
Cyanobacteria-Mediated Immune Responses in Pepper Plants against Fusarium Wilt

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Source of Pathogen
2.2. Growth Conditions of Cyanobacteria
2.3. Preparation of Cyanobacteria Extracts
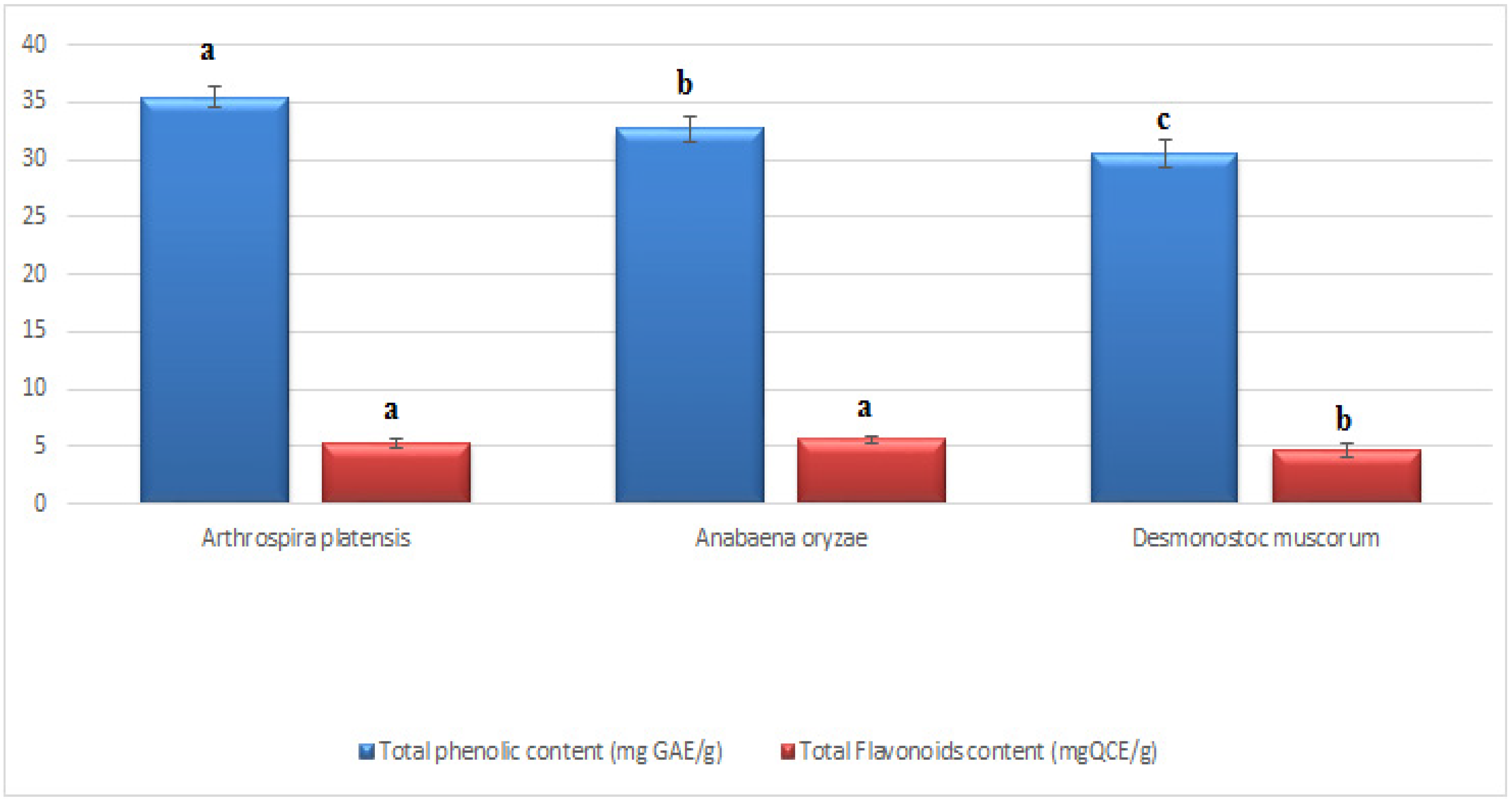
2.4. Total Phenolic Content (TPC) of Tested Cyanobacteria
2.5. Total Flavonoids Content (T.F.C.s) of Tested Cyanobacteria
2.6. Experimental Design
2.7. Disease Symptoms and Disease Index
2.8. Resistance Indicators in Pepper Plant
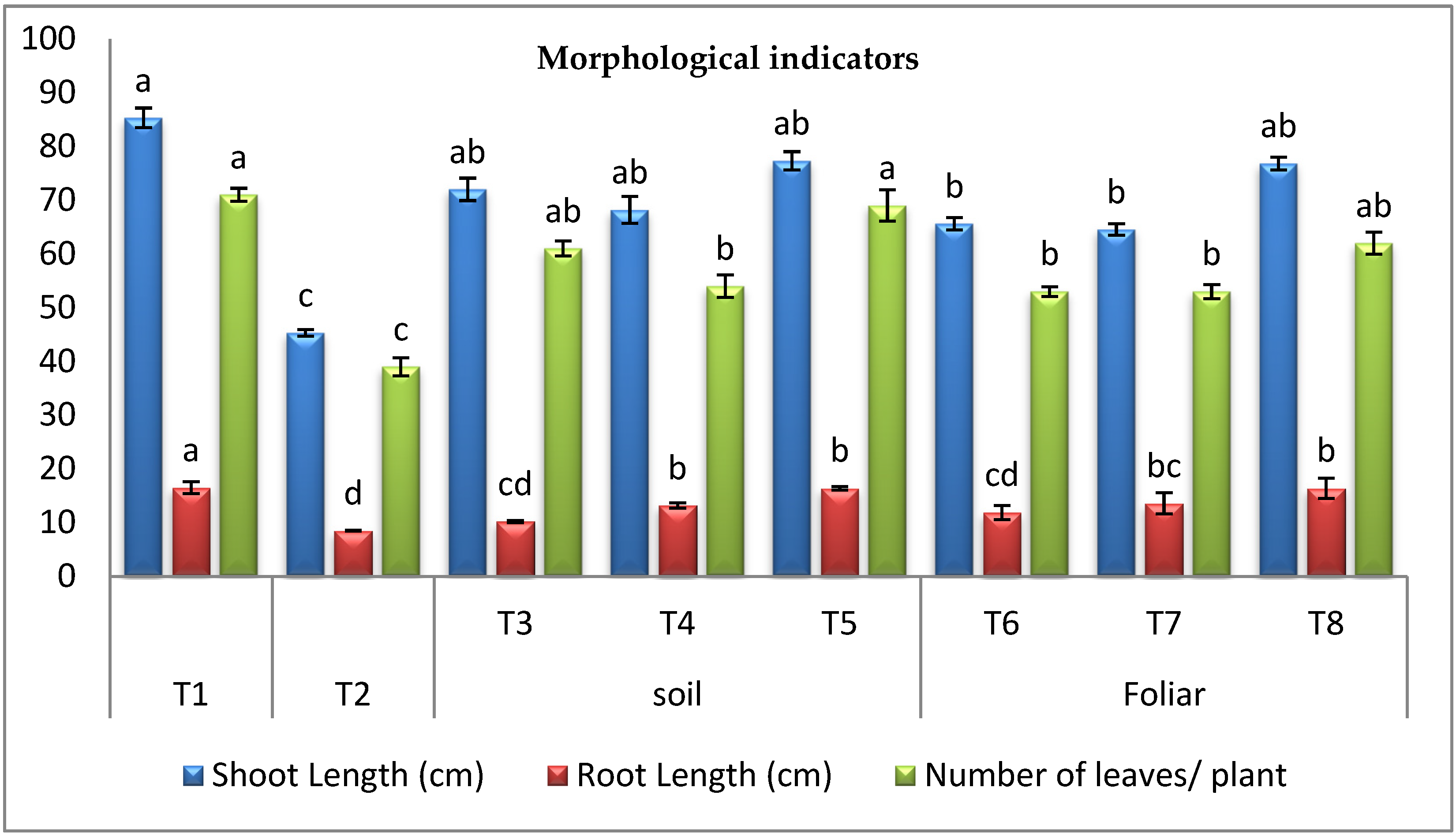
2.8.1. Morphological Resistance Indicators
2.8.2. Photosynthetic Pigment Determination
2.8.3. Estimation of Osmolytes Content
Soluble Sugar Determination
Soluble Protein Estimation
Proline Content Determination
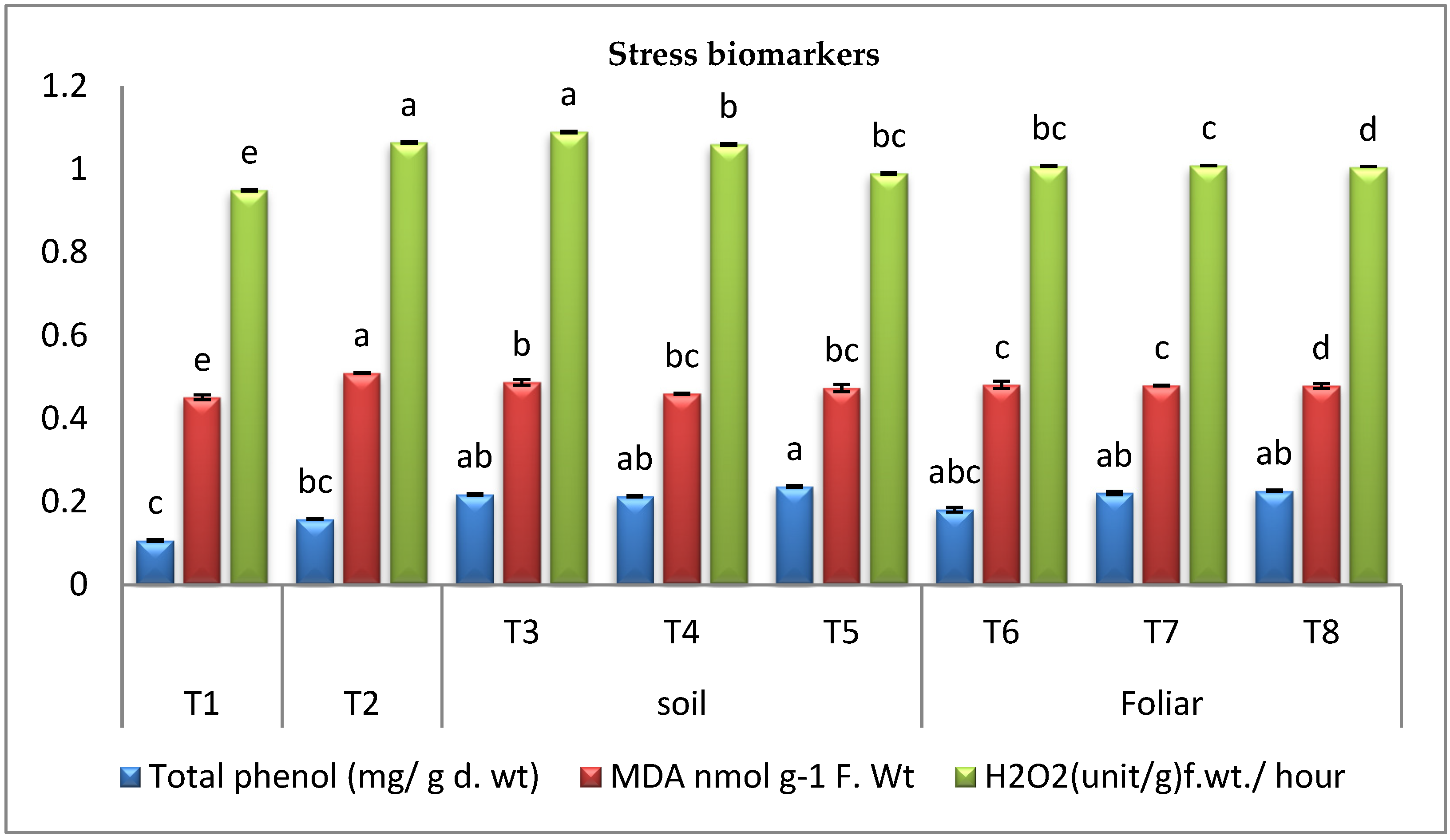
Total Phenol
2.8.4. Estimation of Malondialdehyde (MDA) and Hydrogen Peroxide (H2O2) Contents
2.8.5. Antioxidant Enzymes Activities Assay
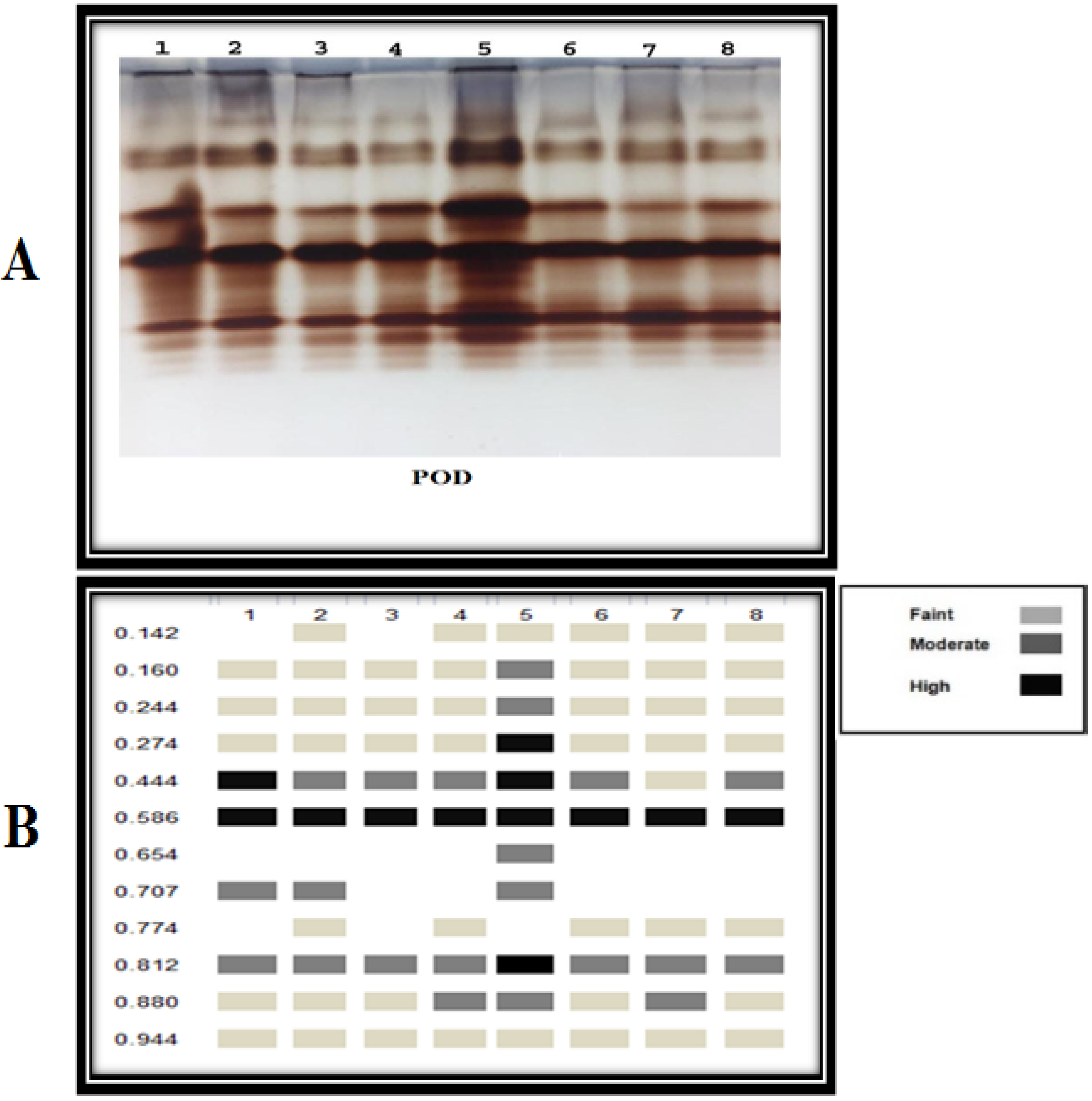
2.8.6. Isozyme Electrophoresis
2.8.7. Endogenous Hormones (IAA, SA, and ABA) Contents
2.9. Statistical Analysis
3. Results
3.1. Total Phenolics and Flavonoids of the Applied Cyanobacteria
3.2. Effect of Cyanobacteria on Disease Index of Infected Pepper Plants
3.3. Resistance Indicators in Pepper Plant
3.3.1. Morphological Indicators
3.3.2. Photosynthetic Pigments
3.3.3. Osmolytes (Soluble Sugar, Soluble Protein, and Proline) Contents
3.3.4. Stress Biomarkers
3.3.5. Oxidative Enzymes Activity
3.3.6. Antioxidant Isozymes
3.3.7. Endogenous Hormones
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tijjani, A.; Khairulmazmi, A. Global Food Demand and the Roles of Microbial Communities in Sustainable Crop Protection and Food Security: An Overview. Role Microb. Communities Sustain. 2021, 29, 81–107. [Google Scholar]
- El-Feky, N.; Essa, T.; Elzaawely, A.A.; El-Zahaby, H.M. Antagonistic activity of some bioagents against root rot diseases of pepper (Capsicum annum L.). Environ. Biodivers. Soil Secur. 2019, 3, 103–104. [Google Scholar] [CrossRef]
- Abdelaziz, A.M.; Dacrory, S.; Hashem, A.H.; Attia, M.S.; Hasanin, M.; Fouda, H.M.; Kamel, S.; ElSaied, H. Protective role of zinc oxide nanoparticles based hydrogel against wilt disease of pepper plant. Biocatal. Agric. Biotechnol. 2021, 35, 102083. [Google Scholar] [CrossRef]
- Rani, L.; Thapa, K.; Kanojia, N.; Sharma, N.; Singh, S.; Grewal, A.S.; Srivastav, A.L.; Kaushal, J. An extensive review on the consequences of chemical pesticides on human health and environment. J. Clean. Prod. 2020, 283, 124657. [Google Scholar] [CrossRef]
- Bhandari, G. An Overview of Agrochemicals and Their Effects on Environment in Nepal. Appl. Ecol. Environ. Sci. 2014, 2, 66–73. [Google Scholar] [CrossRef] [Green Version]
- Alhakim, A.A.; Hashem, A.; Abdelaziz, A.M.; Attia, M.S. Impact of plant growth promoting fungi on biochemical defense performance of Tomato under Fusarial infection. Egypt. J. Chem. 2022. [Google Scholar] [CrossRef]
- Aldinary, A.M.; Abdelaziz, A.M.; Farrag, A.A.; Attia, M.S. Biocontrol of tomato Fusarium wilt disease by a new Moringa endophytic Aspergillus isolates. Mater. Today Proc. 2021. [Google Scholar] [CrossRef]
- Attia, M.S.; El-Wakil, D.A.; Hashem, A.H.; Abdelaziz, A.M. Antagonistic Effect of Plant Growth-Promoting Fungi Against Fusarium Wilt Disease in Tomato: In vitro and In vivo Study. Appl. Biochem. Biotechnol. 2022, 1–19. Available online: https://link.springer.com/content/pdf/10.1007/s12010-022-03975-9.pdf (accessed on 3 March 2022). [CrossRef]
- Rausher, M.D. Co-evolution and plant resistance to natural enemies. Nature 2001, 411, 857–864. [Google Scholar] [CrossRef]
- Attia, M.S.; El-Sayyad, G.S.; Elkodous, M.A.; Khalil, W.F.; Nofel, M.M.; Abdelaziz, A.M.; Farghali, A.A.; El-Batal, A.I.; El Rouby, W.M. Chitosan and EDTA conjugated graphene oxide antinematodes in Eggplant: Toward improving plant immune response. Int. J. Biol. Macromol. 2021, 179, 333–344. [Google Scholar] [CrossRef] [PubMed]
- Witzell, J.; Martín, J.A. Phenolic metabolites in the resistance of northern forest trees to pathogens—Past experiences and future prospects. Can. J. For. Res. 2008, 38, 2711–2727. [Google Scholar] [CrossRef]
- Vargas-Hernandez, M.; Macias-Bobadilla, I.; Guevara-Gonzalez, R.; Rico-Garcia, E.; Ocampo-Velazquez, R.; Avila-Juarez, L.; Torres-Pacheco, I. Nanoparticles as Potential Antivirals in Agriculture. Agriculture 2020, 10, 444. [Google Scholar] [CrossRef]
- Ab Rahman, S.F.S.; Singh, E.; Pieterse, C.M.; Schenk, P.M. Emerging microbial biocontrol strategies for plant pathogens. Plant Sci. 2018, 267, 102–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Kim, J.-D. Inhibitory effect of algal extracts on mycelial growth of the tomato-wilt pathogen, Fusarium oxysporum f. sp. lycopersici. Mycobiology 2008, 36, 242–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.-M.; Ryu, C.-M. Algae as New Kids in the Beneficial Plant Microbiome. Front. Plant Sci. 2021, 12, 599742. [Google Scholar] [CrossRef]
- Refaay, D.A.; El-Marzoki, E.M.; Abdel-Hamid, M.I.; Haroun, S.A. Effect of foliar application with Chlorella vulgaris, Tetradesmus dimorphus, and Arthrospira platensis as biostimulants for common bean. J. Appl. Phycol. 2021, 33, 3807–3815. [Google Scholar] [CrossRef]
- Swain, D.L.; Singh, D.; Horton, D.E.; Mankin, J.S.; Ballard, T.C.; Diffenbaugh, N.S. Remote Linkages to Anomalous Winter Atmospheric Ridging Over the Northeastern Pacific. J. Geophys. Res. Atmos. 2017, 12, 194–209. [Google Scholar] [CrossRef]
- Abdel-Hafez, S.I.I.; Abo-Elyousr, K.A.M.; Abdel-Rahim, I.R. Fungicidal activity of extracellular products of cyanobacteria against Alternaria porri. Eur. J. Phycol. 2015, 50, 239–245. [Google Scholar] [CrossRef]
- Morales-Jiménez, M.; Gouveia, L.; Yáñez-Fernández, J.; Castro-Muñoz, R.; Barragán-Huerta, B.E. Production, Preparation and Characterization of Microalgae-Based Biopolymer as a Potential Bioactive Film. Coatings 2020, 10, 120. [Google Scholar] [CrossRef] [Green Version]
- Hlima, H.B.; Bohli, T.; Kraiem, M.; Ouederni, A.; Mellouli, L.; Michaud, P.; Abdelkafi, S.; Smaoui, S. Combined effect of Spirulina platensis and Punica granatum peel extacts: Phytochemical content and antiphytophatogenic activity. Appl. Sci. 2019, 9, 5475. [Google Scholar] [CrossRef] [Green Version]
- Imara, D.A.; Zaky, W.H.; Ghebrial, E.W. Performance of Soil Type, Cyanobacterium Spirulina platensis and Biofertilizers on Controlling Damping-off, Root Rot and Wilt Diseases of Moringa (Moringa oleifera Lam.) in Egypt. Egypt. J. Phytopathol. 2021, 49, 10–28. [Google Scholar] [CrossRef]
- Souza, M.M.D.; Prietto, L.; Ribeiro, A.C.; Souza, T.D.D.; Badiale-Furlong, E. Assessment of the antifungal activity of Spirulina platensis phenolic extract against Aspergillus flavus. Ciência e Agrotecnologia 2011, 35, 1050–1058. [Google Scholar] [CrossRef] [Green Version]
- El-Mougy, N.S.; Abdel-Kader, M.M. Effect of Commercial Cyanobacteria Products on the Growth and Antagonistic Ability of Some Bioagents under Laboratory Conditions. J. Pathog. 2013, 2013, 838329. [Google Scholar] [CrossRef] [PubMed]
- Afify, A.; Ashour, A. Use of Cyanobacteria for Controlling Flax Seedling Blight. J. Agric. Chem. Biotechnol. 2018, 9, 259–261. [Google Scholar] [CrossRef]
- Prasanna, R.; Chaudhary, V.; Gupta, V.; Babu, S.; Kumar, A.; Singh, R.; Shivay, Y.S.; Nain, L. Cyanobacteria mediated plant growth promotion and bioprotection against Fusarium wilt in tomato. Eur. J. Plant Pathol. 2013, 136, 337–353. [Google Scholar] [CrossRef]
- Abdelaziz, A.M.; El-Wakil, D.A.; Attia, M.S.; Ali, O.M.; AbdElgawad, H.; Hashem, A.H. Inhibition of Aspergillus flavus Growth and Aflatoxin Production in Zea mays L. Using Endophytic Aspergillus fumigatus. J. Fungi 2022, 8, 482. [Google Scholar] [CrossRef] [PubMed]
- Dutta, A.; Mandal, A.; Kundu, A.; Malik, M.; Chaudhary, A.; Khan, M.R.; Shanmugam, V.; Rao, U.; Saha, S.; Patanjali, N.; et al. Deciphering the Behavioral Response of Meloidogyne incognita and Fusarium oxysporum toward Mustard Essential Oil. Front. Plant Sci. 2021, 12, 1791. [Google Scholar] [CrossRef]
- Harun, R.; Yip, J.W.; Thiruvenkadam, S.; Ghani, W.A.; Cherrington, T.; Danquah, M.K. Algal biomass conversion to bioethanol–a step-by-step assessment. Biotechnol. J. 2014, 9, 73–86. [Google Scholar] [CrossRef]
- Singh, D.P.; Prabha, R.; Verma, S.; Meena, K.K.; Yandigeri, M. Antioxidant properties and polyphenolic content in terrestrial cyanobacteria. 3 Biotech 2017, 7, 134. [Google Scholar] [CrossRef] [Green Version]
- Prasanna, R.; Bidyarani, N.; Babu, S.; Hossain, F.; Shivay, Y.S.; Nain, L. Cyanobacterial inoculation elicits plant defense response and enhanced Zn mobilization in maize hybrids. Cogent Food Agric. 2015, 1, 998507. [Google Scholar] [CrossRef]
- Senousy, H.H.; El-Sheekh, M.M.; Saber, A.A.; Khairy, H.M.; Said, H.A.; Alhoqail, W.A.; Abu-Elsaoud, A.M. Biochemical Analyses of Ten Cyanobacterial and Microalgal Strains Isolated from Egyptian Habitats, and Screening for Their Potential against Some Selected Phytopathogenic Fungal Strains. Agronomy 2022, 12, 1340. [Google Scholar] [CrossRef]
- Senousy, H.H.; Ellatif, S.A.; Ali, S. Assessment of the antioxidant and anticancer potential of different isolated strains of cyanobacteria and microalgae from soil and agriculture drain water. Environ. Sci. Pollut. Res. 2020, 27, 18463–18474. [Google Scholar] [CrossRef]
- Rippka, R.; Deruelles, J.; Waterbury, J.B.; Herdman, M.; Stanier, R.Y. Generic Assignments, Strain Histories and Properties of Pure Cultures of Cyanobacteria. Microbiology 1979, 111, 1–61. [Google Scholar] [CrossRef] [Green Version]
- Zarrouk, C. Contribution a l’etude d’une Cyanophycee. Influence de Divers Facteurs Physiques et Chimiques sur la Croissance et la Photosynthese de Spirulina mixima. Ph.D. Thesis, University of Paris, Paris, France, 1966. [Google Scholar]
- Bello, A.S.; Ben-Hamadou, R.; Hamdi, H.; Saadaoui, I.; Ahmed, T. Application of Cyanobacteria (Roholtiella sp.) Liquid Extract for the Alleviation of Salt Stress in Bell Pepper (Capsicum annuum L.) Plants Grown in a Soilless System. Plants 2021, 11, 104. [Google Scholar] [CrossRef]
- Jain, S.; Jain, A.; Jain, S.; Malviya, N.; Jain, V.; Kumar, D. Estimation of total phenolic, tannins, and flavonoid contents and antioxidant activity of Cedrus deodara heart wood extracts. Egypt. Pharm. J. 2015, 14, 10. [Google Scholar] [CrossRef]
- Chang, C.-C.; Yang, M.H.; Wen, H.M.; Chern, J. Estimation of total flavonoid content in propolis by two complementary colorimetric methods. J. Food Drug Anal. 2002, 10, 3. [Google Scholar]
- Farrag, A.; Attia, M.S.; Younis, A.; Abd Elaziz, A.M.A. Potential impacts of elicitors to improve tomato plant disease resistance. Azhar. Bull. Sci. 2017, 9, 311–321. [Google Scholar]
- Abdelaziz, A.M.; Salem, S.S.; Khalil, A.M.A.; El-Wakil, D.A.; Fouda, H.M.; Hashem, A.H. Potential of biosynthesized zinc oxide nanoparticles to control Fusarium wilt disease in eggplant (Solanum melongena) and promote plant growth. BioMetals 2022, 35, 601–616. [Google Scholar] [CrossRef]
- The Chlorophylls. 1966. Available online: https://www.elsevier.com/books/the-chlorophylls/vernon/978-1-4832-3289-8 (accessed on 12 March 2022).
- Badawy, A.; Alotaibi, M.; Abdelaziz, A.; Osman, M.; Khalil, A.; Saleh, A.; Mohammed, A.; Hashem, A. Enhancement of Seawater Stress Tolerance in Barley by the Endophytic Fungus Aspergillus ochraceus. Metabolites 2021, 11, 428. [Google Scholar] [CrossRef]
- Umbreit, W.W.; Burris, R.H.; Stauffer, J.F. Manometric Techniques: A Manual Describing Methods Applicable to the Study of Tissue Metabolism. Available online: https://www.car.chula.ac.th/display7.php?bib=b1247738 (accessed on 12 March 2022).
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Irigoyen, J.J.; Einerich, D.W.; Sanchez-Diaz, M. Water stress induced changes in concentrations of proline and total soluble sugars in nodulated alfalfa (Medicago sativa) plants. Physiol. Plant. 1992, 84, 55–60. [Google Scholar] [CrossRef]
- Dai, G.; Andary, C.; Cosson-Mondolot, L.; Boubals, D. Polyphenols and Resistance of Grapevines to Downy Mildew. Acta Hortic. 1993, 381, 763–766. [Google Scholar] [CrossRef]
- Mukherjee, S.P.; Choudhuri, M.A. Implications of water stress-induced changes in the levels of endogenous ascorbic acid and hydrogen peroxide in Vigna seedlings. Physiol. Plant. 1983, 58, 166–170. [Google Scholar] [CrossRef]
- Knegt, E.; Bruinsma, J. A rapid, sensitive and accurate determination of indolyl-3-acetic acid. Phytochemistry 1973, 12, 753–756. [Google Scholar] [CrossRef]
- Snedecor, G.W.; Cochran, W.G. Statistical Methods, 7th ed.; Iowa State University USA: Ames, IA, USA, 1980; pp. 80–86. [Google Scholar]
- Righini, H.; Roberti, R. Algae and Cyanobacteria as Biocontrol Agents of Fungal Plant Pathogens. In Plant Microbe Interface; Springer: Cham, Switzerland, 2019; pp. 219–238. [Google Scholar]
- Usharani, G.; Srinivasan, G.; Sivasakthi, S.; Saranraj, P. Antimicrobial activity of Spirulina platensis solvent extracts against pathogenic bacteria and fungi. Adv. Biol. Res. 2015, 9, 292–298. [Google Scholar]
- Kulik, M.M. The potential for using cyanobacteria (blue-green algae) and algae in the biological control of plant pathogenic bacteria and fungi. Eur. J. Plant Pathol. 1995, 101, 585–599. [Google Scholar] [CrossRef]
- Bai, G.; Shaner, G. Management and resistance in wheat and barley to Fusarium head blight. Annu. Rev. Phytopathol. 2004, 42, 135–161. [Google Scholar] [CrossRef]
- Gunupuru, L.R.; Patel, J.S.; Sumarah, M.W.; Renaud, J.B.; Mantin, E.G.; Prithiviraj, B. A plant biostimulant made from the marine brown algae Ascophyllum nodosum and chitosan reduce Fusarium head blight and mycotoxin contamination in wheat. PLoS ONE 2019, 14, e0220562. [Google Scholar] [CrossRef]
- Ahmed, A.F.; Attia, M.S.; Faramawy, F.; Salaheldin, M.M. Impact of PGPM in Pepper Immune-Response to Fusarium Wilt Disease. Saudi J. Pathol. Microbiol. (SJPM) 2018, 3, 5–15. [Google Scholar]
- Kaul, S.; Choudhary, M.; Gupta, S.; Dhar, M.K. Engineering Host Microbiome for Crop Improvement and Sustainable Agriculture. Front. Microbiol. 2021, 12, 635917. [Google Scholar] [CrossRef]
- Tan, C.-Y.; Dodd, I.C.; Chen, J.E.; Phang, S.-M.; Chin, C.F.; Yow, Y.-Y.; Ratnayeke, S. Regulation of algal and cyanobacterial auxin production, physiology, and application in agriculture: An overview. J. Appl. Phycol. 2021, 33, 2995–3023. [Google Scholar] [CrossRef]
- Wang, L.Y.; Liu, J.L.; Wang, W.X.; Sun, Y. Exogenous melatonin improves growth and photosynthetic capacity of cucumber under salinity-induced stress. Photosynthetica 2016, 54, 19–27. [Google Scholar] [CrossRef]
- He, Y.; Shi, Y.; Zhang, X.; Xu, X.; Wang, H.; Li, L.; Zhang, Z.; Shang, H.; Wang, Z.; Wu, J.-L. The OsABCI7 Transporter Interacts with OsHCF222 to Stabilize the Thylakoid Membrane in Rice. Plant Physiol. 2020, 184, 283–299. [Google Scholar] [CrossRef]
- Grzesik, M.; Romanowska-Duda, Z.; Kalaji, H. Effectiveness of cyanobacteria and green algae in enhancing the photosynthetic performance and growth of willow (Salix viminalis L.) plants under limited synthetic fertilizers application. Photosynthetica 2017, 55, 510–521. [Google Scholar] [CrossRef]
- Kunui, K.; Singh, S.S. Protective role of antioxidative enzymes and antioxidants against iron-induced oxidative stress in the cyanobacterium Anabaena sphaerica isolated from iron rich paddy field of Chhattisgarh, India. Indian J. Exp. Biol. (IJEB) 2020, 58. [Google Scholar]
- Foyer, C.H.; Shigeoka, S. Understanding Oxidative Stress and Antioxidant Functions to Enhance Photosynthesis. Plant Physiol. 2010, 155, 93–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Attia, M.S.; Younis, A.M.; Ahmed, A.F.; Elaziz, A.M.A. Comprehensive Management for Wilt Disease Caused by Fusarium Oxysporum in Tomato Plant. Int. J. Innov. Sci. Eng. Technol. 2016, 4, 2348–7968. [Google Scholar]
- Singh, S. A review on possible elicitor molecules of cyanobacteria: Their role in improving plant growth and providing tolerance against biotic or abiotic stress. J. Appl. Microbiol. 2014, 117, 1221–1244. [Google Scholar] [CrossRef] [PubMed]
- Attia, M.S.; El-Naggar, H.A.; Abdel-Daim, M.M.; El-Sayyad, G.S. The potential impact of Octopus cyanea extracts to improve eggplant resistance against Fusarium-wilt disease: In vivo and in vitro studies. Environ. Sci. Pollut. Res. 2021, 28, 35854–35869. [Google Scholar] [CrossRef] [PubMed]
- Anaraki, Z.E.; Tafreshi, S.A.H.; Shariati, M. Transient silencing of heat shock proteins showed remarkable roles for HSP70 during adaptation to stress in plants. Environ. Exp. Bot. 2018, 155, 142–157. [Google Scholar] [CrossRef]
- Singh, D.P.; Prabha, R.; Yandigeri, M.S.; Arora, D.K. Cyanobacteria-mediated phenylpropanoids and phytohormones in rice (Oryza sativa) enhance plant growth and stress tolerance. Antonie Leeuwenhoek 2011, 100, 557–568. [Google Scholar] [CrossRef]
- Nowruzi, B.; Bouaïcha, N.; Metcalf, J.S.; Porzani, S.J.; Konur, O. Plant-cyanobacteria interactions: Beneficial and harmful effects of cyanobacterial bioactive compounds on soil-plant systems and subsequent risk to animal and human health. Phytochemistry 2021, 192, 112959. [Google Scholar] [CrossRef] [PubMed]
- Hashem, A.; Tabassum, B.; Fathi Abd Allah, E. Bacillus subtilis: A plant-growth promoting rhizobacterium that also impacts biotic stress. Saudi J. Biol. Sci. 2019, 26, 1291–1297. [Google Scholar] [CrossRef] [PubMed]
- Hashem, A.; Abdelaziz, A.; Askar, A.; Fouda, H.; Khalil, A.; Abd-Elsalam, K.; Khaleil, M. Bacillus megaterium-Mediated Synthesis of Selenium Nanoparticles and Their Antifungal Activity against Rhizoctonia solani in Faba Bean Plants. J. Fungi 2021, 7, 195. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive Oxygen Species, Oxidative Damage, and Antioxidative Defense Mechanism in Plants under Stressful Conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef] [Green Version]
- Jayamohan, N.S.; Patil, S.V.; Kumudini, B.S. Reactive oxygen species (ROS) and antioxidative enzyme status in Solanum lycopersicum on priming with fluorescent Pseudomonas spp. against Fusarium oxysporum. Biologia 2018, 73, 1073–1082. [Google Scholar] [CrossRef]
- Bari, R.; Jones, J.D.G. Role of plant hormones in plant defence responses. Plant Mol. Biol. 2008, 69, 473–488. [Google Scholar] [CrossRef]
- Arif, Y.; Sami, F.; Siddiqui, H.; Bajguz, A.; Hayat, S. Salicylic acid in relation to other phytohormones in plant: A study towards physiology and signal transduction under challenging environment. Environ. Exp. Bot. 2020, 175, 104040. [Google Scholar] [CrossRef]
- Ashraf, M.; Akram, N.A.; Arteca, R.N.; Foolad, M.R. The Physiological, Biochemical and Molecular Roles of Brassinosteroids and Salicylic Acid in Plant Processes and Salt Tolerance. Crit. Rev. Plant Sci. 2010, 29, 162–190. [Google Scholar] [CrossRef]
- Bayona-Morcillo, P.J.; Plaza, B.M.; Gómez-Serrano, C.; Rojas, E.; Jiménez-Becker, S. Effect of the foliar application of cyanobacterial hydrolysate (Arthrospira platensis) on the growth of Petunia x hybrida under salinity conditions. J. Appl. Phycol. 2020, 32, 4003–4011. [Google Scholar] [CrossRef]
- Attia, M.S.; El-Sayyad, G.S.; Abd Elkodous, M.; El-Batal, A.I. The effective antagonistic potential of plant growth-promoting rhizobacteria against Alternaria solani-causing early blight disease in tomato plant. Sci. Hortic. 2020, 266, 109289. [Google Scholar] [CrossRef]
- Kumar, G.; Teli, B.; Mukherjee, A.; Bajpai, R.; Sarma, B.K. Secondary Metabolites from Cyanobacteria: A Potential Source for Plant Growth Promotion and Disease Management. In Secondary Metabolites of Plant Growth Promoting Rhizomicroorganisms; Springer: Singapore, 2019; pp. 239–252. [Google Scholar]
- Mauch-Mani, B.; Mauch, F. The role of abscisic acid in plant–pathogen interactions. Curr. Opin. Plant Biol. 2005, 8, 409–414. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, K.; Pflugmacher, M.; Klages, S.; Mäser, A.; Mock, A.; Stahl, D.J. Accumulation of the hormone abscisic acid (ABA) at the infection site of the fungus Cercospora beticola supports the role of ABA as a repressor of plant defence in sugar beet. Mol. Plant Pathol. 2008, 9, 661–673. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Method of Application | Disease Symptoms Classes | DI (Disease Index) (%) | Protection (%) | ||||
|---|---|---|---|---|---|---|---|---|
| 0 | 1 | 2 | 3 | 4 | ||||
| D. muscorum | Through soil | 2 | 1 | 2 | 0 | 1 | 37.5 | 54.99 |
| A. oryzae | 1 | 3 | 1 | 1 | 0 | 33.33 | 60.03 | |
| A. platensis | 2 | 1 | 3 | 0 | 0 | 29.16 | 65 | |
| D. muscorum | Through Foliar | 5 | 0 | 1 | 0 | 0 | 8.3 | 90 |
| A. oryzae | 3 | 0 | 3 | 0 | 0 | 25 | 69.9 | |
| A. platensis | 5 | 1 | 0 | 0 | 0 | 4.16 | 95 | |
| Control infected | 0 | 0 | 1 | 2 | 3 | 83.33 | 0 | |
| Treatments | Method of Application | Total Carbohydrate | Total Protein | Free Proline |
|---|---|---|---|---|
| Healthy control | 30 ± 2.4 a | 34.2 ± 0.2 a | 0.76 ± 0.002 d | |
| Infected control | 18.45 ± 1.62 c | 14.66 ± 1.6 f | 0.85 ± 0.001 e | |
| D. muscorum | Through soil | 19.55 ± 0.82 c | 29.08 ± 0.4 b | 0.913 ± 0.001 a |
| A. oryzae | 27.7 ± 0.9 a | 22.26 ± 1 e | 0.86± 0.001 b | |
| A. platensis | 24.02 ± 2.62 b | 29.56 ±0.3 b | 0.91 ± 0.002 a | |
| D. muscorum | Through Foliar | 19.74 ± 1.4 c | 26.46 ± 0.9 c | 0.862± 0.001 c |
| A. oryzae | 20.7 ± 1.7 c | 24.8 ± 0.7 d | 0.863 ± 0.001 c | |
| A. platensis | 21.38 ± 1.26 bc | 28.56 ± 0.7 b | 0.864 ± 0.001 bc | |
| LSD at 0.05 | 2.967 | 1.507 | 0.0029 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdelaziz, A.M.; Attia, M.S.; Salem, M.S.; Refaay, D.A.; Alhoqail, W.A.; Senousy, H.H. Cyanobacteria-Mediated Immune Responses in Pepper Plants against Fusarium Wilt. Plants 2022, 11, 2049. https://doi.org/10.3390/plants11152049
Abdelaziz AM, Attia MS, Salem MS, Refaay DA, Alhoqail WA, Senousy HH. Cyanobacteria-Mediated Immune Responses in Pepper Plants against Fusarium Wilt. Plants. 2022; 11(15):2049. https://doi.org/10.3390/plants11152049
Chicago/Turabian StyleAbdelaziz, Amer Morsy, Mohamed S. Attia, Marwa S. Salem, Dina A. Refaay, Wardah A. Alhoqail, and Hoda H. Senousy. 2022. "Cyanobacteria-Mediated Immune Responses in Pepper Plants against Fusarium Wilt" Plants 11, no. 15: 2049. https://doi.org/10.3390/plants11152049