

Crop Rotation with Marigold Promotes Soil Bacterial Structure to Assist in Mitigating Clubroot Incidence in Chinese Cabbage

 ,
,
Abstract
:1. Introduction
2. Results
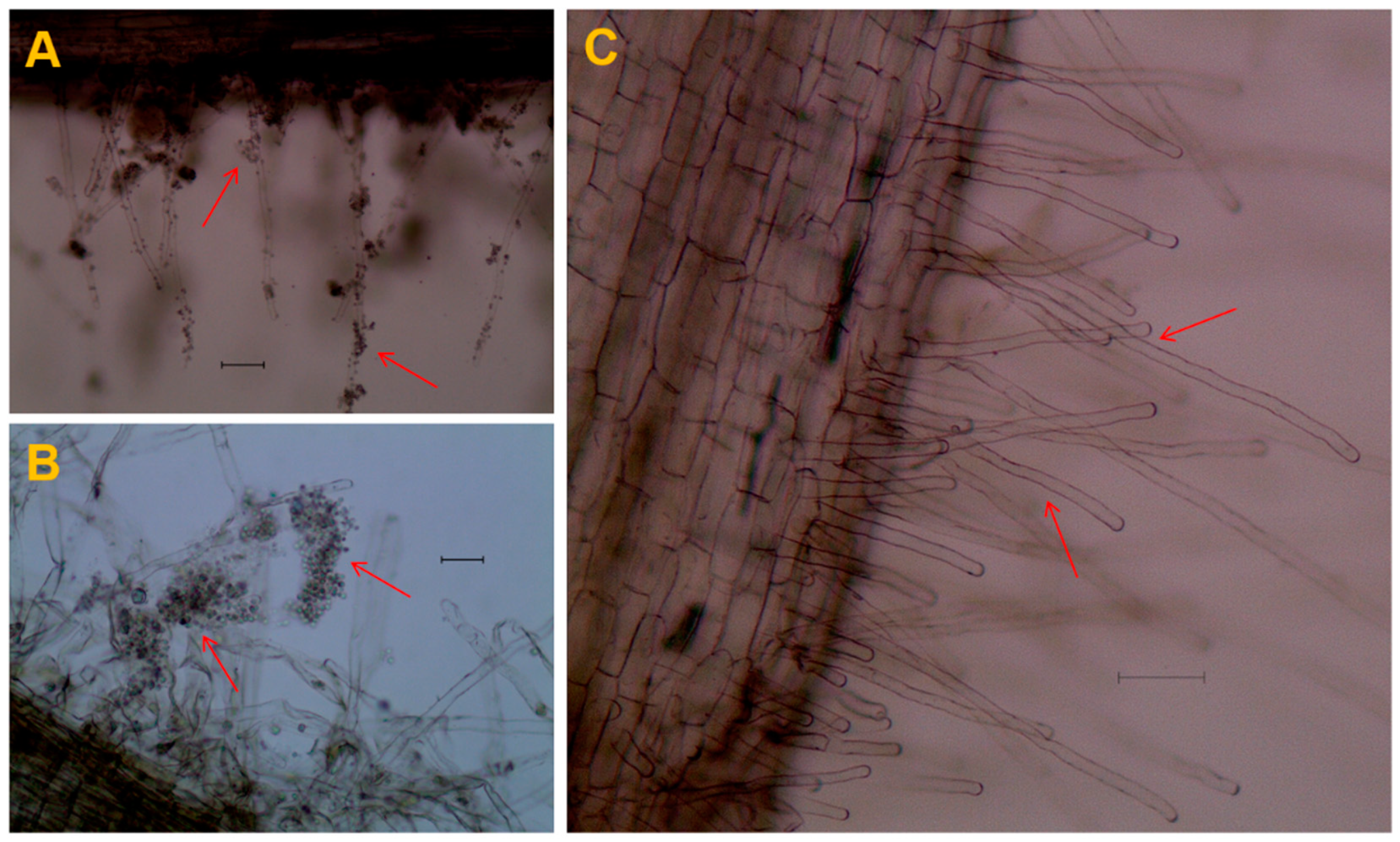
2.1. Marigold Roots Enhance the Germination of P. brassicae Resting Spores
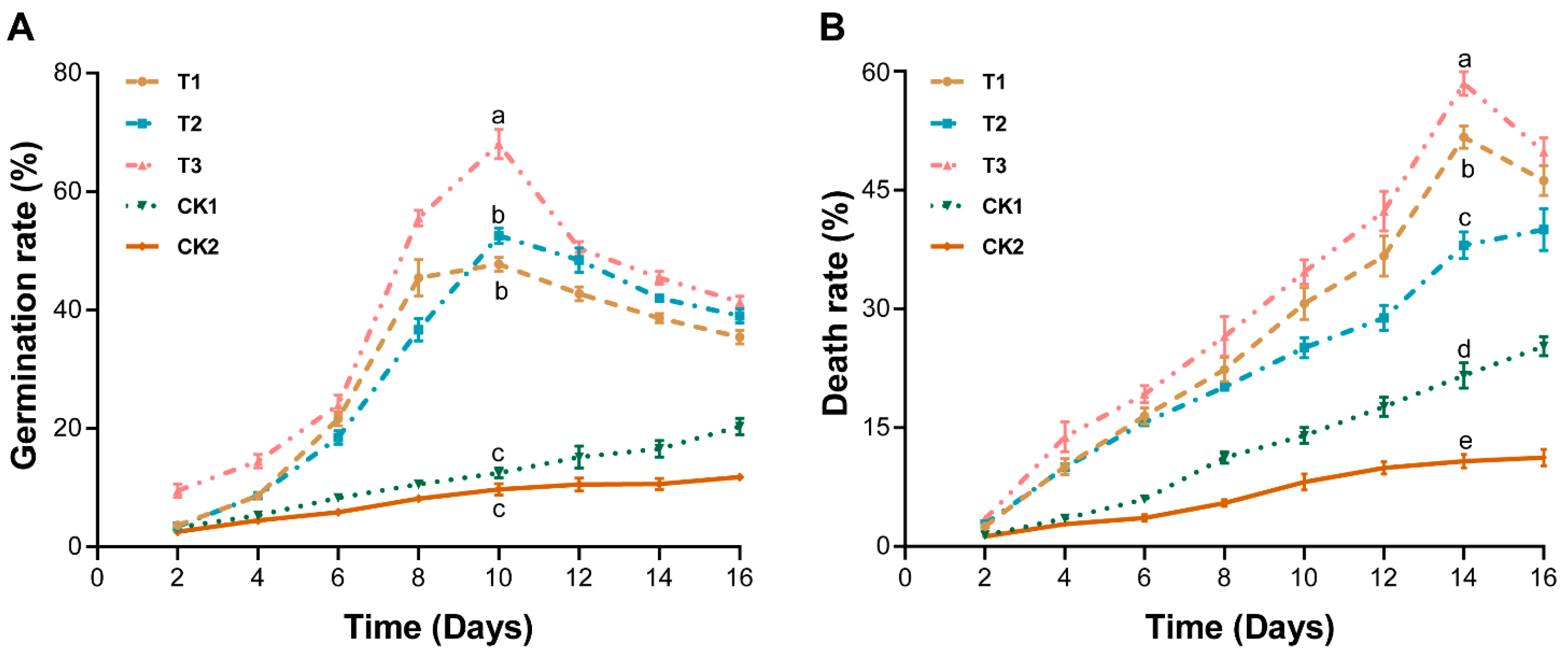
2.2. Marigold Root Exudates, Crude Extract, and Powder Influence the Germination and Death of P. brassicae Resting Spores
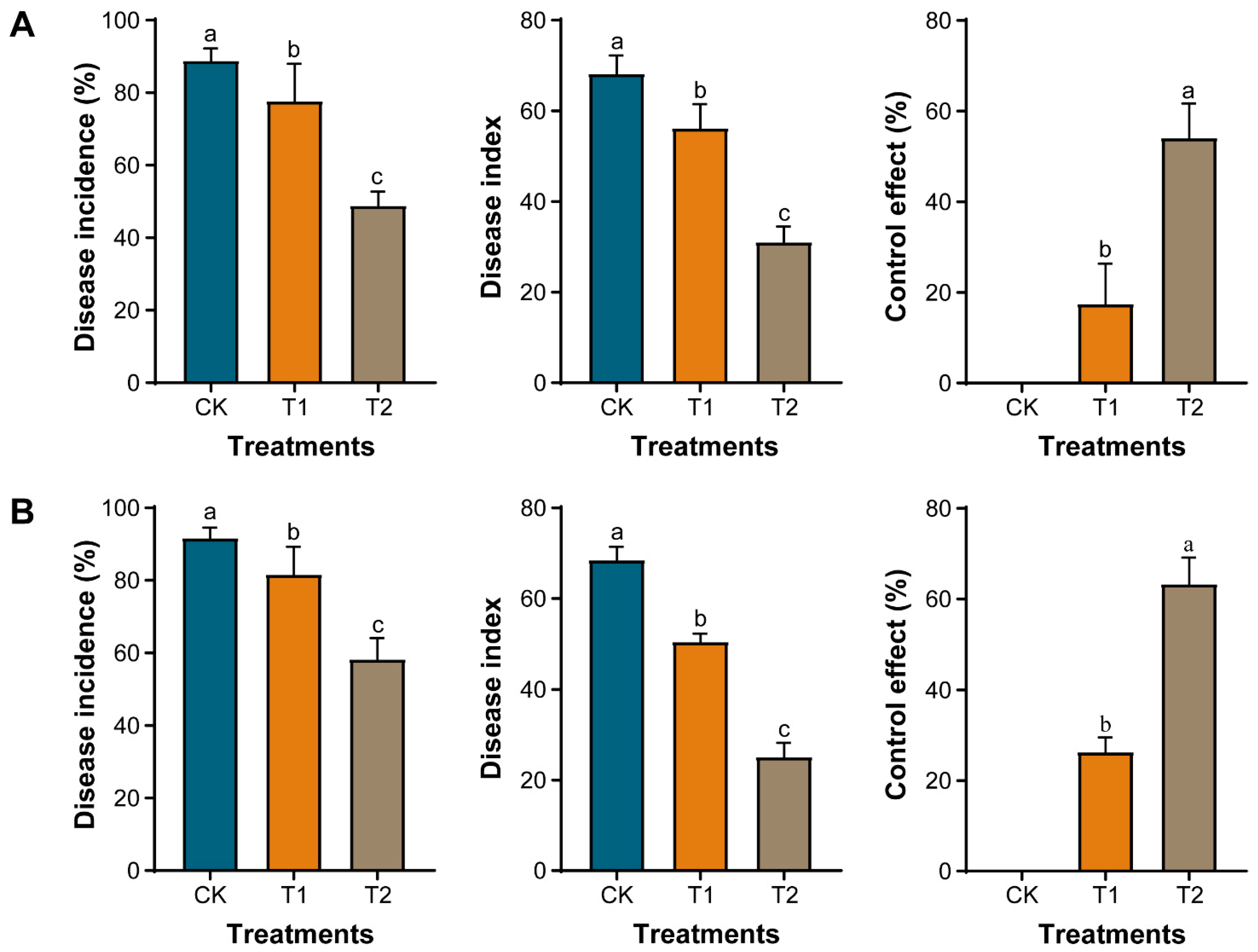
2.3. Effect of Marigold Powder on the Incidence of Clubroot in Chinese Cabbage
2.4. Marigold-Chinese Cabbage Crop Rotation Suppresses the Incidence of Clubroot in Chinese Cabbage under Greenhouse and Field Conditions
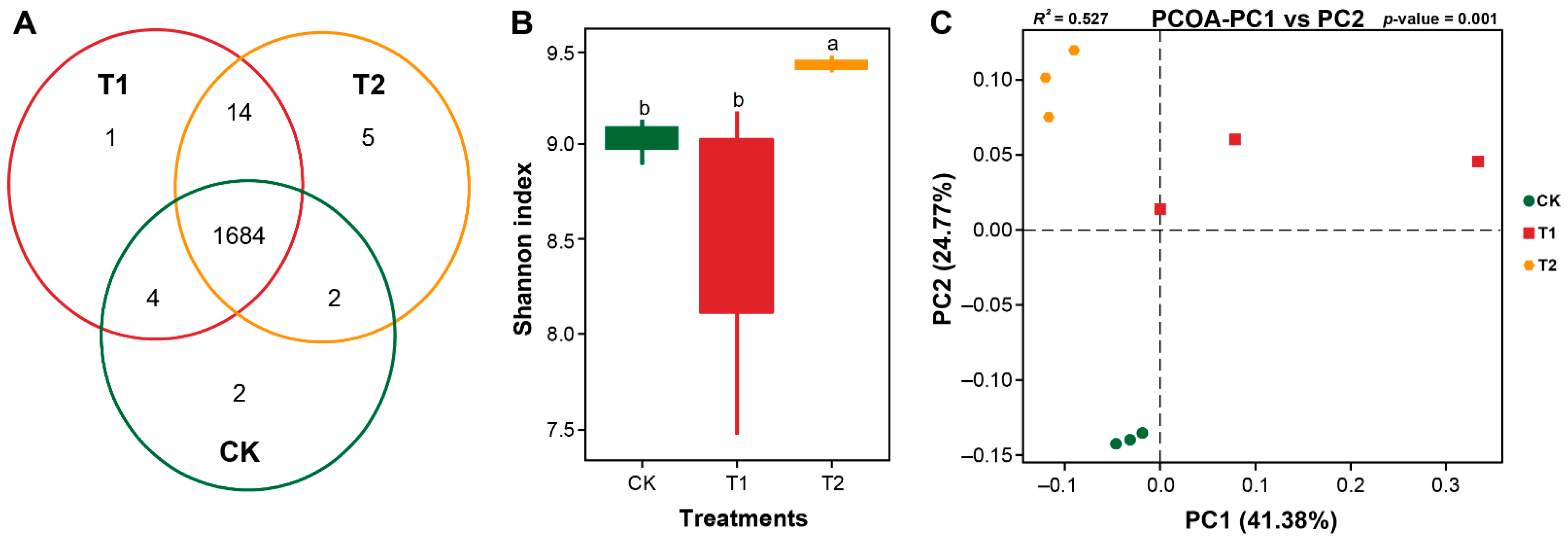
2.5. Marigold-Chinese Cabbage Crop Rotation Affects the Assembly, Diversity, and Structure of Rhizosphere Bacterial Communities
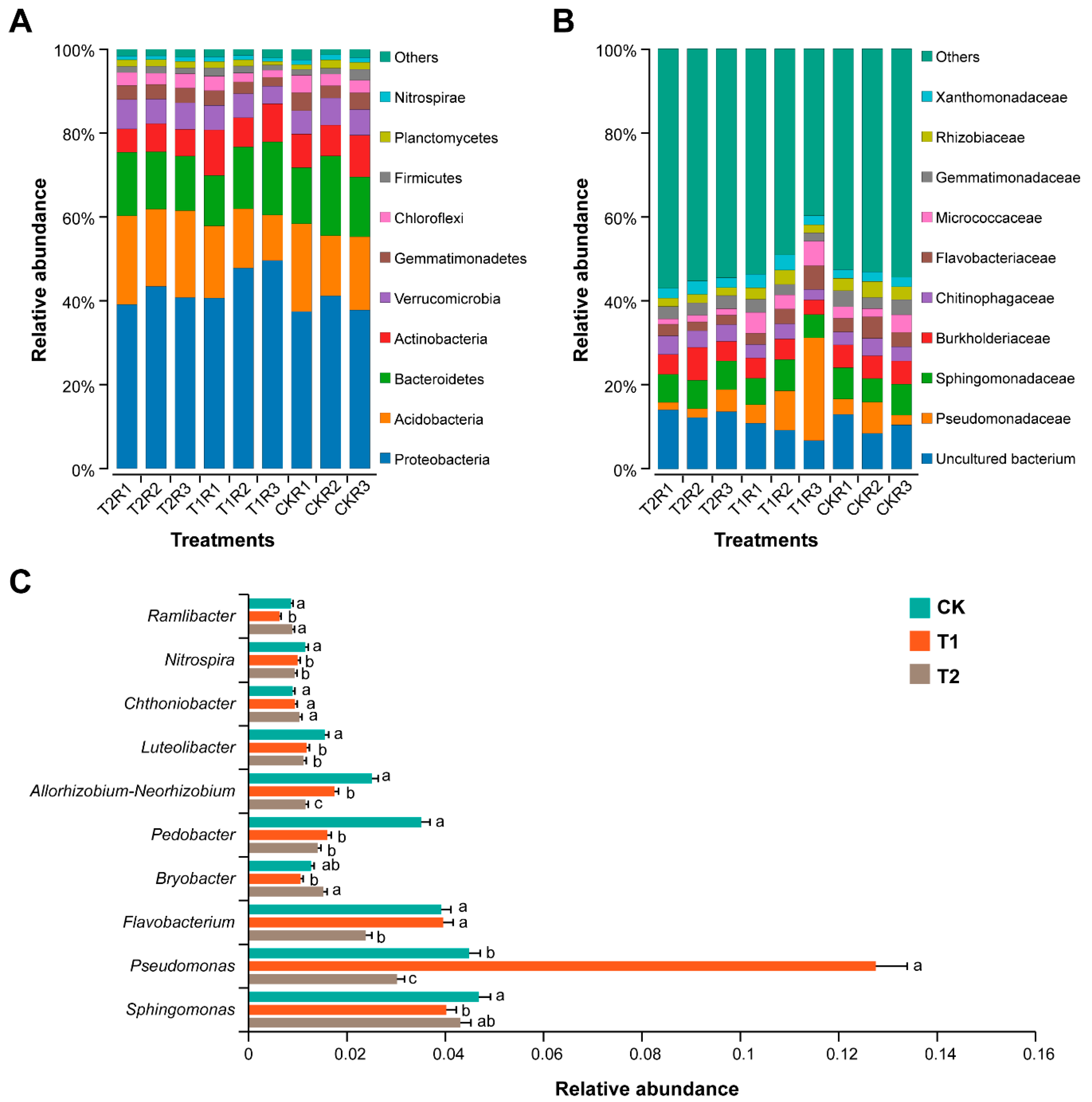
2.6. Impact of Marigold-Chinese Cabbage Crop Rotation on Rhizosphere Bacterial Community Composition
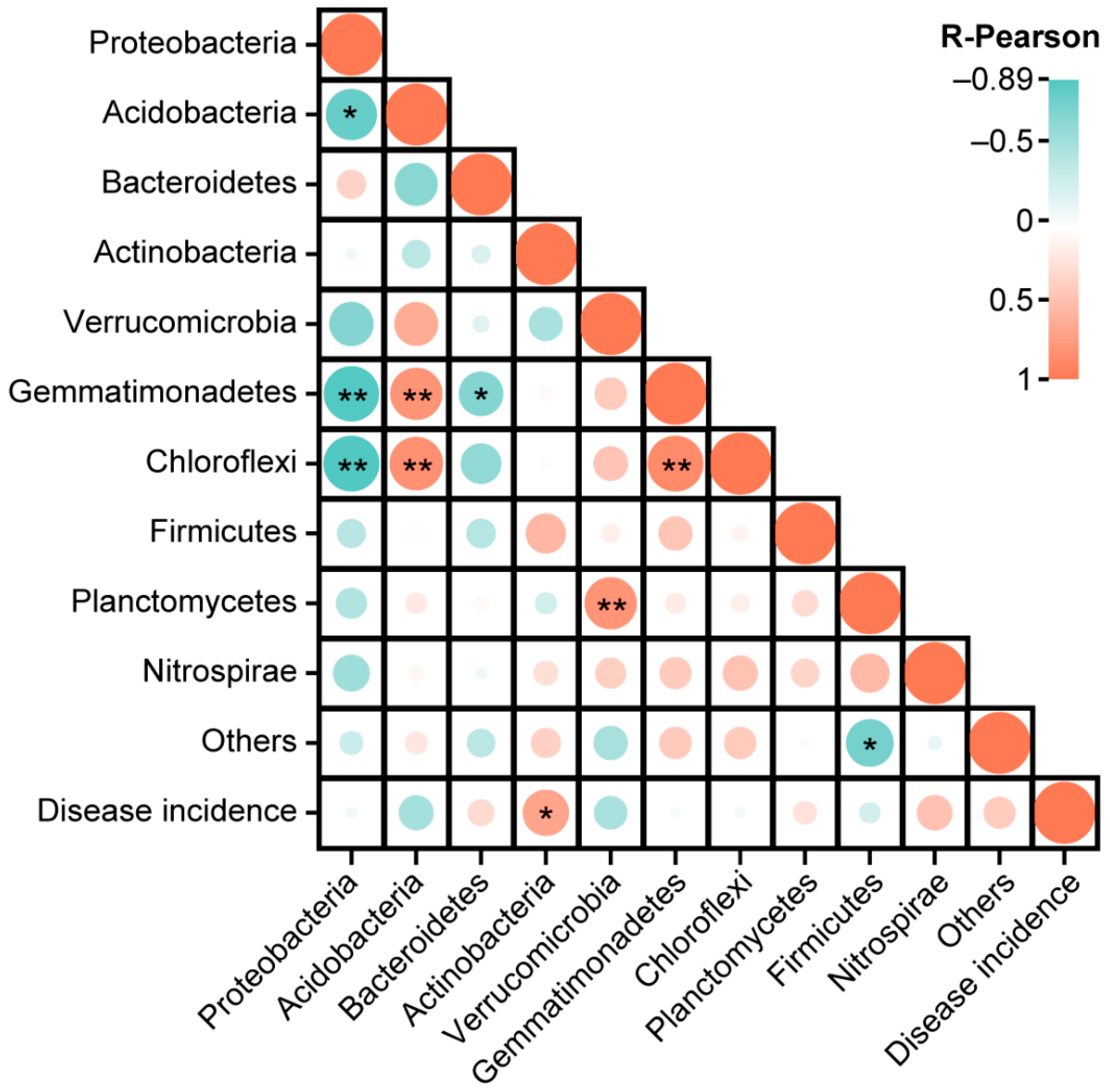
2.7. Correlation Analysis
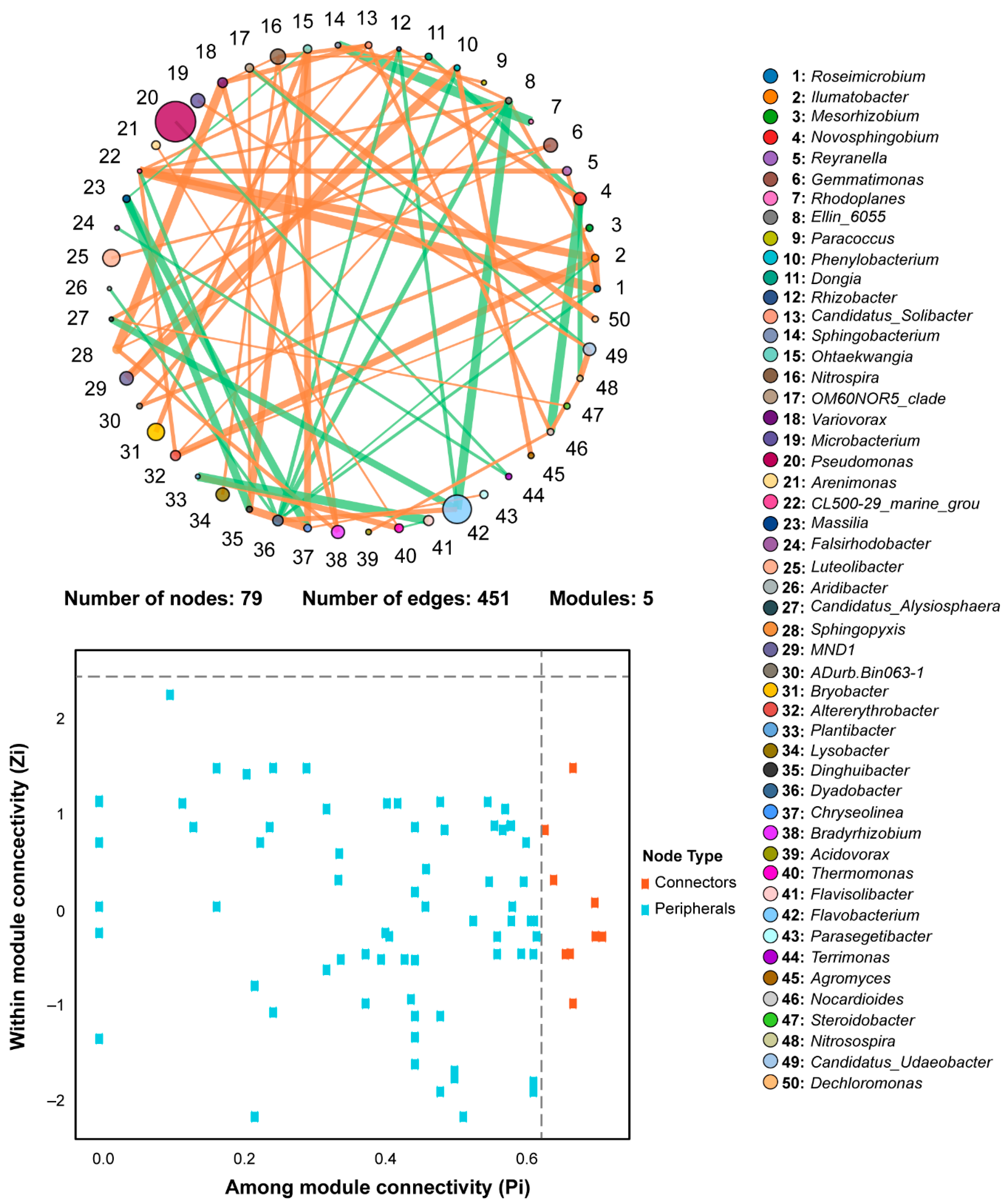
2.8. Co-Occurrence Network Analysis of Rhizosphere Bacterial Communities
3. Discussion
4. Materials and Methods
4.1. Experimental Site and Plant Material
4.2. Preparation of Plasmodiophora Brassicae Spore Suspension
4.3. Assessment of the Effect of Marigold on the Germination of P. brassicae Resting Spores
4.4. Marigold Root Exudates, Crude Extract, and Powder Preparation
4.4.1. Preparation of Root Exudates
4.4.2. Preparation of Marigold Tissues Crude Extract
4.4.3. Preparation of Marigold Powder
4.5. Analysis of Marigold Root Exudates, Crude Extract, and Powder on the Germination and Death of P. brassicae Resting Spores
4.6. Greenhouse Experiments
4.6.1. Evaluation the Effect of Marigold Powder on Clubroot
4.6.2. Investigation of Disease Incidence, Disease Index, and Control Effect
4.6.3. Evaluation of the Effect of Marigold Crop Rotation on Clubroot Incidence in Chinese Cabbage
4.7. Field Experiment
4.8. Soil Samples Collection and DNA Extraction
4.9. High Throughput Amplicon Sequencing and Analysis of Rhizosphere Bacterial Diversity
4.10. Bioinformatics Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wei, L.; Yang, J.; Ahmed, W.; Xiong, X.; Liu, Q.; Huang, Q.; Ji, G. Unraveling the Association between Metabolic Changes in Inter-Genus and Intra-Genus Bacteria to Mitigate Clubroot Disease of Chinese Cabbage. Agronomy 2021, 11, 2424. [Google Scholar] [CrossRef]
- Dixon, G.R. The occurrence and economic impact of Plasmodiophora brassicae and clubroot disease. J. Plant Growth Regul. 2009, 28, 194–202. [Google Scholar] [CrossRef]
- Chai, A.; Xie, X.; Shi, Y.; Li, B. Research status of clubroot (Plasmodiophora brassicae) on cruciferous crops in China. Can. J. Plant Pathol. 2014, 36, 142–153. [Google Scholar] [CrossRef]
- Peng, G.; Pageau, D.; Strelkov, S.E.; Gossen, B.D.; Hwang, S.-F.; Lahlali, R. A> 2-year crop rotation reduces resting spores of Plasmodiophora brassicae in soil and the impact of clubroot on canola. Eur. J. Agron. 2015, 70, 78–84. [Google Scholar] [CrossRef]
- Lachhab, N.; Sanzani, S.M.; Fallanaj, F.; Youssef, K.; Nigro, F.; Boselli, M.; Ippolito, A. Protein hydrolysates as resistance inducers for controlling green mould of citrus fruit. Acta Hortic 2015, 1065, 1593–1598. [Google Scholar] [CrossRef]
- Youssef, K.; Roberto, S.R.; Tiepo, A.N.; Constantino, L.V.; de Resende, J.T.V.; Abo-Elyousr, K.A. Salt solution treatments trigger antioxidant defense response against gray mold disease in table grapes. J. Fungi 2020, 6, 179. [Google Scholar] [CrossRef] [PubMed]
- Hussien, A.; Ahmed, Y.; Al-Essawy, A.-H.; Youssef, K. Evaluation of different salt-amended electrolysed water to control postharvest moulds of citrus. Trop. Plant Pathol. 2018, 43, 10–20. [Google Scholar] [CrossRef]
- Murakami, H.; Tsushima, S.; Kuroyanagi, Y.; Shishido, Y. Reduction of resting spore density of Plasmodiophora brassicae and clubroot disease severity by liming. Soil Sci. Plant Nutr. 2002, 48, 685–691. [Google Scholar] [CrossRef]
- Ohshima, T.; Komyoji, T.; Mitani, S.; Matsuo, N.; Nakajima, T. Development of a novel fungicide, cyazofamid. J. Pestic. Sci. 2004, 29, 147–152. [Google Scholar] [CrossRef]
- Liu, Y.; Huang, X.; Ke, S.; LIU, H.-y. Evaluation of resistance of rapeseed varieties to club root infected by Plasmodiophora brassicae in Sichuan. Chin. J. Oil Crop Sci. 2009, 31, 90–93. [Google Scholar]
- Peng, G.; Lahlali, R.; Hwang, S.-F.; Pageau, D.; Hynes, R.K.; McDonald, M.R.; Gossen, B.D.; Strelkov, S.E. Crop rotation, cultivar resistance, and fungicides/biofungicides for managing clubroot (Plasmodiophora brassicae) on canola. Can. J. Plant Pathol. 2014, 36, 99–112. [Google Scholar] [CrossRef]
- Hiddink, G.A.; Termorshuizen, A.J.; van Bruggen, A.H. Mixed cropping and suppression of soilborne diseases. Genet. Eng. Biofertilisation Soil Qual. Org. Farming 2010, 4, 119–146. [Google Scholar]
- Zhang, W.; Long, X.; Huo, X.; Chen, Y.; Lou, K. 16S rRNA-based PCR-DGGE analysis of actinomycete communities in fields with continuous cotton cropping in Xinjiang, China. Microb. Ecol. 2013, 66, 385–393. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.-y.; Zhang, S.; Zhu, X.-z.; LU, L.-m.; Wang, C.-y.; Li, C.-h.; Cui, J.-j.; Zhou, Z.-g. Effects of soil salinity on rhizosphere soil microbes in transgenic Bt cotton fields. J. Integr. Agric. 2017, 16, 1624–1633. [Google Scholar] [CrossRef]
- Zhang, J.; Wei, L.; Yang, J.; Ahmed, W.; Wang, Y.; Fu, L.; Ji, G. Probiotic consortia: Reshaping the rhizospheric microbiome and its role in suppressing root-rot disease of Panax notoginseng. Front. Microbiol. 2020, 11, 701. [Google Scholar] [CrossRef]
- Cai, Q.; Zhou, G.; Ahmed, W.; Cao, Y.; Zhao, M.; Li, Z.; Zhao, Z. Study on the relationship between bacterial wilt and rhizospheric microbial diversity of flue-cured tobacco cultivars. Eur. J. Plant Pathol. 2021, 160, 265–276. [Google Scholar] [CrossRef]
- Newton, A.; Begg, G.; Swanston, J. Deployment of diversity for enhanced crop function. Ann. Appl. Biol. 2009, 154, 309–322. [Google Scholar] [CrossRef]
- Li, X.-G.; Wang, X.-X.; Dai, C.-C.; Zhang, T.-L.; Xie, X.-G.; Ding, C.-F.; Wang, H.-W. Effects of intercropping with Atractylodes lancea and application of bio-organic fertiliser on soil invertebrates, disease control and peanut productivity in continuous peanut cropping field in subtropical China. Agrofor. Syst. 2014, 88, 41–52. [Google Scholar] [CrossRef]
- Ahmed, W.; Yang, J.; Tan, Y.; Munir, S.; Liu, Q.; Zhang, J.; Ji, G.; Zhao, Z. Ralstonia solanacearum, a deadly pathogen: Revisiting the bacterial wilt biocontrol practices in tobacco and other Solanaceae. Rhizosphere 2022, 21, 100479. [Google Scholar] [CrossRef]
- Gómez-Rodrıguez, O.; Zavaleta-Mejıa, E.; Gonzalez-Hernandez, V.; Livera-Munoz, M.; Cárdenas-Soriano, E. Allelopathy and microclimatic modification of intercropping with marigold on tomato early blight disease development. Field Crops Res. 2003, 83, 27–34. [Google Scholar] [CrossRef]
- Li, Y.; Feng, J.; Zheng, L.; Huang, J.; Yang, Y.; Li, X. Intercropping with marigold promotes soil health and microbial structure to assist in mitigating tobacco bacterial wilt. J. Plant Pathol. 2020, 102, 731–742. [Google Scholar] [CrossRef]
- Hooks, C.R.; Wang, K.-H.; Ploeg, A.; McSorley, R. Using marigold (Tagetes spp.) as a cover crop to protect crops from plant-parasitic nematodes. Appl. Soil Ecol. 2010, 46, 307–320. [Google Scholar] [CrossRef]
- Bakshi, L.; Ghosh, R. Marigold Biopesticide as an Alternative to Conventional Chemical Pesticides. J. Adv. Sci. Res. 2022, 13, 26–33. [Google Scholar] [CrossRef]
- El-Hamawi, M.; Youssef, M.; Zawam, H.S. Management of Meloidogyne incognita, the root-knot nematode, on soybean as affected by marigold and sea ambrosia (damsisa) plants. J. Pest Sci. 2004, 77, 95–98. [Google Scholar] [CrossRef]
- Kumar, N.; Krishnappa, K.; Reddy, B.; Ravichandra, N.; Karuna, K. Intercropping for the management of root-knot nematode, Meloidogyne incognita in vegetable-based cropping systems. Indian J. Nematol. 2005, 35, 46–49. [Google Scholar]
- Hu, Y.; Zhao, W.; Li, X.; Feng, J.; Li, C.; Yang, X.; Guo, Q.; Wang, L.; Chen, S.; Li, Y. Integrated biocontrol of tobacco bacterial wilt by antagonistic bacteria and marigold. Sci. Rep. 2021, 11, 1–12. [Google Scholar] [CrossRef]
- Wu, W.; Dong, Y.; Xie, Y.; Xue, M.; Zhang, J.; Wei, H.; Ji, G.; Wang, Y. Marigold (Tagete erecta): An effective Meloidogyne incognita trap plant. Int. J. Agric. Biol. 2021, 25, 271–276. [Google Scholar] [CrossRef]
- El-Sheekh, M.M.; Mousa, A.S.H.; Farghl, A.A. Biological control of Fusarium wilt disease of tomato plants using seaweed extracts. Arab. J. Sci. Eng. 2020, 45, 4557–4570. [Google Scholar] [CrossRef]
- Uppal, A.; El Hadrami, A.; Adam, L.; Tenuta, M.; Daayf, F. Biological control of potato Verticillium wilt under controlled and field conditions using selected bacterial antagonists and plant extracts. Biol. Control 2008, 44, 90–100. [Google Scholar] [CrossRef]
- Yang, X.-x.; HUANG, X.-q.; WU, W.-x.; XIANG, Y.-j.; Lei, D.; ZHANG, L.; Yong, L. Effects of different rotation patterns on the occurrence of clubroot disease and diversity of rhizosphere microbes. J. Integr. Agric. 2020, 19, 2265–2273. [Google Scholar] [CrossRef]
- Zhang, J.; Ahmed, W.; Dai, Z.; Zhou, X.; He, Z.; Wei, L.; Ji, G. Microbial consortia: An engineering tool to suppress clubroot of Chinese cabbage by changing the rhizosphere bacterial community composition. Biology 2022, 11, 918. [Google Scholar] [CrossRef] [PubMed]
- Karlen, D.L.; Hurley, E.G.; Andrews, S.S.; Cambardella, C.A.; Meek, D.W.; Duffy, M.D.; Mallarino, A.P. Crop rotation effects on soil quality at three northern corn/soybean belt locations. Agron. J. 2006, 98, 484–495. [Google Scholar] [CrossRef]
- Venter, Z.S.; Jacobs, K.; Hawkins, H.-J. The impact of crop rotation on soil microbial diversity: A meta-analysis. Pedobiologia 2016, 59, 215–223. [Google Scholar] [CrossRef]
- Reynolds, L.B.; Potter, J.W.; Ball-Coelho, B.R. Crop rotation with Tagetes sp. is an alternative to chemical fumigation for control of root-lesion nematodes. Agron. J. 2000, 92, 957–966. [Google Scholar] [CrossRef]
- Wongsnansilp, T.; Phinrub, W.; Juntawong, N. Allelopathic effect of marigold (Tagetes erecta L.) leaf extract on growth of Chlorella vulgaris. J. Appl. Biol. Biotechnol. 2021, 10, 3–7. [Google Scholar]
- Neriman, K.; AŞKIN, M.A. Researchs on the effects of the marigold (Tagetes spp.) on the management of root-knot nematodes (Meloidogyne incognita) in nurseries. Int. J. Agric. For. Life Sci. 2018, 2, 56–61. [Google Scholar]
- Terblanche, J.D. Biological Control of Bacteria Wilt in Tobacco Caused by Ralstonia Solanacearum. Ph.D. Thesis, University of the Free State, Bloemfontein, South Africa, 2007. [Google Scholar]
- Tian, J.; Zhu, D.; Wang, J.; Wu, B.; Hussain, M.; Liu, X. Environmental factors driving fungal distribution in freshwater lake sediments across the Headwater Region of the Yellow River, China. Sci. Rep. 2018, 8, 1–8. [Google Scholar] [CrossRef]
- Bai, Y.-C.; Li, B.-X.; Xu, C.-Y.; Raza, M.; Wang, Q.; Wang, Q.-Z.; Fu, Y.-N.; Hu, J.-Y.; Imoulan, A.; Hussain, D. Intercropping Walnut and Tea: Effects on Soil Nutrients, Enzyme Activity and Microbial Communities. Front. Microbiol. 2022, 13, 560. [Google Scholar] [CrossRef]
- Zaeem, M.; Nadeem, M.; Pham, T.H.; Ashiq, W.; Ali, W.; Gilani, S.S.M.; Elavarthi, S.; Kavanagh, V.; Cheema, M.; Galagedara, L. The potential of corn-soybean intercropping to improve the soil health status and biomass production in cool climate boreal ecosystems. Sci. Rep. 2019, 9, 1–17. [Google Scholar] [CrossRef]
- Layek, J.; Das, A.; Mitran, T.; Nath, C.; Meena, R.S.; Yadav, G.S.; Shivakumar, B.; Kumar, S.; Lal, R. Cereal+ legume intercropping: An option for improving productivity and sustaining soil health. In Legumes for Soil Health and Sustainable Management; Springer: Singapore, 2018; pp. 347–386. [Google Scholar]
- Jeong, J.-Y.; Robin, A.H.K.; Natarajan, S.; Laila, R.; Kim, H.-T.; Park, J.-I.; Nou, I.-S. Race-and isolate-specific molecular marker development through genome-realignment enables detection of Korean Plasmodiophora brassicae isolates, causal agents of clubroot disease. Plant Pathol. J. 2018, 34, 506. [Google Scholar] [CrossRef]
- Zhang, H.; Yang, Y.; Mei, X.; Li, Y.; Wu, J.; Li, Y.; Wang, H.; Huang, H.; Yang, M.; He, X. Phenolic acids released in maize rhizosphere during maize-soybean intercropping inhibit Phytophthora blight of soybean. Front. Plant Sci. 2020, 11, 886. [Google Scholar] [CrossRef] [PubMed]
- Harding, M.; Hill, T.; Yang, Y.; Daniels, G.; Hwang, S.; Strelkov, S.; Howard, R.; Feng, J. An improved Evans blue staining method for consistent, accurate assessment of Plasmodiophora brassicae resting spore viability. Plant Dis. 2019, 103, 2330–2336. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Yang, Z.; He, P.; Munir, S.; Wu, Y.; Ho, H.; He, Y. Deciphering the bacterial and fungal communities in clubroot-affected cabbage rhizosphere treated with Bacillus subtilis XF-1. Agric. Ecosyst. Environ. 2018, 256, 12–22. [Google Scholar] [CrossRef]
- Li, J.; Zhao, X.; Nishimura, Y.; Fukumoto, Y. Correlation between bolting and physiological properties in Chinese cabbage (Brassica rapa L. pekinensis group). J. Jpn. Soc. Hortic. Sci. 2010, 79, 294–300. [Google Scholar] [CrossRef]
- Ahmed, W.; Dai, Z.; Liu, Q.; Munir, S.; Yang, J.; Karunarathna, S.C.; Li, S.; Zhang, J.; Ji, G.; Zhao, Z. Microbial crosstalk; Dissecting the core microbiota associated with flue-cured tobacco (Nicotiana tabacum) plants under healthy and diseased state. Front. Microbiol. 2022, 13, 397. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’hara, R.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Package ‘vegan’. Community Ecol. Package Version 2013, 2, 1–295. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Disease Incidence (%) | Disease Index | Control Effect (%) |
|---|---|---|---|
| CK | 86.67 ± 6.67 a | 63.38 ± 1.80 a | ----- |
| T1 | 71.11 ± 7.70 b | 49.84 ± 1.46 b | 21.36 ± 1.37 b |
| T2 | 46.67 ± 6.67 c | 33.33 ± 2.86 c | 47.41 ± 4.40 a |
| Sample ID | Raw Reads | Clean Reads | Average Length (bp) | Number of OTUs |
|---|---|---|---|---|
| T2-R1 | 79,899 | 79,465 | 421 | 1658 |
| T2-R2 | 79,901 | 79,507 | 421 | 1669 |
| T2-R3 | 79,795 | 79,399 | 421 | 1683 |
| T1-R1 | 80,037 | 79,623 | 420 | 1676 |
| T1-R2 | 80,172 | 79,762 | 421 | 1656 |
| T1-R3 | 80,138 | 79,741 | 421 | 1620 |
| CK-R1 | 80,116 | 79,701 | 420 | 1648 |
| CK-R2 | 80,118 | 79,723 | 421 | 1556 |
| CK-R3 | 80,049 | 79,642 | 420 | 1645 |
| Total | 720,225 | 716,563 | ------- | 14,811 |
| Average | 80,025 | 79,618 | 421 | 1646 |
| Treatments | Conditions |
|---|---|
| CK | sdH2O (100 μL/tube) + P. brassicae spore suspension (100 μL/tube) + Chinese cabbage seedling |
| T1 | marigold powder (100 μL/tube) + P. brassicae spore suspension (100 μL/tube) + Chinese cabbage seedling |
| T2 | marigold powder (100 μL/tube) + P. brassicae spore suspension (100 μL/tube) treated for 15 days + Chinese cabbage seedling |
| Treatments | Conditions |
|---|---|
| CK1 | Monocropping of Chinese cabbage |
| T1 | Chinese cabbage seedlings were transplanted immediately after harvesting the marigold crop |
| T2 | Chinese cabbage seedlings were transplanted 15 days after harvesting the marigold crop |
| Treatments | Conditions |
|---|---|
| CK | Monocropping of Chinese cabbage |
| T1 | Crop rotation (marigold crop + Chinese cabbage crop) |
| T2 | Crop rotation (marigold crop + 15 days empty period + Chinese cabbage crop) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Ahmed, W.; Zhou, X.; Yao, B.; He, Z.; Qiu, Y.; Wei, F.; He, Y.; Wei, L.; Ji, G. Crop Rotation with Marigold Promotes Soil Bacterial Structure to Assist in Mitigating Clubroot Incidence in Chinese Cabbage. Plants 2022, 11, 2295. https://doi.org/10.3390/plants11172295
Zhang J, Ahmed W, Zhou X, Yao B, He Z, Qiu Y, Wei F, He Y, Wei L, Ji G. Crop Rotation with Marigold Promotes Soil Bacterial Structure to Assist in Mitigating Clubroot Incidence in Chinese Cabbage. Plants. 2022; 11(17):2295. https://doi.org/10.3390/plants11172295
Chicago/Turabian StyleZhang, Jinhao, Waqar Ahmed, Xinghai Zhou, Bo Yao, Zulei He, Yue Qiu, Fangjun Wei, Yilu He, Lanfang Wei, and Guanghai Ji. 2022. "Crop Rotation with Marigold Promotes Soil Bacterial Structure to Assist in Mitigating Clubroot Incidence in Chinese Cabbage" Plants 11, no. 17: 2295. https://doi.org/10.3390/plants11172295