
Maturation and Post-Harvest Resting of Fruits Affect the Macronutrients and Protein Content in Sweet Pepper Seeds

 , and
, and
Abstract
:1. Introduction
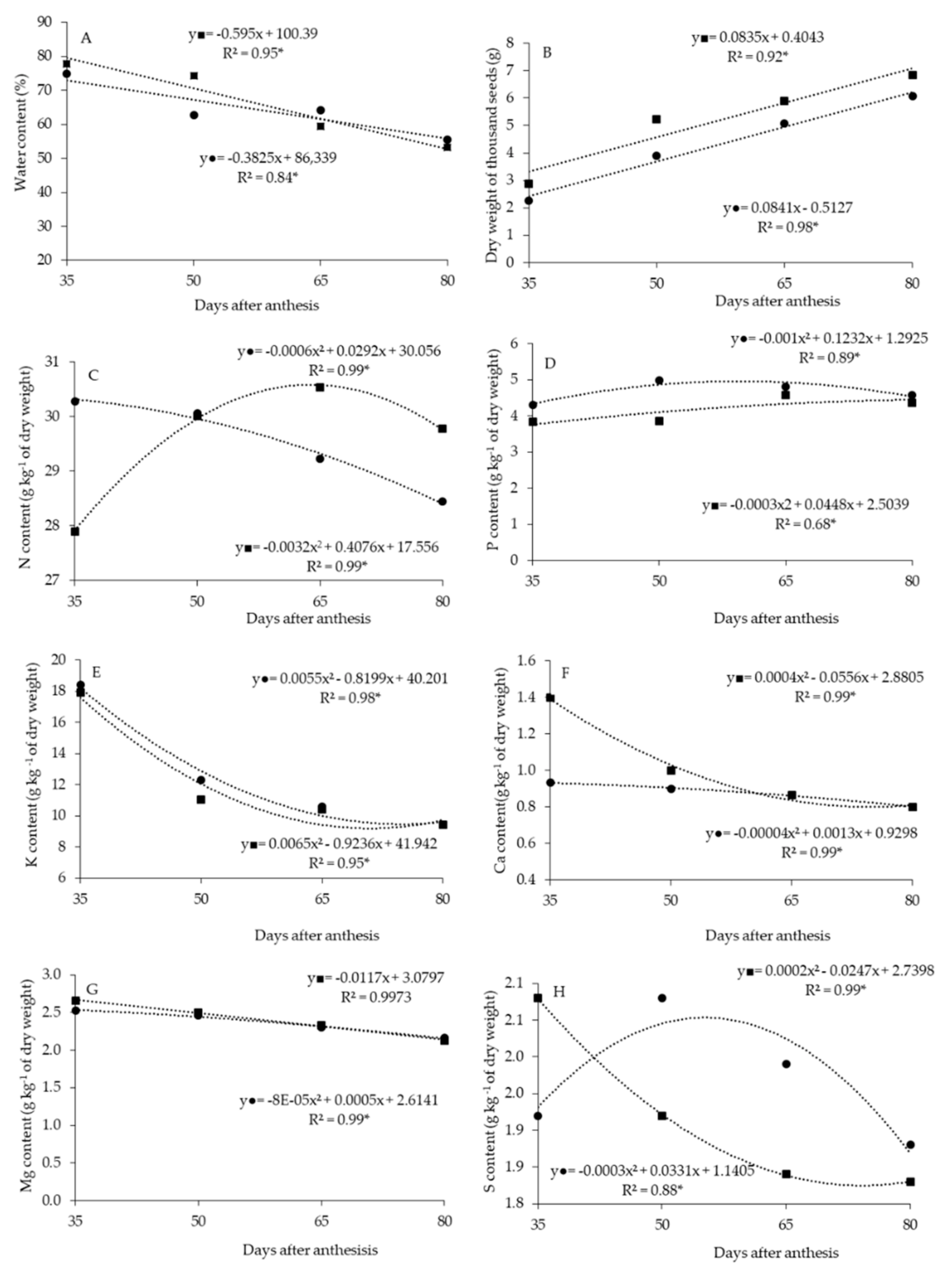
2. Results
2.1. Water Content, Dry Weight of One Thousand and Macronutrient Content of Seeds
2.2. Seed Protein Content
3. Discussion
3.1. Water Content, Dry Weight of One Thousand and Macronutrients Content of Seeds
3.2. Seed Protein Content
4. Materials and Methods
4.1. Site Description
4.2. Experiment Conduction and Experimental Design
4.3. Seed Analysis
4.4. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aguirre, M.; Kiegle, E.; Leo, G.; Ezquer, I. Carbohydrate reserves and seed development: An overview. Plant Reprod. 2018, 31, 263–290. [Google Scholar] [CrossRef] [PubMed]
- Bang, T.C.; Husted, S.; Laursen, K.H.; Persson, D.P.; Schjoerring, J.K. The molecular–physiological functions of mineral macronutrients and their consequences for deficiency symptoms in plants. New Phytol. 2021, 229, 2446–2469. [Google Scholar] [CrossRef] [PubMed]
- Ikram, A.; Saeed, F.; Afzaal, M.; Imran, A.; Niaz, B.; Tufail, T.; Hussain, M.; Anjum, F.M. Nutritional and end-use perspectives of sprouted grains: A comprehensive review. Food Sci. Nutr. 2021, 9, 4617–4628. [Google Scholar] [CrossRef] [PubMed]
- Kołton, A.; Wojciechowska, R.; Leja, M. The effect of various light conditions and different nitrogen forms on nitrogen metabolism in pepper fruits. Folia Hortic. 2012, 24, 153–160. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Lv, J.; Dawuda, M.M.; Xie, J.; Yu, J.; Li, J.; Zhang, X.; Tang, C.; Wang, C.; Gan, Y. Appropriate ammonium-nitrate ratio improves nutrient accumulation and fruit quality in pepper (Capsicum annuum L.). Agronomy 2019, 9, 683. [Google Scholar] [CrossRef] [Green Version]
- Mouzo, D.; Bernal, J.; López-Pedrouso, M.; Franco, D.; Zapata, C. Advances in the biology of seed and vegetative storage proteins based on two-dimensional electrophoresis coupled to mass spectrometry. Molecules 2018, 23, 2462. [Google Scholar] [CrossRef] [Green Version]
- Shewry, P.R.; Napier, J.A.; Tatham, A.S. Seed storage proteins: Structures and biosynthesis. Plant Cell 1995, 7, 945–956. [Google Scholar] [CrossRef] [Green Version]
- Tan-Wilson, A.L.; Wilson, K.A. Mobilization of seed protein reserves. Physiol. Plant. 2012, 145, 140–153. [Google Scholar] [CrossRef]
- Koornneef, M.; Bentsink, L.; Hilhorst, H. Seed dormancy and germination. Curr. Opin. Plant Biol. 2002, 5, 33–36. [Google Scholar] [CrossRef] [Green Version]
- Han, C.; Yang, P. Studies on the molecular mechanisms of seed germination. Proteomics 2015, 15, 1671–1679. [Google Scholar] [CrossRef]
- Nakada-Freitas, P.G.; Lanna, N.B.L.; Silva, P.N.L.; Bardiviesso, E.M.; Tavares, A.E.B.; Claudio, M.T.R.; Cardoso, A.I.I.; Magro, F.O.; Araujo, H.S. The physiological quality of “chilli pepper” seeds, extracted from fruits harvested at different stages of maturation, with and without post-harvest rest. Aust. J. Crop Sci. 2020, 14, 739–743. [Google Scholar] [CrossRef]
- Colombari, L.F.; da Silva, G.F.; Chamma, L.; Chaves, P.P.N.; Martins, B.N.M.; Jorge, L.G.; de Lima Silva, P.N.; Putti, F.F.; Cardoso, A.I.I. Maturation and resting of sweet pepper fruits on physiological quality and biochemical response of seeds. Brazilian Arch. Biol. Technol. 2021, 64, e21200733. [Google Scholar] [CrossRef]
- Medeiros, A.D.; Zavala-León, M.J.; da Silva, L.J.; Oliveira, A.M.S.; dos Santos Dias, D.C.F. Relationship between internal morphology and physiological quality of pepper seeds during fruit maturation and storage. Agron. J. 2020, 112, 25–35. [Google Scholar] [CrossRef]
- Ohanenye, I.C.; Tsopmo, A.; Ejike, C.E.C.C.; Udenigwe, C.C. Germination as a bioprocess for enhancing the quality and nutritional prospects of legume proteins. Trends Food Sci. Technol. 2020, 101, 213–222. [Google Scholar] [CrossRef]
- Cisternas-Jamet, J.; Salvatierra-Martínez, R.; Vega-Gálvez, A.; Stoll, A.; Uribe, E.; Goñi, M.G. Biochemical composition as a function of fruit maturity stage of bell pepper (Capsicum annum) inoculated with Bacillus amyloliquefaciens. Sci. Hortic. 2020, 263, 109107. [Google Scholar] [CrossRef]
- Yildirim, K.C.; Canik Orel, D.; Okyay, H.; Gursan, M.M.; Demir, I. Quality of immature and mature pepper (Capsicum annuum L.) seeds in relation to bio-priming with endophytic pseudomonas and Bacillus spp. Horticulturae 2021, 7, 75. [Google Scholar] [CrossRef]
- Bewley, J.D.; Bradford, K.J.; Hilhorst, H.W.M.; Nonogaki, H. Seeds; Springer: New York, NY, USA, 2013. [Google Scholar]
- Wolny, E.; Betekhtin, A.; Rojek, M.; Braszewska-Zalewska, A.; Lusinska, J.; Hasterok, R. Germination and the early stages of seedling development in Brachypodium distachyon. Int. J. Mol. Sci. 2018, 19, 2916. [Google Scholar] [CrossRef] [Green Version]
- Linkies, A.; Leubner-Metzger, G. Beyond gibberellins and abscisic acid: How ethylene and jasmonates control seed germination. Plant Cell Rep. 2012, 31, 253–270. [Google Scholar] [CrossRef]
- Yan, D.; Duermeyer, L.; Leoveanu, C.; Nambara, E. The functions of the endosperm during seed germination. Plant Cell Physiol. 2014, 55, 1521–1533. [Google Scholar] [CrossRef]
- Bortey, H.M.; Dzomeku, B.M. Fruit and seed quality of okra [Abelmoschus esculentus (L.) Moench] as influenced by harvesting stage and drying method. Indian J. Agric. Res. 2016, 50, 330–334. [Google Scholar] [CrossRef]
- Singkaew, J.; Miyagawa, S.; Wongs-Aree, C.; Vichitsoonthonkul, T.; Sokaokha, S.; Photchanachai, S. Season, fruit maturity, and storage affect on the physiological quality of F1 hybrid ‘VTM580’ tomato seeds and seedlings. Hortic. J. 2017, 86, 121–131. [Google Scholar] [CrossRef] [Green Version]
- Lima, J.M.E.; Smiderle, O.J. Physiologic quality of pepper seeds obtained from to fruit maturation and storage and storage. Semin. Ciências Agrárias 2014, 35, 251. [Google Scholar] [CrossRef]
- Vidigalde Souza Vidigal, D.; Dias, D.C.F.S.; de Rezende Von Pinho, E.V.; dos Santos Dias, L.A. Physiological and enzymatic changes during pepper seeds (Capsicum annuum L.) maturation. Rev. Bras. Sementes 2009, 31, 129–136. [Google Scholar] [CrossRef]
- Caixeta, F.; Von Pinho, É.V.R.; Catão, R.M.G.; Pereira, P.H.A.R.; Catão, H.C.R.M. Physiological and biochemical alterations during germination and storage of habanero pepper seeds. African J. Agric. Res. 2014, 9, 627–635. [Google Scholar] [CrossRef] [Green Version]
- Nogueira, J.L.; da Silva, B.A.; Mógor, Á.F.; de Souza Grzybowski, C.R.; Panobianco, M. Quality of organically produced bell pepper seeds. J. Seed Sci. 2017, 39, 100–105. [Google Scholar] [CrossRef] [Green Version]
- Pinheiro, D.T.; de Oliveira, R.M.; de Souza Silveira, A.; León, M.J.Z.; Brum, L.B.T.L.; dos Santos Dias, D.C.F. Antioxidant enzyme activity and physiological potential of Capsicum baccatum var. baccatum seeds as a function of post-harvest storage of fruit. J. Seed Sci. 2020, 42, e202042028. [Google Scholar] [CrossRef]
- de Souza Vidigal, D.; dos Santos Dias, D.C.F.; dos Santos Dias, L.A.; Finger, F.L. Changes in seed quality during fruit maturation of sweet pepper. Sci. Agric. 2011, 68, 535–539. [Google Scholar] [CrossRef] [Green Version]
- Kano, C.; Cardoso, A.I.I.; Villas Bôas, R.L. Macronutrient content in lettuce affected by potassium side dressing. Hortic. Bras. 2010, 28, 287–291. [Google Scholar] [CrossRef] [Green Version]
- Cardoso, A.I.; Claudio, M.T.; Nakada-Freitas, P.G.; Magro, F.O.; Tavares, A.E. Phosphate fertilization over the accumulation of macronutrients in cauliflower seed production. Hortic. Bras. 2016, 34, 196–201. [Google Scholar] [CrossRef] [Green Version]
- Masclaux-Daubresse, C.; Daniel-Vedele, F.; Dechorgnat, J.; Chardon, F.; Gaufichon, L.; Suzuki, A. Nitrogen uptake, assimilation and remobilization in plants: Challenges for sustainable and productive agriculture. Ann. Bot. 2010, 105, 1141–1157. [Google Scholar] [CrossRef] [Green Version]
- Sandhu, N.; Sethi, M.; Kumar, A.; Dang, D.; Singh, J.; Chhuneja, P. Biochemical and genetic approaches improving nitrogen use efficiency in cereal crops: A review. Front. Plant Sci. 2021, 12, 657629. [Google Scholar] [CrossRef] [PubMed]
- Doyle, J.W.; Nambeesan, S.U.; Malladi, A. Physiology of nitrogen and calcium nutrition in blueberry (Vaccinium sp.). Agronomy 2021, 11, 765. [Google Scholar] [CrossRef]
- Marschner, P. Mineral Nutrition of Higher Plants; Academic Press: London, UK, 2012. [Google Scholar]
- Schachtman, D.P.; Reid, R.J.; Ayling, S.M. Phosphorus uptake by plants: From soil to cell. Plant Physiol. 1998, 116, 447–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, P.J.; Veneklaas, E.J. Nature and nurture: The importance of seed phosphorus content. Plant Soil 2012, 357, 1–8. [Google Scholar] [CrossRef]
- Görlach, B.M.; Sagervanshi, A.; Henningsen, J.N.; Pitann, B.; Mühling, K.H. Uptake, subcellular distribution, and translocation of foliar-applied phosphorus: Short-term effects on ion relations in deficient young maize plants. Plant Physiol. Biochem. 2021, 166, 677–688. [Google Scholar] [CrossRef]
- Lemaire, G.; Sinclair, T.; Sadras, V.; Bélanger, G. Allometric approach to crop nutrition and implications for crop diagnosis and phenotyping. A review. Agron. Sustain. Dev. 2019, 39, 27. [Google Scholar] [CrossRef] [Green Version]
- Sawan, Z.M.; Fahmy, A.H.; Yousef, S.E. Effect of potassium, zinc and phosphorus on seed yield, seed viability and seedling vigor of cotton (Gossypium barbadense L.). Arch. Agron. Soil Sci. 2011, 57, 75–90. [Google Scholar] [CrossRef]
- Ceylan, Y.; Kutman, U.B.; Mengutay, M.; Cakmak, I. Magnesium applications to growth medium and foliage affect the starch distribution, increase the grain size and improve the seed germination in wheat. Plant Soil 2016, 406, 145–156. [Google Scholar] [CrossRef]
- Chandra, N.; Pandey, N. Role of sulfur nutrition in plant and seed metabolism of Glycine max L. J. Plant Nutr. 2016, 39, 1103–1111. [Google Scholar] [CrossRef]
- Mondal, S.; Pramanik, K.; Panda, D.; Dutta, D.; Karmakar, S.; Bose, B. Sulfur in Seeds: An Overview. Plants 2022, 11, 450. [Google Scholar] [CrossRef]
- Dos Reis, A.R.; Boleta, E.H.M.; Alves, C.Z.; Cotrim, M.F.; Barbosa, J.Z.; Silva, V.M.; Porto, R.L.; Lanza, M.G.D.B.; Lavres, J.; Gomes, M.H.F.; et al. Selenium toxicity in upland field-grown rice: Seed physiology responses and nutrient distribution using the μ-XRF technique. Ecotoxicol. Environ. Saf. 2020, 190, 110147. [Google Scholar] [CrossRef]
- Ramakrishna, V. Mobilization of albumins and globulins during germination of Indian bean (Dolichos lablab L. var. lignosus) seeds. Acta Bot. Croat. 2007, 66, 135–142. [Google Scholar]
- Zhao, M.; Zhang, H.; Yan, H.; Qiu, L.; Baskin, C.C. Mobilization and role of starch, protein, and fat reserves during seed germination of six wild grassland species. Front. Plant Sci. 2018, 9, 234. [Google Scholar] [CrossRef] [Green Version]
- Schlereth, A.; Standhardt, D.; Mock, H.-P.; Müntz, K. Stored cysteine proteinases start globulin mobilization in protein bodies of embryonic axes and cotyledons during vetch (Vicia sativa L.) seed germination. Planta 2001, 212, 718–727. [Google Scholar] [CrossRef]
- Müntz, K.; Belozersky, M.A.; Dunaevsky, Y.E.; Schlereth, A.; Tiedemann, J. Stored proteinases and the initiation of storage protein mobilization in seeds during germination and seedling growth. J. Exp. Bot. 2001, 52, 1741–1752. [Google Scholar] [CrossRef] [Green Version]
- Trani, P.E.; Tivelli, S.W.; Carrijo, O.A. Fertirrigação em Hortaliças, 2nd ed.; Instituto Agronômico: Campinas, Brazil, 2011. [Google Scholar]
- Brasil. Regras Para Análise de Sementes; MAPA/ACS: Brasília, Brazil, 2009; 399p. [Google Scholar]
- AOAC. Official Methods of Analysis of AOAC International; AOAC International: Rockville, MD, USA, 2016. [Google Scholar]
- Bradford, M. A Rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Post-Harvest Resting | DWTS | P | K | Ca | Mg |
|---|---|---|---|---|---|
| g kg−1 of Dry Matter | |||||
| Without | 4.3b | 4.7a | 12.4a | 0.87b | 2.4a |
| With | 5.2a | 4.4b | 12.3a | 1.02a | 2.4a |
| CV (%) | 2.5 | 1.41 | 4.07 | 8.79 | 7.10 |
| Post-Harvest Resting | N Content (g kg−1 of Dry Matter) | S Content (g kg−1 of Dry Matter) | ||||||
|---|---|---|---|---|---|---|---|---|
| Days after Anthesis | ||||||||
| 35 | 50 | 65 | 80 | 35 | 50 | 65 | 80 | |
| Without | 30a | 30a | 29b | 28b | 1.9b | 2.1a | 2.0a | 1.9a |
| With | 28b | 30a | 31a | 30a | 2.1a | 1.9b | 1.8b | 1.8a |
| CV (%) | 3.65 | 6.43 | ||||||
| Post-Harvest Resting | Albumin | Globulin | Prolamine | Glutelin |
|---|---|---|---|---|
| mg g−1 of Dry Matter | ||||
| Without | 59.6b | 17.5a | 6.9a | 13.5b |
| With | 61.3a | 16.7a | 5.8b | 16.2a |
| CV (%) | 2.2 | 8.5 | 8.8 | 7.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Colombari, L.F.; Chamma, L.; da Silva, G.F.; Zanetti, W.A.L.; Putti, F.F.; Cardoso, A.I.I. Maturation and Post-Harvest Resting of Fruits Affect the Macronutrients and Protein Content in Sweet Pepper Seeds. Plants 2022, 11, 2084. https://doi.org/10.3390/plants11162084
Colombari LF, Chamma L, da Silva GF, Zanetti WAL, Putti FF, Cardoso AII. Maturation and Post-Harvest Resting of Fruits Affect the Macronutrients and Protein Content in Sweet Pepper Seeds. Plants. 2022; 11(16):2084. https://doi.org/10.3390/plants11162084
Chicago/Turabian StyleColombari, Lidiane Fernandes, Larissa Chamma, Gustavo Ferreira da Silva, Willian Aparecido Leoti Zanetti, Fernando Ferrari Putti, and Antonio Ismael Inácio Cardoso. 2022. "Maturation and Post-Harvest Resting of Fruits Affect the Macronutrients and Protein Content in Sweet Pepper Seeds" Plants 11, no. 16: 2084. https://doi.org/10.3390/plants11162084