
Effects of Nitrogen Forms on the Growth and Nitrogen Accumulation in Buchloe dactyloides Seedlings

 ,
,
Abstract
:1. Introduction
2. Results
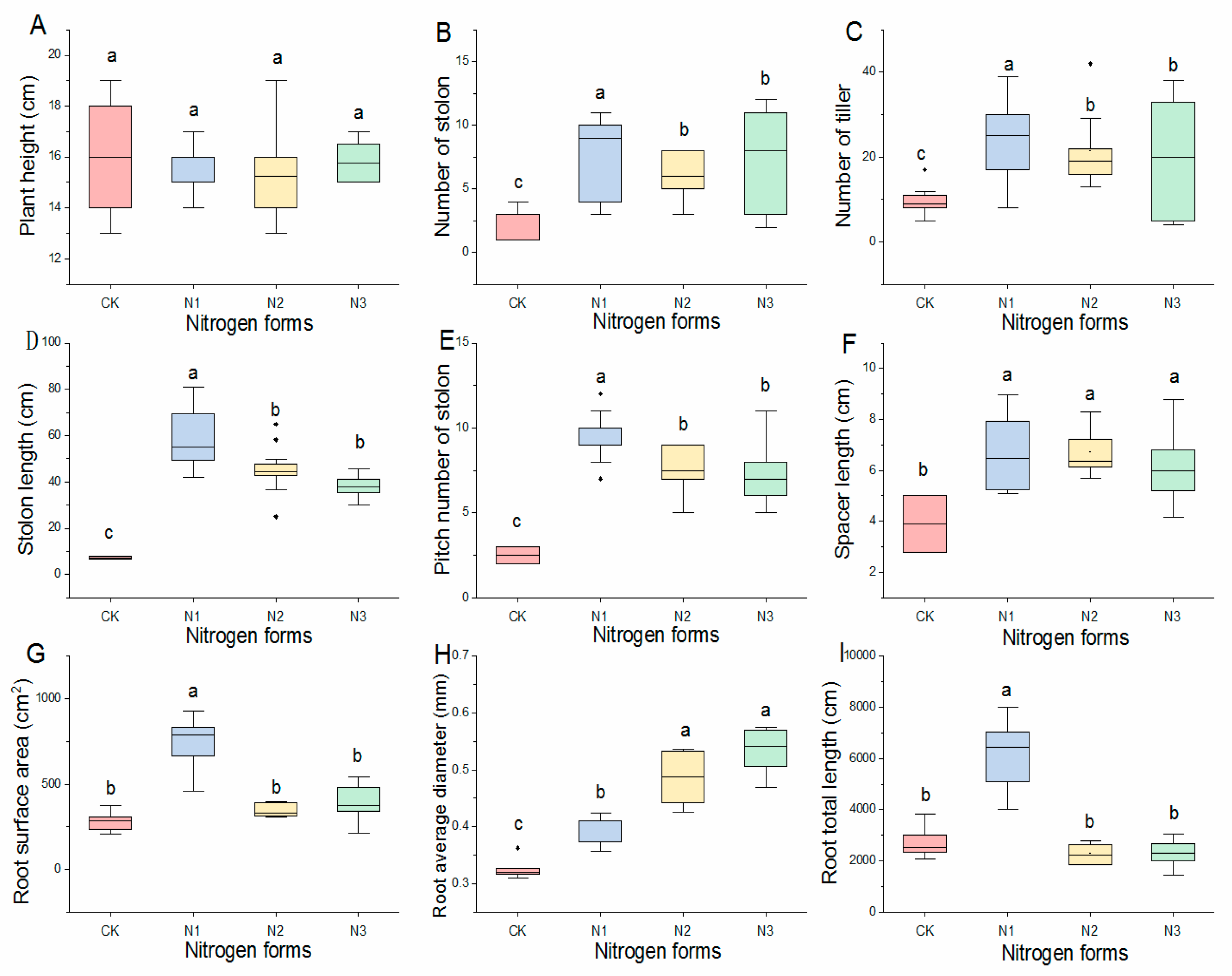
2.1. Effect of Nitrogen Form on Growth Characters
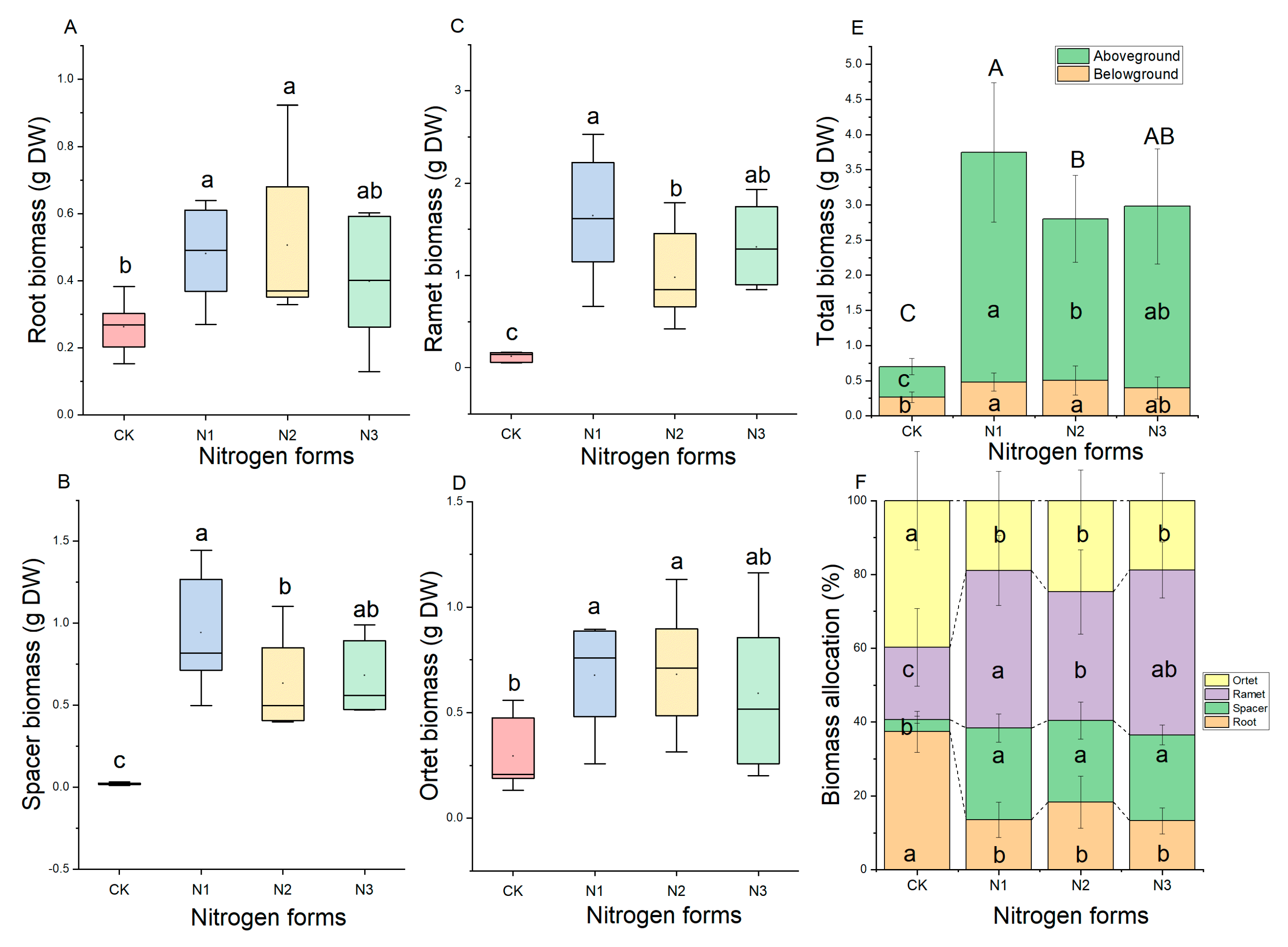
2.2. Effect of N Forms on Biomass Allocation
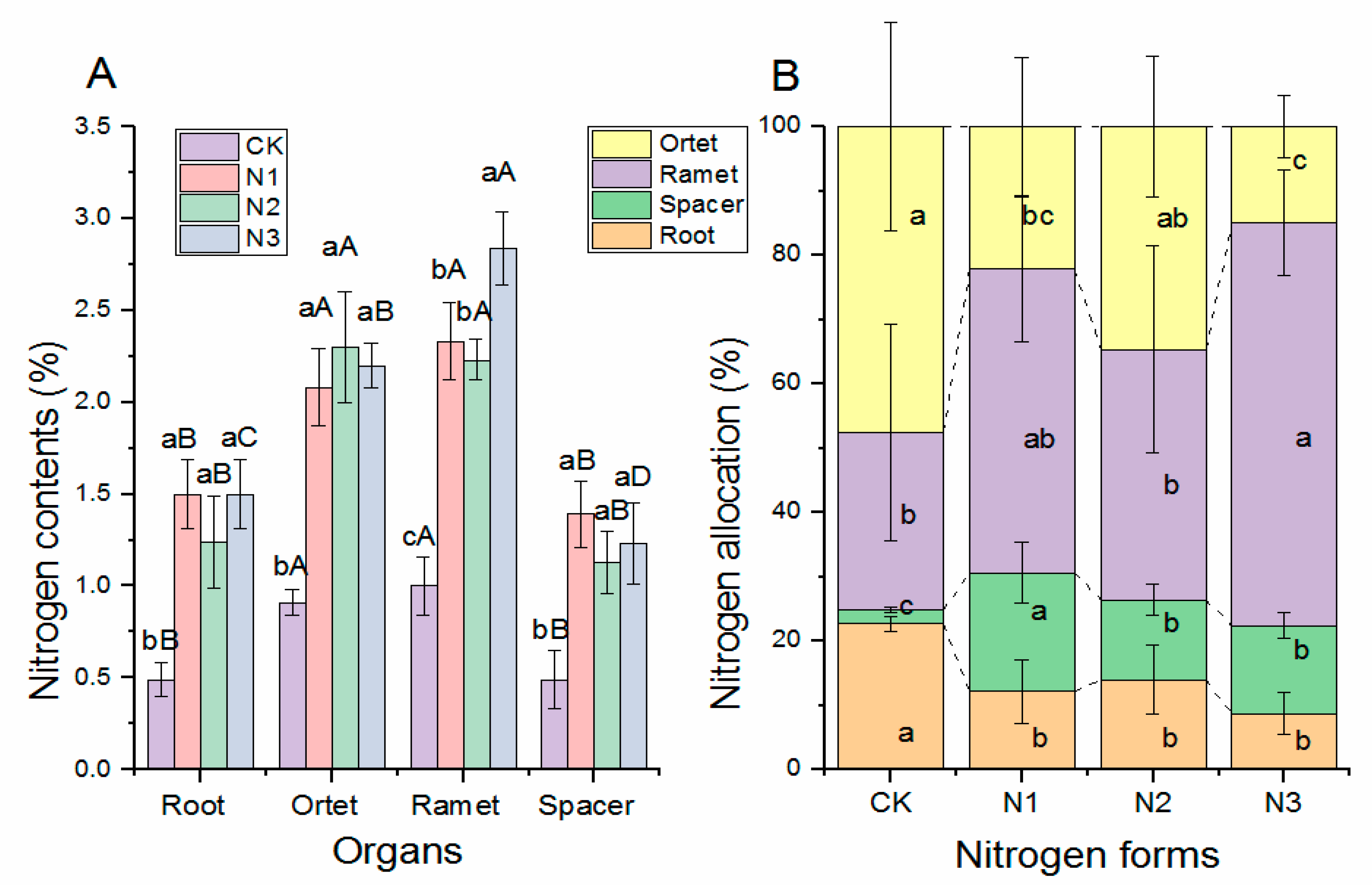
2.3. Effect of N Form on Nitrogen Contents and Allocations
2.4. Effect of N Form on 15N Enrichment and 15N Distribution
3. Discussion
4. Materials and Methods
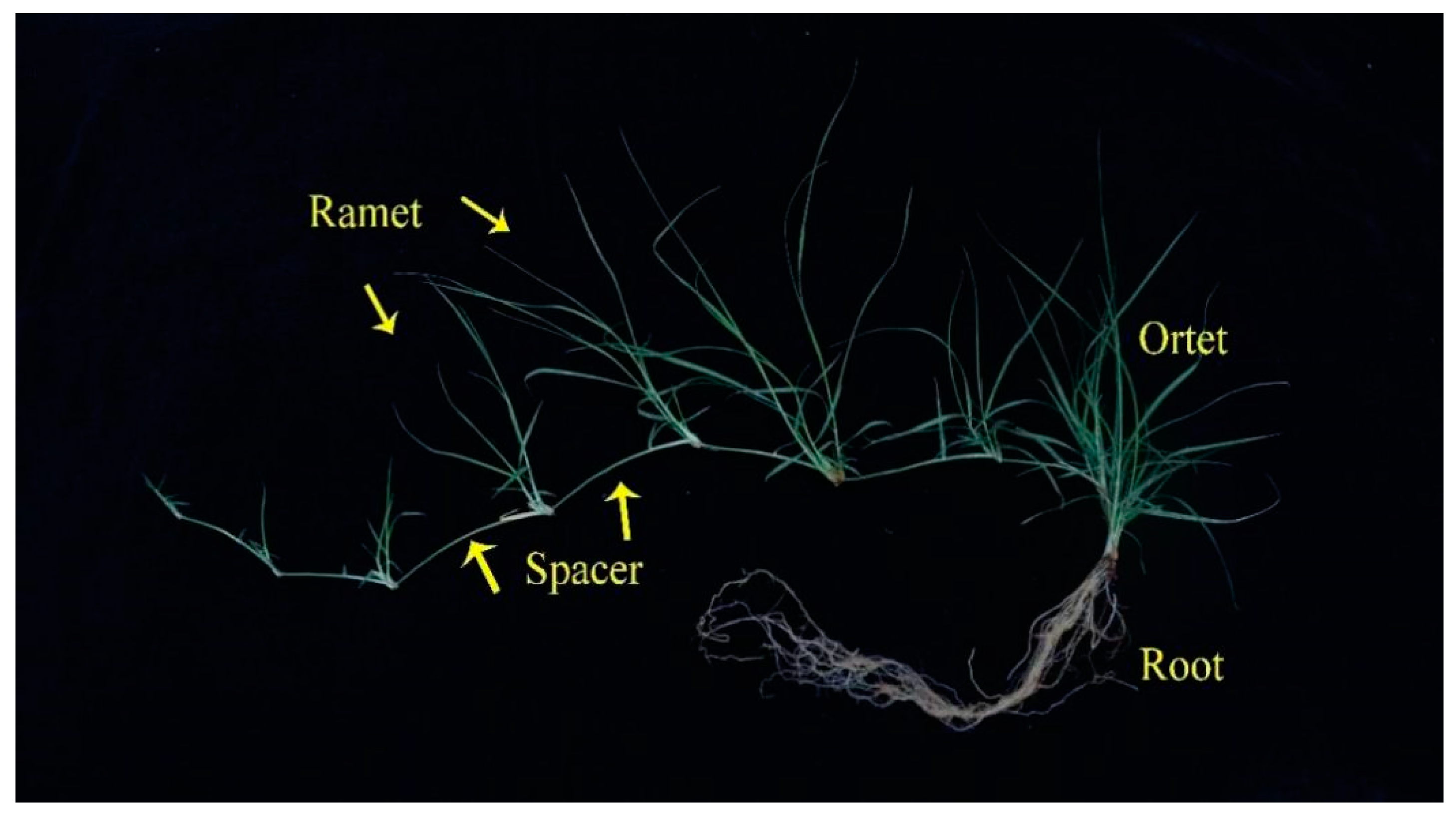
4.1. Plant Material and Experimental Design
4.2. Growth Measurements
4.3. Nitrogen Content Determination
4.4. Nitrogen Isotope Composition Determination
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kraiser, T.; Gras, D.E.; Gutierrez, A.G.; González, B.; Gutiérrez, R.A. A holistic view of nitrogen acquisition in plants. J. Exp. Bot. 2011, 62, 1455–1466. [Google Scholar] [CrossRef]
- Cambui, C.A.; Svennerstam, H.; Gruffman, L.; Nordin, A.; Ganeteg, U.; Nasholm, T. Patterns of plant biomass partitioning depend on nitrogen source. PLoS ONE 2011, 6, e19211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, G.H.; Fan, X.R.; Miller, A.J. Plant nitrogen assimilation and use efficiency. Annu. Rev. Plant Biol. 2012, 63, 153–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Britto, D.T.; Kronzucker, H.J. Ecological significance and complexity of N-source preference in plants. Ann. Bot. 2013, 112, 957–963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, N.; Huang, L.; Chen, H.J.; Huang, X.F.; Song, Q.N.; Yang, Q.P.; Wang, T.C. Nitrogen form plays an important role in the growth of moso bamboo (Phyllostachys edulis) seedlings. PeerJ 2020, 8, e9938. [Google Scholar] [CrossRef] [PubMed]
- Alt, D.S.; Doyle, J.W.; Malladi, A. Nitrogen-source preference in blueberry (Vaccinium sp.): Enhanced shoot nitrogen assimilation in response to direct supply of nitrate. J. Plant Physiol. 2017, 216, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Magalhaes, J.R.; Huber, D.M. Response of ammonium assimilation enzymes to nitrogen form treatments in different plant species. J. Plant Nutr. 1991, 14, 175–185. [Google Scholar] [CrossRef]
- Guo, S.; Zhou, Y.; Shen, Q.; Zhang, F. Effect of ammonium and nitrate nutrition on some physiological processes in higher plants—growth, photosynthesis, photorespiration, and water relations. Plant Biol. 2007, 9, 21–29. [Google Scholar] [CrossRef]
- Nacry, P.; Bouguyon, E.; Gojon, A. Nitrogen acquisition by roots: Physiological and developmental mechanisms ensuring plant adaptation to a fluctuating resource. Plant Soil 2013, 370, 1–29. [Google Scholar] [CrossRef] [Green Version]
- Hachiya, T.; Sakakibara, H. Interactions between nitrate and ammonium in their uptake, allocation, assimilation, and signaling in plants. J. Exp. Bot. 2016, 68, 2501–2512. [Google Scholar] [CrossRef] [PubMed]
- Andrews, M.; Raven, J.A.; Lea, P.J. Do plants need nitrate? The mechanisms by which nitrogen form affects plants. Ann. Appl. Biol. 2013, 163, 174–199. [Google Scholar] [CrossRef]
- Guo, S.; Brück, H.; Sattelmacher, B. Effects of supplied nitrogen form on growth and water uptake of French bean (Phaseolus vulgaris L.) plants. Plant Soil 2002, 239, 267–275. [Google Scholar] [CrossRef]
- Bauer, B.; Wirén, N.V. Modulating tiller formation in cereal crops by the signalling function of fertilizer nitrogen forms. Sci. Rep. 2020, 10, 20504. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.; Mills, H.A.; Wetzstein, H.Y. Studies on effects of nitrogen form on growth, development and nutrient uptake in pecan. J. Plant Nutr. 2002, 25, 497–508. [Google Scholar] [CrossRef]
- Guo, J.X.; Jia, Y.M.; Chen, H.H.; Zhang, L.J.; Yang, J.C.; Zhang, J.; Hu, X.Y.; Ye, X.; Li, Y.; Zhou, Y. Growth, photosynthesis, and nutrient uptake in wheat are affected by differences in nitrogen levels and forms and potassium supply. Sci. Rep. 2019, 9, 1248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qsaki, M.; Shiral, J.; Shinano, T.; Tadano, T. Effects of ammonium and nitrate assimilation on the growth and tuber swelling of potato plants. Soil Sci. Plant Nutr. 1995, 41, 709–719. [Google Scholar] [CrossRef] [Green Version]
- Tsabarducas, V.; Chatzistathis, T.; Therios, I.; Patakas, A. How nitrogen form and concentration affect growth, nutrient accumulation and photosynthetic performance of Olea europaea L. (cv. ‘Kalamon’). Sci. Hortic. 2017, 218, 23–29. [Google Scholar] [CrossRef]
- Horchani, F.; Hajri, R.; Aschi-Smiti, S. Effect of ammonium or nitrate nutrition on photosynthesis, growth, and nitrogen assimilation in tomato plants. J. Plant Nutr. Soil Sci. 2010, 173, 610–617. [Google Scholar] [CrossRef]
- Congreves, K.A.; Otchere, O.; Ferland, D.; Farzadfar, S.; Williams, S.; Arcand, M.M. Nitrogen use efficiency definitions of today and tomorrow. Front. Plant Sci. 2021, 12, 637108. [Google Scholar] [CrossRef] [PubMed]
- Andrews, M.; Lea, P.J.; Raven, J.A.; Lindsey, K. Can genetic manipulation of plant nitrogen assimilation enzymes result in increased crop yield and greater N-use efficiency? An assessment. Ann. Appl. Biol. 2004, 145, 25–40. [Google Scholar] [CrossRef]
- Andrews, M.; Raven, J.A.; Lea, P.J.; Sprent, J.I. A role for shoot protein in shoot-root dry matter allocation in higher plants. Ann. Bot. 2006, 97, 3–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, Z.B.; Li, P.; Chen, Y.H.; Han, W.X.; Fang, J.Y. Nutrient allocation strategies of woody plants: An approach from the scaling of nitrogen and phosphorus between twig stems and leaves. Sci. Rep. 2016, 6, 20099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Tang, Z.Y.; Ji, C.J.; Liu, H.Y.; Ma, W.H.; Mohhamot, A.; Shi, Z.Y.; Sun, W.; Wang, T.; Wang, X.P.; et al. Scaling of nitrogen and phosphorus across plant organs in shrubland biomes across Northern China. Sci. Rep. 2014, 4, 5778. [Google Scholar] [CrossRef] [PubMed]
- Minden, V.; Kleyer, M. Internal and external regulation of plant organ stoichiometry. Plant Biol. 2014, 16, 897–907. [Google Scholar] [CrossRef] [PubMed]
- Ågren, G.I. Stoichiometry and nutrition of plant growth in natural communities. Annu. Rev. Ecol. Evol. S. 2008, 39, 153–170. [Google Scholar] [CrossRef]
- Frank, K.W.; Gaussoin, R.E.; Fry, J.D.; Frost, M.D.; Baird, J.H. Nitrogen, phosphorus, and potassium effects on seeded buffalograss establishment. Hortscience 2002, 37, 371–373. [Google Scholar] [CrossRef] [Green Version]
- Hadle, J.J.; Russell, F.L.; Beck, J.B. Are buffalograss (Buchloë dactyloides) cytotypes spatially and ecologically differentiated? Am. J. Bot. 2019, 106, 1116–1125. [Google Scholar] [CrossRef]
- Akbarzadeh, M.; Tehranifar, A.; Alizadeh, B.; Ansari, H. The effects of regulated deficit irrigation in growth response to Büchloe dactyloides (Bowie and Cody) compared with Festuca arundinacea. J. Ornam. Plants 2018, 8, 1–11. [Google Scholar]
- Sun, H.; Zhao, X.F.; He, M.P. Analysis of ecological benefits of side slope protection by typical vegetation along the highway in Southwest areas. Res. Soil Water Conserv. 2015, 22, 6–11. (In Chinese) [Google Scholar]
- Dudley, M.M.; Jacobi, W.R.; Brown, C.S. Roadway deicer effects on the germination of native grasses and forbs. Water Air Soil Poll. 2014, 225, 1984. [Google Scholar] [CrossRef]
- Ma, Z.L. Establishment, management and protection technology of Buffalo Grass in canal slope protection in the middle route of South-to-North Water Diversion Project. Technol. Soil Water Conserv. 2018, 186, 31–33. (In Chinese) [Google Scholar]
- Honeker, L.K.; Neilson, J.W.; Root, R.A.; Gil-loaiza, J.; Chorover, J.; Maier, R.M. Bacterial rhizoplane colonization patterns of Buchloe dactyloides growing in metalliferous mine tailings reflect plant status and biogeochemical conditions. Microb. Ecol. 2017, 74, 853–867. [Google Scholar] [CrossRef] [PubMed]
- Honeker, L.K.; Neilson, J.W.; Chorover, J.; Maier, R.M. Effect of re-acidification on buffalo grass rhizosphere and bulk microbial communities during phytostabilization of metalliferous mine tailings. Front. Microbiol. 2019, 10, 1209. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.M. Experimental study on the nutrition bowl of Buffalo Grass in the capital airport flight area. Civil Airports 2018, 337, 66–68. (In Chinese) [Google Scholar]
- Frank, K.W.; Gaussoin, R.E.; Riordan, T.P.; Shearman, R.C.; Johnson, P.G. Nitrogen rate and mowing height effects on turf-type Buffalograss. Crop Sci. 2004, 44, 1615–1621. [Google Scholar] [CrossRef]
- Alderman, E.J.; Hoyle, J.A.; Keeley, S.J.; Fry, J.D. Buffalograss divot recovery as affected by nitrogen source and rate. Crop Forage Turfgrass Manag. 2017, 3, 1–6. [Google Scholar] [CrossRef]
- Springer, T.L.; Taliaferro, C.M.; Hattey, J.A. Nitrogen source and rate effects on the production of buffalograss forage grown with irrigation. Crop Sci. 2005, 45, 668–672. [Google Scholar] [CrossRef]
- Glinski, D.S.; Mills, H.A.; Karnok, K.J.; Carrow, R.N. Nitrogen form influences root growth of sodded creeping bentgrass. Hortscience 1990, 25, 932–933. [Google Scholar] [CrossRef] [Green Version]
- McCrimmon, J.N.; Karnok, K.J.; Mills, H.A. Effect of Nitrogen-Form on Plant Growth and Nutrient Composition of Creeping Bentgrass. In Optimization of Plant Nutrition; Fragoso, M.A.C., Van Beusichem, M.L., Houwers, A., Eds.; Springer: Dordrecht, The Netherlands, 1993; pp. 291–297. [Google Scholar]
- Giehl, R.F.H.; Gruber, B.D.; von Wirén, N. It’s time to make changes: Modulation of root system architecture by nutrient signals. J. Exp. Bot. 2014, 65, 769–778. [Google Scholar] [CrossRef] [PubMed]
- Britto, D.T.; Kronzucker, H.J. NH4+ toxicity in higher plants: A critical review. J. Plant Physiol. 2002, 159, 567–584. [Google Scholar] [CrossRef] [Green Version]
- Esteban, R.; Ariz, I.; Cruz, C.; Moran, J.F. Review: Mechanisms of ammonium toxicity and the quest for tolerance. Plant Sci. 2016, 248, 92–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roosta, H.R.; Schjoerring, J.K. Effects of nitrate and potassium on ammonium toxicity in cucumber plants. J. Plant Nutr. 2008, 31, 1270–1283. [Google Scholar] [CrossRef]
- Ten Hoopen, F.; Cuin, T.A.; Pedas, P.; Hegelund, J.N.; Shabala, S.; Schjoerring, J.K.; Jahn, T.P. Competition between uptake of ammonium and potassium in barley and Arabidopsis roots: Molecular mechanisms and physiological consequences. J. Exp. Bot. 2010, 61, 2303–2315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Britto, D.T.; Kronzucker, H.J. Plant nitrogen transport and its regulation in changing soil environments. J. Crop Improv. 2006, 15, 1–23. [Google Scholar] [CrossRef]
- Li, B.; Li, G.; Kronzucker, H.J.; Baluška, F.; Shi, W. Ammonium stress in Arabidopsis: Signaling, genetic loci, and physiological targets. Trends Plant Sci. 2014, 19, 107–114. [Google Scholar] [CrossRef]
- Tucker, M. Primary Nutrients and Plant Growth. In Essential Plant Nutrients; Scribd, Ed.; North Carolina Department of Agriculture: Raleigh, NC, USA, 2004. [Google Scholar]
- Onoda, Y.; Hirose, K.H. Allocation of nitrogen to cell walls decreases photosynthetic nitrogen-use efficiency. Funct. Ecol. 2004, 18, 419–425. [Google Scholar] [CrossRef]
- Tinker, P.B.; Nye, P.H. Solute Movement in the Rhizosphere; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Yan, B.G.; Fan, Z.H.; Wang, X.M.; He, G.X.; Shi, L.T.; Liu, G.C. Plants adapted to nutrient limitation allocate less biomass into stems in an arid-hot grassland. New Phytol. 2016, 211, 1232–1240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hermans, C.; Hammond, J.P.; White, P.J.; Verbruggen, N. How do plants respond to nutrient shortage by biomass allocation? Trends Plant Sci. 2006, 11, 610–617. [Google Scholar] [CrossRef] [PubMed]
- Grechi, I.; Vivin, P.; Hilbert, G.; Milin, S.; Robert, T.; Gaudillere, J.P. Effect of light and nitrogen supply on internal C:N balance and control of root-to-shoot biomass allocation in grapevine. Environ. Exp. Bot. 2007, 59, 139–149. [Google Scholar] [CrossRef]
- Shaver, G.R.; Chapin, F.S., III. Production: Biomass relationships and element cycling in contrasting arctic vegetation types. Ecol. Monogr. 1991, 61, 1–31. [Google Scholar] [CrossRef]
- Tilman, D.; Wedin, D. Plant traits and resource reduction for five grasses growing on a nitrogen gradient. Ecology 1991, 72, 685–700. [Google Scholar] [CrossRef]
- Epstein, E.; Bloom, A.J. Mineral Nutrition of Plants: Principles and Perspectives, 2nd ed.; Sinauer Associates Inc.: Sunderland, MA, USA, 2005. [Google Scholar]
- Yoneyama, T.; Ito, O.; Engelaar, W.M.H.G. Uptake, metabolism, and distribution of nitrogen in crop plants traced by enriched and natural 15N: Progress over the last 30 years. Phytochem. Rev. 2003, 2, 121–132. [Google Scholar] [CrossRef]
- Bedard-Haughn, A.; Groenigen, J.W.V.; Kessel, C.V. Tracing 15N through landscapes: Potential uses and precautions. J. Hydrol. 2003, 272, 175–190. [Google Scholar] [CrossRef]
- Hill, T.D.; Sommer, N.R.; Kanaskie, C.R.; Santos, E.A.; Oczkowski, A.J. Nitrogen uptake and allocation estimates for Spartina alterniflora and Distichlis spicata. J. Exp. Mar. Biol. Ecol. 2018, 507, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Barickman, T.C.; Kopsell, D.A. Nitrogen form and ratio impact Swiss chard (Beta vulgaris subsp. cicla) shoot tissue carotenoid and chlorophyll concentrations. Sci. Hortic. 2016, 204, 99–105. [Google Scholar] [CrossRef]
- Borrero, C.; Trillas, M.I.; Delgado, A.; Avile´s, M. Effect of ammonium/nitrate ratio in nutrient solution on control of Fusarium wilt of tomato by (Trichoderma asperellum) T34. Plant Pathology 2012, 61, 132–139. [Google Scholar] [CrossRef]
- O’Callaghan, M.; Gerard, E.M.; Carter, P.E.; Lardner, R.; Sarathchandra, U.; Burch, G.; Ghani, A.; Bell, N. Effect of the nitrification inhibitor dicyandiamide (DCD) on microbial communities in a pasture soil amended with bovine urine. Soil Biol. Biochem. 2010, 42, 1425–1436. [Google Scholar] [CrossRef]
- Di, H.J.; Cameron, K.C.; Sherlock, R.R. Comparison of the effectiveness of a nitrification inhibitor, dicyandiamide, in reducing nitrous oxide emissions in four different soils under different climatic and management conditions. Soil Use Manag. 2007, 23, 1–9. [Google Scholar] [CrossRef]
- Monaghan, R.M.; Smith, L.C.; Ledgard, S.F. The effectiveness of a granular formulation of dicyandiamide (DCD) in limiting nitrate leaching from a grazed dairy pasture. N. Z. J. Agr. Res. 2009, 52, 145–159. [Google Scholar] [CrossRef]
- Yousfi, S.; Serret, M.D.; Marquez, A.J.; Voltas, J.; Araus, J.L. Combined use of δ13C, δ18O and δ15N tracks nitrogen metabolism and genotypic adaptation of durum wheat to salinity and water deficit. New Phytol. 2012, 194, 230–244. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factors | df | N Contents | N Allocation | ||
|---|---|---|---|---|---|
| F | p | F | p | ||
| Organs (O) | 3 | 144.162 | <0.01 | 53.658 | <0.01 |
| Treatments (T) | 3 | 120.894 | <0.01 | 0.002 | 1 |
| O ∗ T | 9 | 5.627 | <0.01 | 8.299 | <0.01 |
| Organs | |||||
|---|---|---|---|---|---|
| Root | Spacer | Ramet | Ortet | ||
| 15N abundance (atom %) | N1 | 4.22 ± 0.13 b | 4.53 ± 0.23 b | 4.40 ± 0.39 bB | 5.09 ± 0.10 a |
| N2 | 4.39 ± 0.48 b | 4.94 ± 0.85 ab | 4.89 ± 0.80 abAB | 5.64 ± 0.60 a | |
| N3 | 4.66 ± 0.51 b | 5.45 ± 0.86 ab | 5.48 ± 0.63 abA | 5.93 ± 0.72 a | |
| Ndff (%) | N1 | 3.91 ± 0.13 b | 4.22 ± 0.23 b | 4.09 ± 0.40 bB | 4.79 ± 0.11 a |
| N2 | 4.08 ± 0.48 b | 4.64 ± 0.86 ab | 4.59 ± 0.82 abAB | 5.35 ± 0.60 a | |
| N3 | 4.35 ± 0.52 b | 5.15 ± 0.87 ab | 5.19 ± 0.64 abA | 5.65 ± 0.73 a | |
| 15N accumulation amount (mg/plant) | N1 | 3.45 ± 1.14 | 3.68 ± 1.00 | 3.54 ± 0.90 | 4.24 ± 1.46 |
| N2 | 2.52 ± 0.47 b | 2.83 ± 0.37 ab | 2.80 ± 0.32 ab | 3.30 ± 0.52 a | |
| N3 | 3.41 ± 1.26 | 4.05 ± 1.58 | 4.07 ± 1.47 | 4.44 ± 1.67 | |
| 15N distribution (%) | N1 | 22.98 ± 0.77 c | 24.80 ± 0.66 b | 23.99 ± 1.50 bc | 28.23 ± 1.30 a |
| N2 | 21.94 ± 1.00 c | 24.78 ± 1.49 b | 24.53 ± 0.91 b | 28.75 ± 1.00 a | |
| N3 | 21.44 ± 0.81 c | 25.24 ± 1.00 b | 25.54 ± 0.88 b | 27.78 ± 0.38 a | |
| Treatment (mM) | CK | N1 | N2 | N3 |
|---|---|---|---|---|
| -NH4NO3 | +NO3− | +NH4+ | +NH4NO3 | |
| KCL | 1.25 | 0 | 1.5 | 0.25 |
| CaCL2·6H2O | 1.25 | 0 | 1.25 | 0 |
| MgSO4·7H2O | 0.5 | 0.5 | 0.5 | 0.5 |
| KH2(PO4) | 0.25 | 0.25 | 0 | 0 |
| KNO3 | 0 | 1.25 | 0 | 1.25 |
| Ca(NO3)2·4H2O | 0 | 1.25 | 0 | 1.25 |
| NH4H2PO4 | 0 | 0 | 0.25 | 0.25 |
| NH4CL | 0 | 0 | 3.5 | 3.5 |
| H3BO3 | 0.0116 | 0.0116 | 0.0116 | 0.0116 |
| MnCL2·4H2O | 0.0046 | 0.0046 | 0.0046 | 0.0046 |
| ZnSO4·7H2O | 0.00019 | 0.00019 | 0.00019 | 0.00019 |
| Na2MoO4 | 0.00012 | 0.00012 | 0.00012 | 0.00012 |
| CuSO4·5H2O | 0.00008 | 0.00008 | 0.00008 | 0.00008 |
| FeSO4·7H2O | 0.0125 | 0.0125 | 0.0125 | 0.0125 |
| Na2EDTA | 0.0125 | 0.0125 | 0.0125 | 0.0125 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, L.; Meng, H.; Teng, K.; Fan, X.; Zhang, H.; Teng, W.; Yue, Y.; Wu, J. Effects of Nitrogen Forms on the Growth and Nitrogen Accumulation in Buchloe dactyloides Seedlings. Plants 2022, 11, 2086. https://doi.org/10.3390/plants11162086
Guo L, Meng H, Teng K, Fan X, Zhang H, Teng W, Yue Y, Wu J. Effects of Nitrogen Forms on the Growth and Nitrogen Accumulation in Buchloe dactyloides Seedlings. Plants. 2022; 11(16):2086. https://doi.org/10.3390/plants11162086
Chicago/Turabian StyleGuo, Lizhu, Huizhen Meng, Ke Teng, Xifeng Fan, Hui Zhang, Wenjun Teng, Yuesen Yue, and Juying Wu. 2022. "Effects of Nitrogen Forms on the Growth and Nitrogen Accumulation in Buchloe dactyloides Seedlings" Plants 11, no. 16: 2086. https://doi.org/10.3390/plants11162086