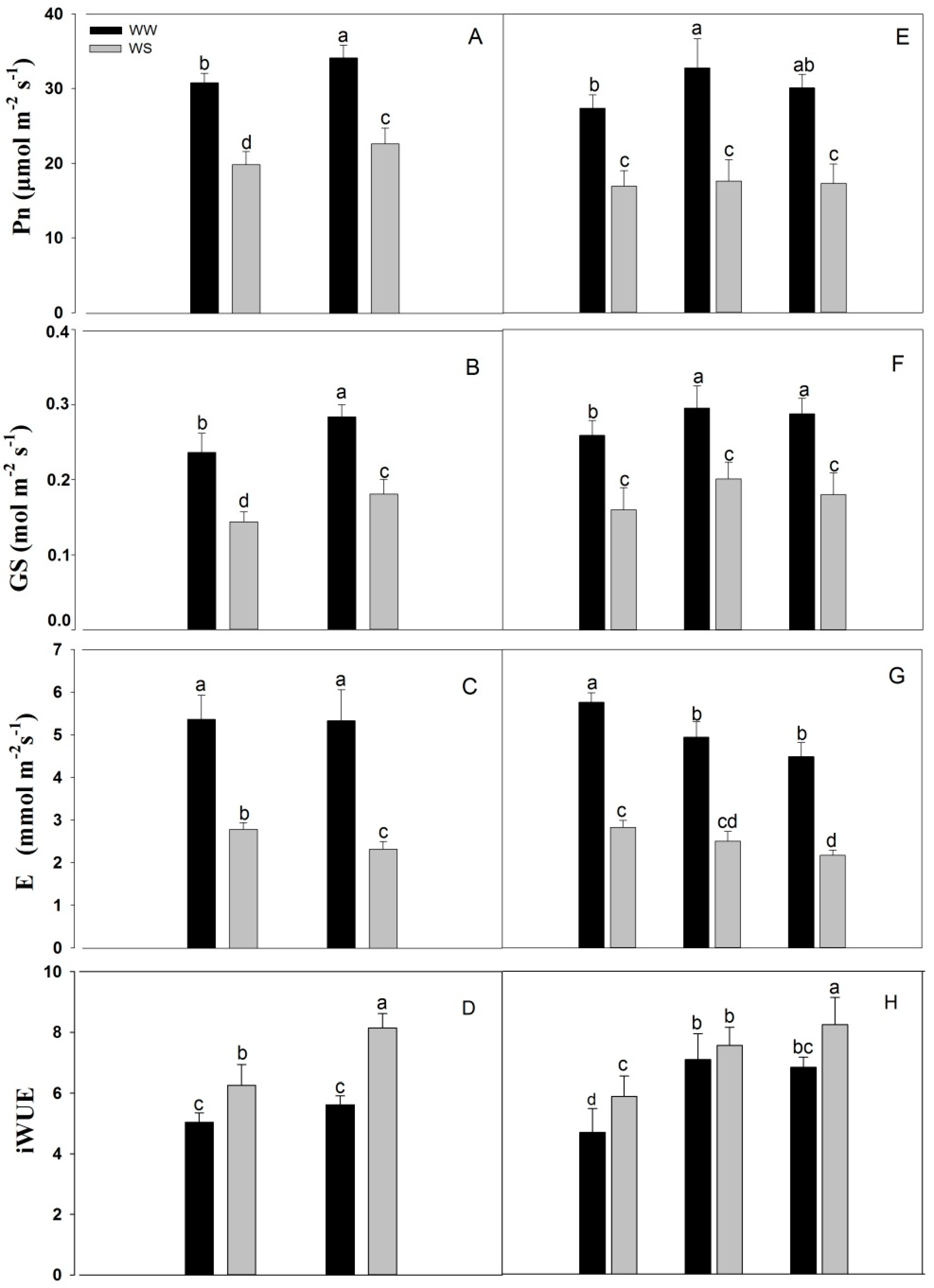
2.4. Leaf Gas Exchange Parameters
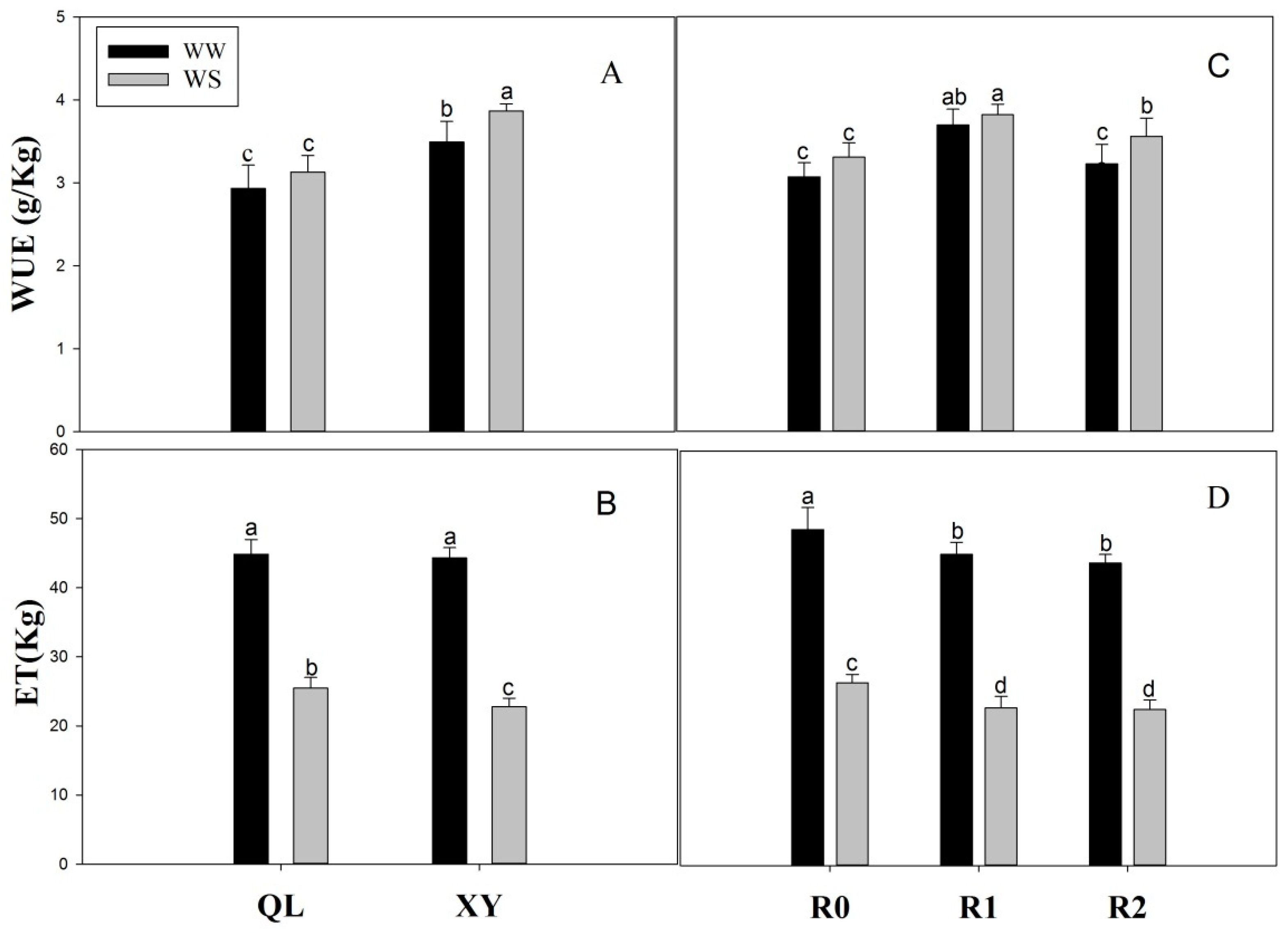
2.5. Grain Yield (GY) and WUE
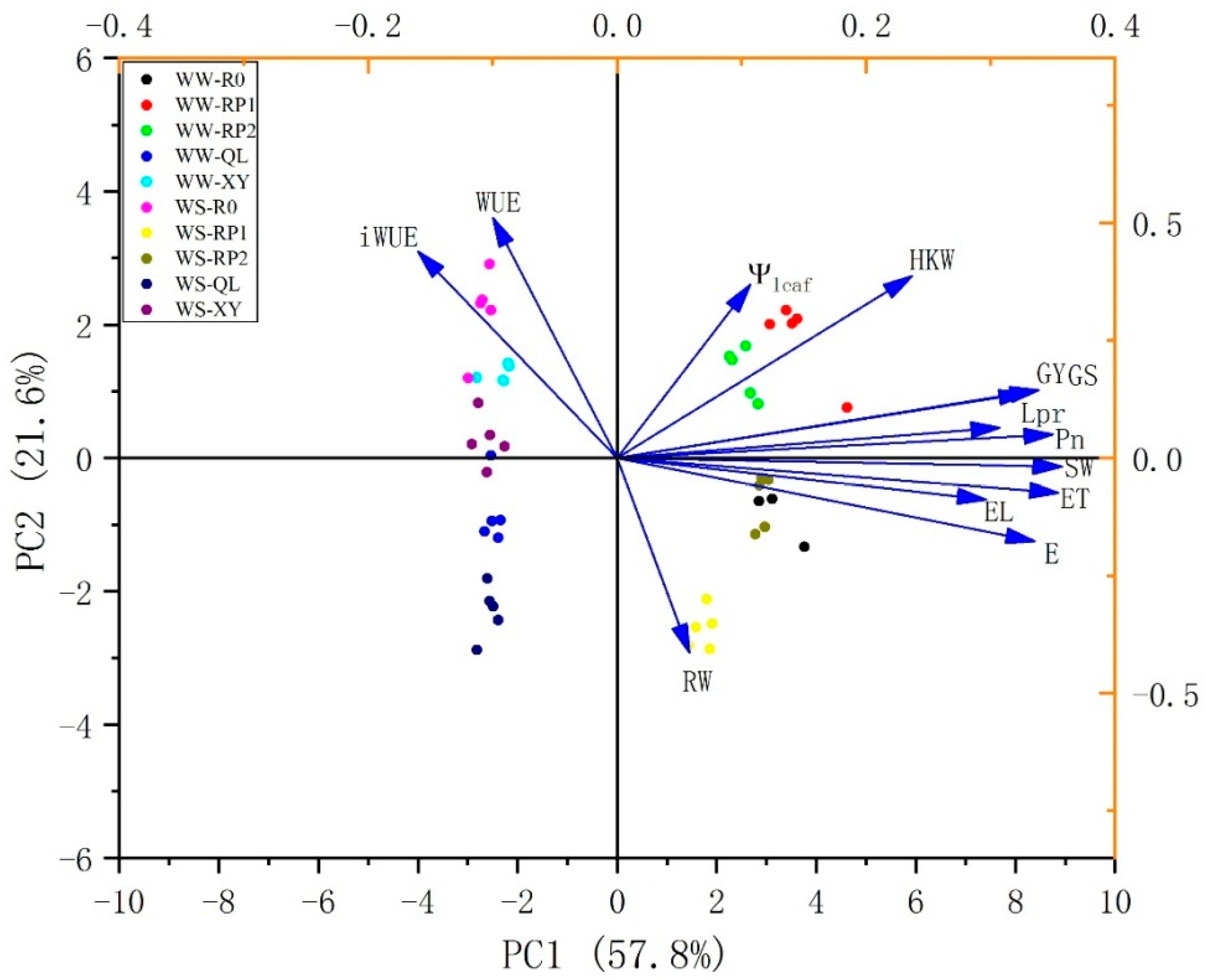
2.6. Principal Component Analysis (PCA) of Maize Growth and Physiological Parameters
3. Discussion
4. Material and Methods
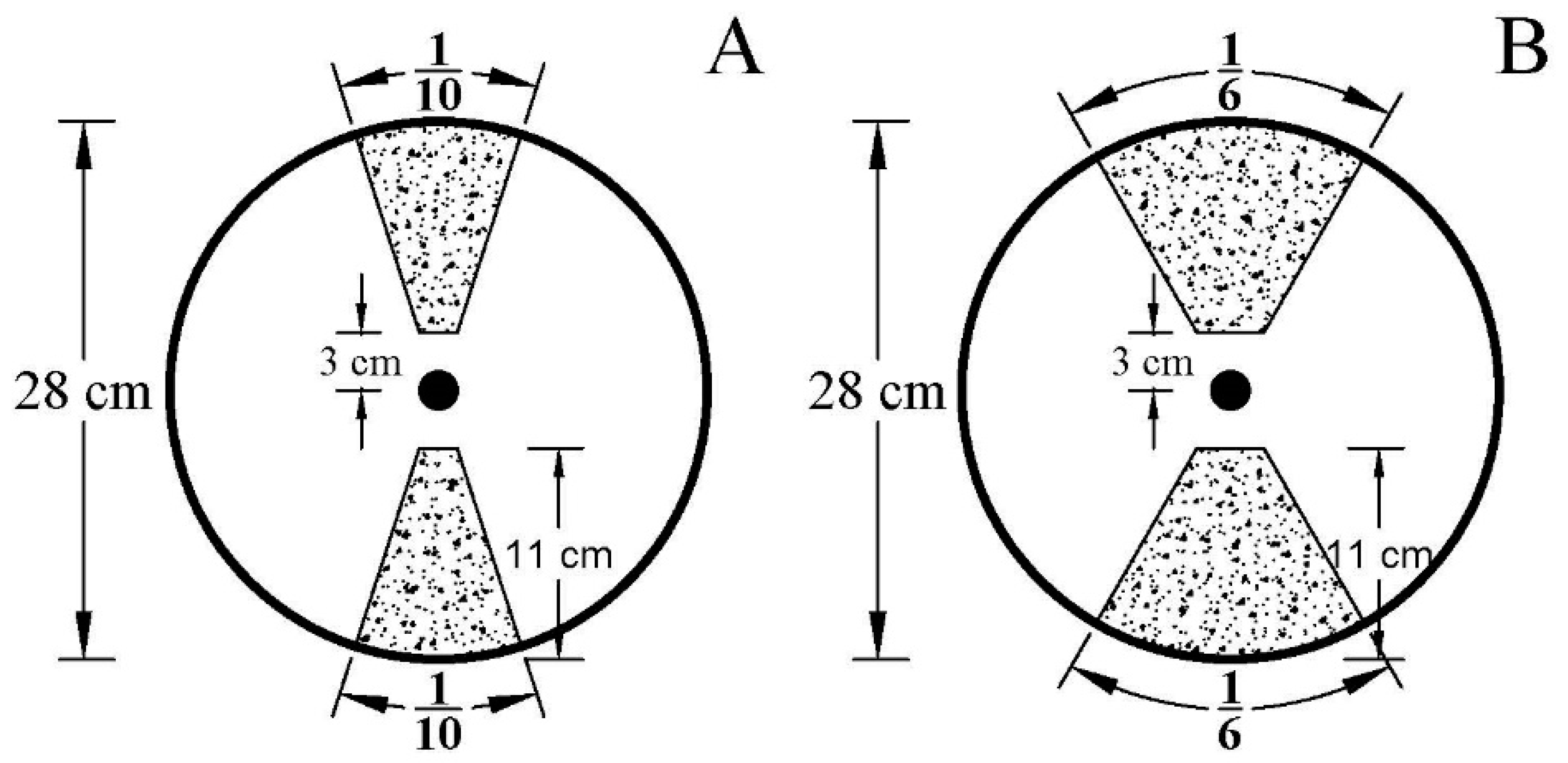
4.1. Plant Materials and Experimental Design
4.1.1. Experiment 1
4.1.2. Experiment 2
4.2. Measurements
4.2.1. Root Hydraulic Conductivity and Leaf Water Potential Measurements
4.2.2. Leaf Gas Exchange Parameters
4.2.3. Root Sampling and Measurements
4.2.4. Water Use Efficiency (WUE) and Harvest Index (HI)
4.3. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yu, L.; Gao, X.; Zhao, X. Global synthesis of the impact of droughts on crops’ water-use efficiency (WUE): Towards both high WUE and productivity. Agric. Syst. 2020, 177, 102723. [Google Scholar] [CrossRef]
- Vörösmarty, C.J.; Green, P.; Salisbury, J.; Lammers, R.B. Global water resources: Vulnerability from climate change and population growth. Science 2000, 289, 284–288. [Google Scholar] [CrossRef]
- Chen, Y.L.; Palta, J.; Prasad, P.V.V.; Siddique, K.H.M. Phenotypic variability in bread wheat root systems at the early vegetative stage. BMC Plan Biol. 2020, 20, 185. [Google Scholar] [CrossRef]
- Fang, Y.; Liang, L.Y.; Liu, S.; Xu, B.C.; Siddique, K.H.M.; Palta, J.A.; Chen, Y.L. Wheat cultivars with small root length density in the topsoil increased post-anthesis water use and grain yield in the semi-arid region on the Loess Plateau. Eur. J. Agron. 2021, 126, 126243. [Google Scholar] [CrossRef]
- Tajima, R. Importance of individual root traits to understand crop root system in agronomic and environmental contexts. Breed. Sci. 2021, 71, 13–19. [Google Scholar] [CrossRef]
- Palta, J.A.; Chen, X.; Milroy, S.P.; Rebetzke, G.J.; Watt, M. Large root systems: Are they useful in adapting wheat to dry environments? Funct. Plant Biol. 2011, 38, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Palta, J.A.; Watt, M. Vigorous crop root systems: Form and function for improving the capture of water and nutrients. In Crop Physiology; Academic Press: Cambridge, MA, USA, 2009; pp. 309–325. [Google Scholar] [CrossRef]
- Lilley, J.M.; Kirkegaard, J.A. Benefits of increased soil exploration by wheat roots. Field Crop. Res. 2011, 122, 118–130. [Google Scholar] [CrossRef]
- Liao, M.; Palta, J.A.; Fillery, I.R.P. Root characteristics of vigorous wheat improve early nitrogen uptake. Aust. J. Agric. Res. 2006, 57, 1097. [Google Scholar] [CrossRef]
- Palta, J.A.; Yang, J. Crop root system behaviour and yield. Field Crop. Res. 2014, 165, 1–4. [Google Scholar] [CrossRef]
- Ehdaie, B.; Layne, A.P.; Waines, J.G. Root system plasticity to drought influences grain yield in bread wheat. Euphytica 2011, 86, 219–232. [Google Scholar] [CrossRef]
- Song, L.; Li, F.M.; Fan, X.W.; Xiong, Y.C.; Wang, W.Q.; Wu, X.B.; Turner, N.C. Soil water availability and plant competition affect the yield of spring wheat. Eur. J. Agron. 2009, 31, 51–60. [Google Scholar] [CrossRef]
- Aziz, M.M.; Palta, J.A.; Siddique, K.H.M.; Sadras, V.O. Five decades of selection for yield reduced root length density and increased nitrogen uptake per unit root length in Australian wheat varieties. Plant Soil 2016, 413, 181–192. [Google Scholar] [CrossRef]
- Passioura, J.B. Roots and drought resistance. Agr. Water Manag. 1983, 7, 265–280. [Google Scholar] [CrossRef]
- Zhu, L.; Zhang, D.Y. Donald’s Ideotype and Growth Redundancy: A pot experimental test using an old and a modern spring wheat cultivar. PLoS ONE 2013, 8, e70006. [Google Scholar] [CrossRef]
- Zhang, S.Q.; Deng, X.P.; Shan, L. Changes of water use efficiency in wheat evolution and the relations with root growth. Chin. Sci. Bull. 2002, 47, 1879–1883. [Google Scholar] [CrossRef]
- Bertholdsson, N.O.; Brantestam, A.K. A century of Nordic barley breeding effects on early vigour root and shoot growth, straw length, harvest index and grain weight. Eur. J. Agron. 2009, 30, 266–274. [Google Scholar] [CrossRef]
- Hu, C.L.; Sadras, V.O.; Lu, G.Y.; Zhang, R.Z.; Yang, X.Y.; Zhang, S.L. Root pruning enhances wheat yield, harvest index and water-use efficiency in semiarid area. Field Crop. Res. 2019, 230, 62–71. [Google Scholar] [CrossRef]
- Fang, Y.; Du, Y.L.; Wang, J.; Wu, A.J.; Qiao, S.; Xu, B.C.; Zhang, S.Q.; Siddique, K.H.M.; Chen, Y.L. Moderate drought stress affected root growth and grain yield in old, modern and newly released cultivars of winter wheat. Front. Plant Sci. 2017, 8, 672–682. [Google Scholar] [CrossRef]
- Feddes, R.A.; Raats, P.A.C. Parameterizing the soil-water-plant root system. In Unsaturated Zone Modeling Progress, Challenges and Applications; Kluwer Academic Publishers: Berlin/Heidelberg, Germany, 2004; pp. 94–141. [Google Scholar]
- Palta, J.A.; Turner, N.C. Crop root system traits cannot be seen as a silver bullet delivering drought resistance. Plant Soil 2018, 439, 31–43. [Google Scholar] [CrossRef]
- Knipfer, T.; Fricke, W. Water uptake by seminal and adventitious roots in relation to whole-plant water flow in barley (Hordeum vulgare L.). J. Exp. Bot. 2011, 62, 717–733. [Google Scholar] [CrossRef]
- Sanchez-Romera, B.; Calvo-Polanco, M.N.; Ruiz-Lozano, J.M.; Zamarreno, M.A.; Arbona, V.; Garcıa-Mina, J.M.; Gomez-Cadenas, A.; Aroca, R. Involvement of the def-1 mutation in the response of tomato plants to arbuscular mycorrhizal symbiosis under well-watered and drought conditions. Plant Cell Physiol. 2018, 59, 248–261. [Google Scholar] [CrossRef] [PubMed]
- Lopez, F.B.; Nobel, P.S. Root hydraulic conductivity of two cactus species in relation to root age, temperature, and soil water status. J. Exp. Bot. 1991, 42, 143–149. [Google Scholar] [CrossRef]
- Zhao, C.X.; Deng, X.P.; Shan, L. Changes in root hydraulic conductivity during wheat evolution. J. Integr. Plant Biol. 2005, 47, 302–310. [Google Scholar] [CrossRef]
- Chai, Y.L. Effects of Drought Stress and Rehydration on Water Conductivity of Sorghum. Master’s Thesis, Northwest A&F University, Xianyang, China, 2011. [Google Scholar]
- Gorska, A.; Ye, Q.; Holbrook, N.M.; Zwieniecki, M. A Nitrate control of root hydraulic properties in plants: Translating local information to whole plant response. Plant Physiol. 2008, 148, 1159–1167. [Google Scholar] [CrossRef] [PubMed]
- Vysotskaya, L.B.; Arkhipova, T.N.; Timergalina, L.N.; Veselov, S.Y.; Dedov, A.V.; Kudoyarova, G.R. Effect of partial root excision on transpiration, root hydraulic conductance and leaf growth in wheat seedlings. J. Plant Physiol. 2004, 160, 1011–1015. [Google Scholar] [CrossRef]
- Wang, W.F.; Yang, X.Q.; Zhang, S.Q.; Shan, L. Effects of root excision and water stress on root hydraulics and TaPIPs expression in wheat seedlings. Acta Agron. Sin. 2013, 39, 1462–1468. [Google Scholar] [CrossRef]
- Wei, H.; Nie, X.Q.; Fan, M.S. Effects of root excision on the properties of root biology and water metabolism of maize seedlings. Chin. Agric. Sci. Bull. 2009, 25, 115–118. [Google Scholar]
- Vysotskaya, L.B.; Timergalina, L.N.; Simonyan, M.V.; Veselov, S.Y.; Kudoyarova, G.R. Growth rate, IAA and cytokinin content of wheat seedling after root pruning. Plant Growth Regul. 2001, 33, 51–57. [Google Scholar] [CrossRef]
- Wang, X.L.; Zhang, S.Q.; Shan, L. Effects of cultivars intercropping on maize water balance under different planting densities. Chin. J. Eco-Agri. 2013, 21, 171–178. [Google Scholar] [CrossRef]
- Luo, Z.; Kong, X.; Zhang, Y.; Li, W.; Zhang, D.; Dai, J.; Fang, S.; Chu, J.; Dong, H. Leaf-derived jasmonate mediates water uptake from hydrated cotton roots under partial root-zone irrigation. Plant Physiol. 2019, 180, 1660–1676. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Li, S.; He, X.; Jiang, W.; Zhang, D.; Liu, B.; Li, Q. CO2 enrichment enhanced drought resistance by regulating growth, hydraulic conductivity and phytohormone contents in the root of cucumber seedlings. Plant Physiol. Biochem. 2020, 152, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Hirasawa, T. Leaf photosynthesis of upland and lowland crops grown under moisture-rich conditions. In The Leaf: A Platform for Performing Photosynthesis; Adams, W., III, Terashima, I., Eds.; Springer: Berlin/Heidelberg, Germany, 2018; Volume 44. [Google Scholar] [CrossRef]
- Liu, X.F.; Zhang, S.Q.; Shan, L. Heterosis for water uptake by maize (Zea mays L.) roots under water deficit: Responses at cellular, single-root and whole-root system levels. J. Arid. Land. 2013, 5, 255–265. [Google Scholar] [CrossRef]
- Rodriguez-Dominguez, C.M.; Forner, A.; Martorell, S.; Choat, B.; Lopez, R.; Peters, J.M.; Sack, L. Leaf water potential measurements using the pressure chamber: Synthetic testing of assumptions towards best practices for precision and accuracy. Plant Cell Environ. 2022, 45, 2037–2061. [Google Scholar] [CrossRef] [PubMed]
- Steudle, E. Water uptake by plant roots: An integration of views. Plant Soil. 2000, 226, 45–56. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, X.; Wang, N.; Chen, Y.; Zhang, S. Changes in the yield and associated photosynthetic traits of dry-land winter wheat (Triticum aestivum L.) from the 1940s to the 2010s in Shaanxi Province of China. Field Crop. Res. 2014, 167, 1–10. [Google Scholar] [CrossRef]
- Tracy, S.R.; Nagel, K.A.; Postma, J.A.; Fassbender, H.; Wasson, A.; Watt, M. Crop Improvement from phenotyping roots: Highlights reveal expanding opportunities. Trends Plant Sci. 2020, 25, 105–118. [Google Scholar] [CrossRef]
- Fang, Y.; Xu, B.C.; Turner, N.; Li, F.M. Grain yield, dry matter accumulation and remobilization, and root respiration in winter wheat as affected by seeding rate and root pruning. Eur. J. Agron. 2010, 33, 257–266. [Google Scholar] [CrossRef]
- Ma, S.C.; Li, F.M.; Xu, B.C.; Huang, Z.B. Effect of lowering the root/shoot ratio by pruning roots on water use efficiency and grain yield of winter wheat. Field Crop. Res. 2010, 115, 158–164. [Google Scholar] [CrossRef]
- Song, X.; Zhou, G.; He, Q. Critical leaf water content for maize photosynthesis under drought stress and its response to rewatering. Sustainability 2021, 13, 7218. [Google Scholar] [CrossRef]
- Shao, R.; Jia, S.; Tang, Y.; Zhang, J.; Li, H.; Li, L.; Chen, J.; Guo, J.; Wang, H.; Yang, Q.; et al. Soil water deficit suppresses development of maize ear by altering metabolism and photosynthesis. Environ. Exp. Bot. 2021, 192, 104651. [Google Scholar] [CrossRef]
- Puangbut, D.; Jogloy, S.; Vorasoot, N.; Songsri, P. Photosynthetic and physiological responses to drought of Jerusalem artichoke genotypes differing in drought resistance. Agr. Water Manag. 2022, 259, 107252. [Google Scholar] [CrossRef]
- Simonin, K.A.; Burns, E.; Choat, B.; Barbour, M.M.; Dawson, T.E.; Franks, P.J. Increasing leaf hydraulic conductance with transpiration rate minimizes the water potential drawdown from stem to leaf. J. Exp. Bot. 2015, 66, 1303–1315. [Google Scholar] [CrossRef] [PubMed]
- Kodama, A.; Watanabe, T.; Yamaguchi, M.; Narita, R.; Katsuhara, M.; Sato, K.; Ookawa, T.; Hirasawa, T. Accession difference in leaf photosynthesis, root hydraulic conductance and gene expression of root aquaporins under salt stress in barley seedlings. Plant Prod. Sci. 2020, 24, 73–82. [Google Scholar] [CrossRef]
- Lou, C.H. Signal transport and integral behavior in maintaining water economy in higher plant. Chin. Bull Bot. 2000, 17, 475–477. [Google Scholar]
- Siddique, K.; Belford, R.K.; Tennant, D. Root:shoot ratios of old and modern, tall and semi-dwarf wheats in a mediterranean environment. Plant Soil 1990, 121, 89–98. [Google Scholar] [CrossRef]
- Figueroa-Bustos, V.; Palta, J.; Chen, Y.; Siddique, K. Characterization of root and shoot traits in wheat cultivars with putative differences in root system size. Agronomy 2018, 8, 109. [Google Scholar] [CrossRef]
- Chai, S.W.; Liu, W.Z.; Li, Y.Y. Effect of root cutting on leaf photosynthesis rate and water use efficiency of maize. Chin. J. App. Ecol. 2002, 12, 1716–1718. [Google Scholar] [CrossRef]
- Hall, A.J.; Richards, R.A. Prognosis for genetic improvement of yield potential and water-limited yield of major grain crops. Field Crop. Res. 2013, 143, 18–33. [Google Scholar] [CrossRef]
- Liu, H.S.; Li, F.M.; Hao, X.U. Carbon consumption of roots and its relationship to yield formation in spring wheat as affected by soil moisture. Chin. J. Plant Ecol. 2004, 28, 191–197. [Google Scholar] [CrossRef]
- Chimungu, J.G.; Brown, K.M.; Lynch, J.P. Reduced root cortical cell file number improves drought tolerance in maize. Plant Physiol. 2014, 166, 1943–1955. [Google Scholar] [CrossRef] [Green Version]
- Sreeman, S.M.; Vijayaraghavareddy, P.; Sreevathsa, R.; Rajendrareddy, S.; Arakesh, S.; Bharti, P.; Dharmappa, P.; Soolanayakanahally, R. Introgression of Physiological Traits for a comprehensive improvement of drought adaptation in crop plants. Front. Chem. 2018, 6, 92. [Google Scholar] [CrossRef] [PubMed]
- Corales, M.; Nguyen, N.T.A.; Abiko, T.; Mochizuki, T. Mapping quantitative trait loci for water uptake of rice under aerobic conditions. Plant Prod. Sci. 2020, 23, 436–451. [Google Scholar] [CrossRef]
- Xiu, W.W.; Tian, X.D.; Chen, C.X. Comparative study on the characteristics of maize root under the conditions of saturated irrigation in different eras. J. Maize Sci. 2013, 21, 78–87. [Google Scholar]
- Liu, M.; Wu, G.J.; Lu, D.X. Improvement of nitrogen use efficiency and the relationship with root system characters of maize cultivars in different year. Plant Nutr. Fert. Sci. 2017, 23, 71–82. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Items | Pot Experiment 1 | Pot Experiment 2 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| V6 | V12 | V6 | V12 | ||||||||
| QL | XY | QL | XY | R0 | RP1 | RP2 | R0 | RP1 | RP2 | ||
| WW | Root (g) | 24.53 ± 1.59 a | 20.01 ± 1.44 b | 39.42 ± 2.47 a | 30.13 ± 3.48 b | 26.38 ± 1.51 a | 21.51 ± 1.14 b | 18.86 ± 1.28 c | 41.06 ± 1.91 a | 38.62 ± 1.77 ab | 36.53 ± 0.71 b |
| Shoot (g) | 64.14 ± 4.11 a | 54.15 ± 1.84 bc | 129.61 ± 7.91 a | 119.20 ± 9.01 a | 58.58 ± 1.57 a | 56.40 ± 3.05 a | 56.05 ± 2.05 a | 127.74 ± 15.38 a | 131.42 ± 10.37 a | 125.02 ± 3.48 a | |
| R/S | 0.38 ± 0.03 b | 0.34 ± 0.01 c | 0.31 ± 0.03 c | 0.26 ± 0.05 d | 0.45 ± 0.01 a | 0.38 ± 0.02 b | 0.33 ± 0.03 c | 0.32 ± 0.03 bc | 0.29 ± 0.02 c | 0.29 ± 0.01 c | |
| WS | Root (g) | 22.98 ± 1.23 a | 18.91 ± 1.56 b | 29.29 ± 1.40 ab | 23.58 ± 1.65 b | 24.89 ± 0.75 a | 20.37 ± 0.40 b | 17.51 ± 0.71 c | 28.62 ± 2.03 c | 26.22 ± 0.67 cd | 24.15 ± 0.93 d |
| Shoot (g) | 59.36 ± 2.87 ab | 52.16 ± 1.87 c | 70.84 ± 7.48 b | 67.12 ± 2.32 b | 53.81 ± 1.17 b | 52.19 ± 3.17 b | 51.73 ± 1.80 b | 78.75 ± 2.70 b | 76.03 ± 13.36 b | 74.53 ± 8.17 b | |
| R/S | 0.43 ± 0.02 a | 0.36 ± 0.01 b | 0.42 ± 0.06 a | 0.35 ± 0.03 b | 0.47 ± 0.02 a | 0.39 ± 0.02 b | 0.34 ± 0.02 c | 0.39 ± 0.02 a | 0.35 ± 0.05 b | 0.30 ± 0.01 bc | |
| Treatment | Grain Yield (g/pot) | 100-Kernel Weight (g) | Ear Length (cm) | HI |
|---|---|---|---|---|
| WW | ||||
| QL | 131.56b ± 7.31 | 29.18 ± 0.84 ab | 15.62 ± 1.02 b | 0.54 ± 0.03 b |
| XY | 154.65 ± 5.88 a | 31.83 ± 1.31 a | 17.45 ± 0.38 a | 0.57 ± 0.05 a |
| R0 | 148.01 ± 4.67 b | 36.51 ± 0.92 b | 15.58 ± 1.79 a | 0.53 ± 0.01 b |
| RP1 | 167.06 ± 2.47 a | 38.83 ± 0.89 a | 16.62 ± 0.99 a | 0.57 ± 0.05 a |
| RP2 | 143.88 ± 8.73 b | 36.75 ± 0.84 b | 15.07 ± 1.74 a | 0.55 ± 0.02 b |
| WS | ||||
| QL | 74.43 ± 2.40 d | 25.83 ± 0.90 d | 12.81 ± 0.88 d | 0.50 ± 0.02 c |
| XY | 88.03 ± 3.90 c | 28.78 ± 1.86 c | 14.05 ± 1.07 c | 0.53 ± 0.02 b |
| R0 | 85.90 ± 1.15 c | 29.12 ± 0.64 d | 9.50 ± 1.12 b | 0.47 ± 0.01 c |
| RP1 | 89.52 ± 2.78 c | 31.83 ± 0.41 c | 10.66 ± 1.83 b | 0.48 ± 0.02 c |
| RP2 | 83.77 ± 2.38 c | 32.68 ± 2.19 c | 10.87 ± 0.95 b | 0.46 ± 0.03 c |
| Probability level of ANOVA | ||||
| W | ** | ** | * | ** |
| R | ** | * | NS | ** |
| W × R | * | NS | NS | NS |
| HKW | EL | WUE | ET | Lpr | Ψleaf | Pn | GS | E | iWUE | RW | SW | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| GY | 0.796 ** | 0.692 ** | 0.731 * | 0.903 ** | 0.701 ** | 0.892 ** | 0.885 ** | 0.857 * | 0.786 * | −0.301 * | 0.126 | 0.912 ** |
| HKW | 0.319 * | 0.321 * | 0.623 ** | 0.431 ** | 0.638 ** | 0.625 ** | 0.724 | 0.463 | 0.153 | −0.126 | 0.588 ** | |
| EL | −0.338 | 0.743 ** | 0.211 | 0.414 | 0.760 ** | 0.640 * | 0.472 | −0.327 * | 0.332 | 0.530 | ||
| WUE | −0.398 ** | 0.692 * | 0.242 | −0.206 | −0.551 | −0.474 ** | 0.655 ** | −0.493 * | −0.285 * | |||
| ET | 0.689 ** | 0.614 * | 0.584 | 0.839 * | 0.932 ** | −0.575 ** | 0.348 * | 0.838 ** | ||||
| Lpr | 0.851 ** | 0.642 | 0.570 | 0.662 ** | −0.150 | −0.218 | 0.745 ** | |||||
| Ψleaf | 0.563 | −0.861 * | −0.847 ** | 0.357 * | −0.063 | 0.198 | ||||||
| Pn | 0.864 ** | 0.835 ** | −0.263 | 0.068 | 0.835 ** | |||||||
| GS | 0.728 * | −0.264 | 0.028 | 0.769 ** | ||||||||
| E | −0.702 ** | 0.419 ** | 0.778 ** | |||||||||
| iWUE | −0.741 ** | −0.434 |
| Texture | PH | Bulk Density (g/cm3) | Available N (mg/kg) | Available p (mg/kg) | Available K (mg/kg) | Organic Matter (g/kg) | Total N (mg/kg) |
|---|---|---|---|---|---|---|---|
| Loam | 7.6 | 1.37 | 14.22 | 13.8 | 124.6 | 0.92 | 18.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, M.; Zhang, L.; Ren, Y.; Zhang, T.; Zhang, S.; Li, H.; Chen, Y.; Zhang, S. The Higher Water Absorption Capacity of Small Root System Improved the Yield and Water Use Efficiency of Maize. Plants 2022, 11, 2300. https://doi.org/10.3390/plants11172300
Yan M, Zhang L, Ren Y, Zhang T, Zhang S, Li H, Chen Y, Zhang S. The Higher Water Absorption Capacity of Small Root System Improved the Yield and Water Use Efficiency of Maize. Plants. 2022; 11(17):2300. https://doi.org/10.3390/plants11172300
Chicago/Turabian StyleYan, Minfei, Li Zhang, Yuanyuan Ren, Tingting Zhang, Shaowei Zhang, Hongbing Li, Yinglong Chen, and Suiqi Zhang. 2022. "The Higher Water Absorption Capacity of Small Root System Improved the Yield and Water Use Efficiency of Maize" Plants 11, no. 17: 2300. https://doi.org/10.3390/plants11172300