
ABA and Not Chilling Reduces Heat Requirement to Force Cherry Blossom after Endodormancy Release


Abstract
:1. Introduction
- (1)
- Pure forcing or 1-phase models, e.g., thermal time or spring warming models ([12] and the references therein), only consider the heat accumulation during the ontogenetic phase in spring. The advantage of these models is that they do not have to take the dormancy phases into account. However, they should only be used if endodormancy is assuredly released before heat accumulation starts. A challenge of these models is the precise definition of the starting date of forcing accumulation, according to our notation referred as t1*, which cannot be observed on the tree and is thus mostly an optimized model parameter and not physiologically based.
- (2)
- Chilling/forcing or 2-phase models, which can be realized as sequential or parallel models, try to consider both the dormancy and growth phase in order to calculate the timing of phenological events ([12] and the references therein). This approach is justified, because air temperature and/or photoperiod are considered to be the driving factors for the induction, maintenance and release of dormancy. Shortening daylength and/or low temperature signals are responsible for initiation and development of endodormancy in autumn [13]. However, after the induction of endodormancy, air temperature is the most important factor to leave this state in favor of resumption of growth potential [9]. Thus, phenological models assume that a sufficient amount of chill temperatures are needed for endodormancy release, which is highly variable between tree species, provenances and cultivars [14,15]. Afterwards, by elevated air temperatures in spring, the subsequent quiescent phase of ecodormancy is overcome and results in the beginning of ontogenetic development. However, even for this category of models, determining the exact date of endodormancy release (t1) and the beginning of ontogenetic development (t1*) is challenging, because these parameters are predominantly statistically derived. Sequential phenology models assume that forcing temperatures are only effective if a certain chill requirement of the plant is met, i.e., endodormancy is released [16]. In parallel models [17,18,19,20], chill and heat are accumulated simultaneously with the assumption that a lack of chilling can be substituted by a higher forcing amount and vice versa (Equation (1)). In these models, chilling is usually calculated from autumn of the previous year (September or November) and forcing from January or February, both until the onset date of the phenological spring event [17,21,22,23].
2. Results
2.1. Average Timing and Duration of Dormancy Phases and Ontogenetic Development for ‘Summit’ Flower Buds
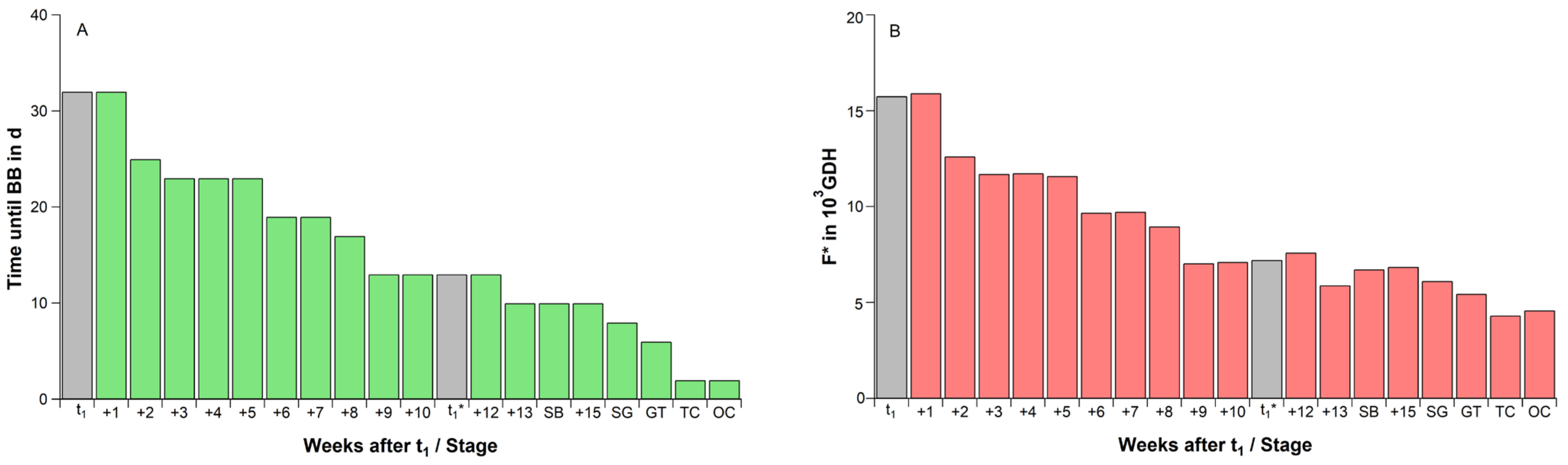
2.2. Average Chill and Heat Accumulation of ‘Summit’ Flower Buds during Dormancy Phases and Ontogenetic Development
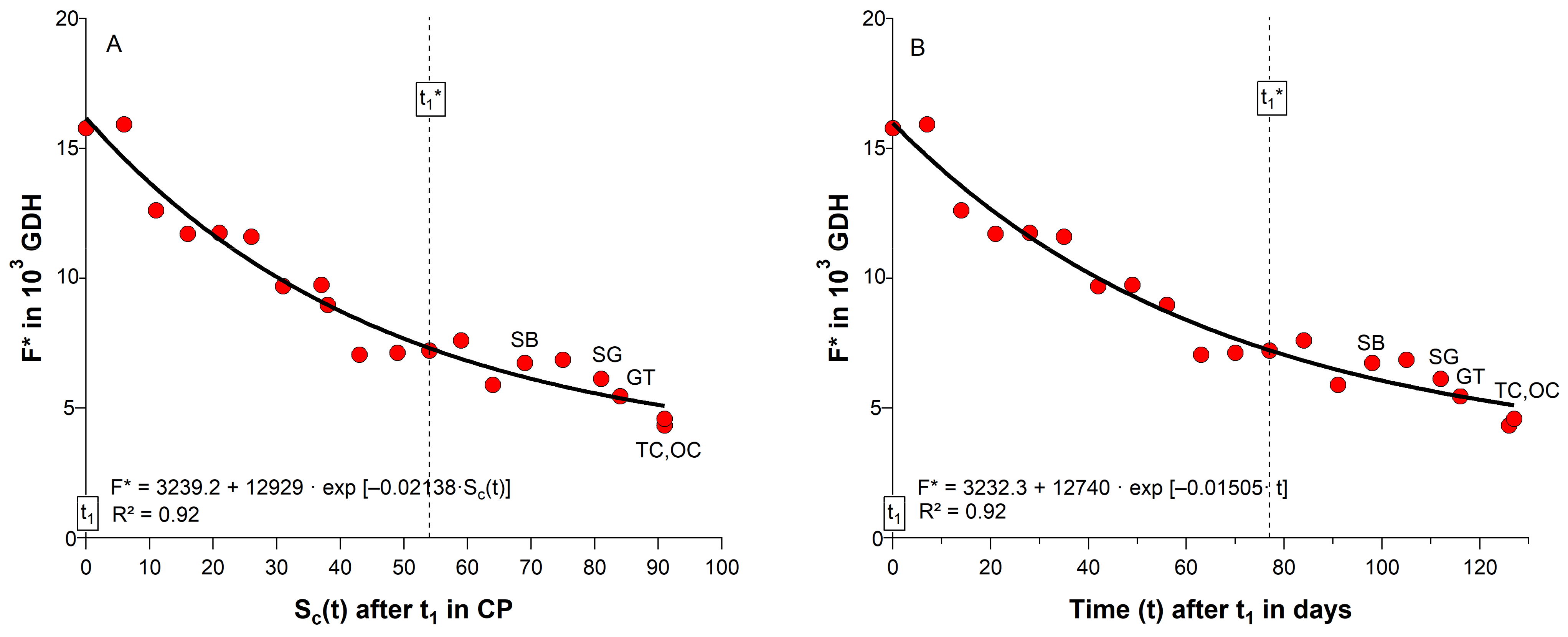
2.3. Substitution of Chilling and Forcing under Controlled Conditions
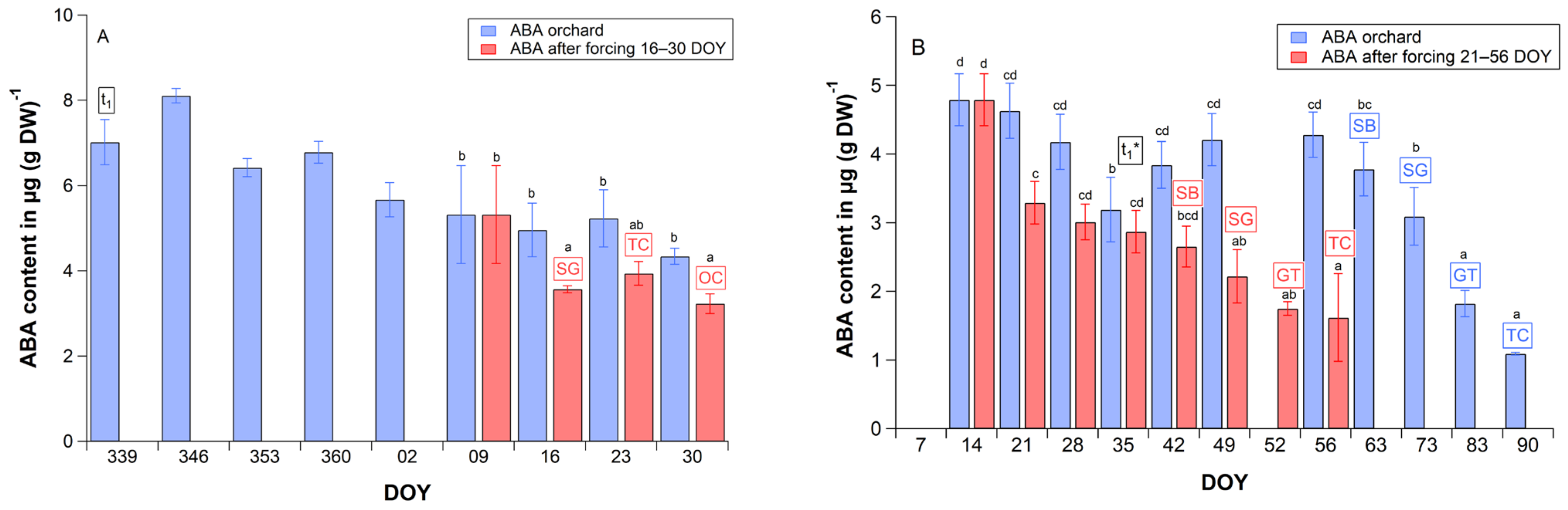
2.4. Abscisic Acid Content in Cherry Flower Buds
3. Discussion
3.1. Timing and Duration of Ecodormancy Phase
3.2. Chill and Forcing Compensation
3.3. ABA Content in Cherry Flower Buds
3.4. Physiological Function of ABA
4. Material and Methods
4.1. Experimental Site
4.2. Phenological Observation in the Orchard
4.3. Determination of Ecodormancy Phase (t1–t1*)
4.4. Chilling and Forcing Demand under Controlled Conditions
4.5. Targeted Analysis of the Abscisic Acid (ABA) Content in Flower Buds
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Samish, R.M. Dormancy in woody plants. Annu. Rev. Plant Physiol. 1954, 5, 183–204. [Google Scholar] [CrossRef]
- Cooke, J.E.K.; Eriksson, M.E.; Junttila, O. The dynamic nature of bud dormancy in trees: Environmental control and molecular mechanisms. Plant Cell Environ. 2012, 35, 1707–1728. [Google Scholar] [CrossRef] [PubMed]
- Rinne, P.L.H.; Kaikuranta, P.M.; van der Schoot, C. The shoot apical meristem restores its symplasmic organization during chilling-induced release from dormancy. Plant J. 2001, 26, 249–264. [Google Scholar] [CrossRef] [PubMed]
- Fadón, E.; Fernandez, E.; Behn, H.; Fadón, E.; Rodrigo, J.; Herrero, M. Is there a specific stage to rest? Morphological changes in flower primordia in relation to endodormancy in sweet cherry (Prunus avium L.). Trees Struct. Funct. 2018, 32, 1583–1594. [Google Scholar] [CrossRef]
- Lang, G.A.; Early, J.D.; Martin, G.C.; Darnell, R.L. Endodormancy, paradormancy, and ecodormancy—Physiological terminology and classification for dormancy research. Hortscience 1987, 22, 371–377. [Google Scholar]
- Singh, R.K.; Svystun, T.; AlDahmash, B.; Jönsson, A.M.; Bhalerao, R.P. Photoperiod- and temperature-mediated control of phenology in trees—A molecular perspective. New Phytol. 2017, 213, 511–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heide, O.M. Interaction of photoperiod and temperature in the control of growth and dormancy of Prunus species. Sci. Hortic. 2008, 115, 309–314. [Google Scholar] [CrossRef]
- Chmielewski, F.M.; Götz, K.P. Identification and Timing of Dormant and Ontogenetic Phase for Sweet Cherries in Northeast Germany for Modelling Purposes. J. Hortic. 2017, 4, 205. [Google Scholar] [CrossRef] [Green Version]
- Fadón, E.; Fernandez, E.; Behn, H.; Luedeling, E.A. Conceptual Framework for Winter Dormancy in Deciduous Trees. Agronomy 2020, 10, 241. [Google Scholar] [CrossRef] [Green Version]
- Chmielewski, F.M.; Götz, K.P. Metabolites in cherry buds to detect winter dormancy. Metabolites 2022, 12, 247. [Google Scholar] [CrossRef]
- Hänninen, H.; Kramer, K.; Tanino, K.; Zhang, R.; Wu, J.; Fu, Y.H. Experiments are necessary in process-based tree phenology modelling. Trends Plant Sci. 2019, 24, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Chuine, I.; Cortazar-Atauri, I.G.; Kramer, K.; Hänninen, H. Plant development models. In Phenology: An Integrative Environmental Science, 2nd ed.; Schwartz, M.D., Ed.; Springer Science and Business Media B.V.: Dordrecht, The Netherlands, 2013; pp. 275–293. [Google Scholar]
- Yang, Q.; Gao, Y.; Wu, X.; Bai, T.M.S.; Teng, Y. Bud endodormancy in deciduous fruit trees: Advances and prospects. Hortic. Res. 2021, 8, 139. [Google Scholar] [CrossRef] [PubMed]
- Hannerz, M.; Ekberg, I.; Norell, L. Variation in chilling requirements for completing bud rest between provenances of Norway spruce. Silvae Genet. 2003, 52, 161–168. [Google Scholar]
- Alburquerque, N.; García-Montiel, F.; Carrillo, A.; Burgos, L. Chilling and heat requirements of sweet cherry cultivars and the relationship between altitude and the probability of satisfying the chill requirements. Environ. Exp. Bot. 2008, 64, 162–170. [Google Scholar] [CrossRef]
- Richardson, E.A.; Seeley, S.D.; Walker, D.R. A model for estimating the completion of rest for Redhaven and Elberta peach trees. Hortscience 1974, 9, 331–332. [Google Scholar]
- Cannell, M.G.R.; Smith, R.I. Thermal time, chill days and prediction of budburst in Picea sitchensis. J. Appl. Ecol. 1983, 20, 951–963. [Google Scholar] [CrossRef]
- Murray, M.B.; Cannell, M.G.R.; Smith, R.I. Date of budburst of 15 tree species in Britain following climatic warming. J. Appl. Ecol. 1989, 26, 693–700. [Google Scholar] [CrossRef]
- Landsberg, J.J. Apple fruit bud development and growth; analysis and an empirical model. Ann. Bot. 1974, 38, 1013–1023. [Google Scholar] [CrossRef]
- Harrington, C.A.; Gould, P.J.; St. Clair, J.B. Modeling the effects of winter environment on dormancy release of Douglas-fir. For. Ecol. Manag. 2010, 259, 798–808. [Google Scholar] [CrossRef]
- Vitasse, J.; Basler, D. What role for photoperiod in the bud burst phenology of European beech. Eur. J. Forest Res. 2013, 132, 1–8. [Google Scholar] [CrossRef]
- Fu, Y.H.; Piao, S.; Vitasse, Y.; Zhao, H.; De Boeck, H.J.; Liu, Q.; Yang, H.; Weber, U.; Hänninen, H.; Janssens, I.A. Increased heat requirement for leaf flushing in temperate woody species over 1980–2012: Effects of chilling, precipitation and insolation. Glob. Change Biol. 2015, 21, 2687–2697. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wu, C.; Ciais, P.; Penuelas, J.; Dai, J.; Fu, Y.; Ge, Q. Overestimation of the effect of climatic warming on spring phenology due to misrepresentation of chilling. Nat. Commun. 2020, 11, 4945. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, H.; Blanke, M. Substitution of winter chilling by spring forcing for flowering using sweet cherry as a model crop. Sci. Hortic. 2019, 244, 75–81. [Google Scholar] [CrossRef]
- Menzel, A.; Yuan, Y.; Hamann, A.; Ohl, U.; Matiu, M. Chilling and Forcing from Cut Twigs—How to Simplify Phenological Experiments for Citizen Science. Front. Plant Sci. 2020, 11, 561413. [Google Scholar] [CrossRef]
- Fadón, E.; Fernandez, E.; Do, H.T.; Kunz, A.; Krefting, P.; Luedeling, E. Chill and heat accumulation modulates phenology in temperate fruit trees. Acta Hortic. 2021, 1327, 413–420. [Google Scholar] [CrossRef]
- Hänninen, H. Boreal and Temperate Trees in a Changing Climate: Modelling the Ecophysiology of Seasonality, Biometeorology 3; Springer Science and Business Media: Dordrecht, The Netherlands, 2016; p. 342. [Google Scholar]
- Pope, K.S.; Da Silva, D.; Brown, P.H.; DeJong, T.M. A biologically based approach to modeling spring phenology in temperate deciduous trees. Agric. For. Meteorol. 2014, 198–199, 15–23. [Google Scholar] [CrossRef]
- Darbyshire, R.; Pope, K.; Goodwin, I. An evaluation of the chill overlap model to predict flowering time in apple. Sci. Hortic. 2016, 198, 142–149. [Google Scholar] [CrossRef]
- Richardson, A.D.; Anderson, R.S.; Altaf Arain, M.; Barr, A.G.; Bohrer, G.; Chen, G.; Chen, J.M.; Ciais, P.; Davis, K.J.; Desai, A.R.; et al. Terrestrial biosphere models need better representation of vegetation phenology: Results from the North American Carbon Program Site Synthesis. Glob. Change Biol. 2012, 18, 566–584. [Google Scholar] [CrossRef] [Green Version]
- Chmielewski, F.M.; Götz, K.P. Performance of models for the beginning of sweet cherry blossom under current and changed climate conditions. Agric. For. Meteorol. 2016, 218–219, 85–91. [Google Scholar] [CrossRef]
- Basler, D. Evaluating phenological models for the prediction of leaf-out dates in six temperate tree species across central Europe. Agric. For. Meteorol. 2016, 217, 10–21. [Google Scholar] [CrossRef]
- Chmielewski, F.M.; Baldermann, S.; Götz, K.P.; Homann, T.; Gödeke, K.; Schumacher, F.; Huschek, G.; Rawel, H.M. Abscisic Acid Related Metabolites in Sweet Cherry Buds (Prunus avium L.). J. Hortic. 2018, 5, 221. [Google Scholar] [CrossRef]
- Vimont, N.; Schwarzenberg, A.; Domijan, M.; Donkpegan, A.S.L.; Beauvieux, R.; Le Dantec, L.; Arkoun, M.; Jamois, F.; Yvin, J.C.; Wigge, P.A.; et al. Fine tuning of hormonal signaling is linked to dormancy status in sweet cherry flower buds. Tree Physiol. 2020, 41, 544–561. [Google Scholar] [CrossRef] [PubMed]
- Baron, K.D.; Schroeder, D.F.; Stasolla, C. Transcriptional response of abscisic acid (ABA) metabolism and transport to cold and heat stress applied at the reproductive stage of development in Arabidopsis thaliana. Plant Sci. 2012, 188–189, 48–59. [Google Scholar] [CrossRef] [PubMed]
- Tylewicz, S.; Petterle, A.; Marttila, S.; Miskolczi, P.; Azeez, A.; Singh, R.K.; Immanen, J.; Mähler, N.; Hvidsten, T.R.; Eklund, D.M.; et al. Photoperiodic control of seasonal growth is mediated by ABA acting on cell–cell communication. Science 2018, 360, 212–215. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Sherif, S.M. Hormonal orchestration of bud dormancy cycle in deciduous woody perennials. Front. Plant Sci. 2019, 10, 1136. [Google Scholar] [CrossRef]
- Beil, I.; Kreyling, J.; Meyer, C.; Lemcke, N.; Malyshev, A.V. Late to bed, late to rise—Warmer autumn temperatures delay spring phenology by delaying dormancy. Glob. Change Biol. 2021, 27, 5806–5817. [Google Scholar] [CrossRef]
- Pan, W.; Liang, J.; Sui, J.; Li, J.; Liu, C.; Xin, Y.; Zhang, Y.; Wang, S.; Zhao, Y.; Zhang, J.; et al. ABA and bud dormancy in perennials: Current knowledge and future perspective. Genes 2021, 12, 1635. [Google Scholar] [CrossRef]
- Stass, A.; Horst, W.J. Chemistry, Biochemistry, and Biology of 1–3 Beta Glucans and Related Polysaccharides; Academic Press: Cambridge, MA, USA, 2009; ISBN 978-0-12-373971-1. [Google Scholar] [CrossRef]
- Vimont, N.; Fouché, M.; Campoy, J.A.; Tong, M.; Arkoun, M.; Yvin, J.C.; Wigge, P.A.; Dirlewanger, E.; Cortijo, S.; Wenden, B. From bud formation to flowering: Transcriptomic state defines the cherry developmental phases of sweet cherry bud Dormancy. BMC Genom. 2019, 20, 974. [Google Scholar] [CrossRef] [Green Version]
- Nambara, E.; Marion-Poll, A. Abscisic acid biosynthesis and catabolism. Ann. Rev. Plant Biol. 2005, 56, 165–185. [Google Scholar] [CrossRef] [Green Version]
- Burla, B.; Pfrunder, S.; Nagy, R.; Francisco, R.M.; Lee, Y.; Martinoia, E. Vacuolar transport of abscisic acid glucosyl ester is mediated by ATP-binding cassette and proton-antiport mechanisms in Arabidopsis. Plant Physiol. 2013, 163, 1446–1458. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Kang, J.Y.; Cho, D.I.; Park, J.H.; Kim, S.Y. ABF2, an ABRE-binding bZIP factor, is an essential component of glucose signaling and its overexpression affects multiple stress tolerance. Plant J. 2004, 40, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Meier, U. Growth Stages of Mono-and Dicotyledonous Plants: BBCH Monograph; Open Agrar Repositorium: Quedlinburg, Germany, 2018; p. 204. [Google Scholar] [CrossRef]
- Fadón, E.; Herrero, M.; Rodrigo, J. Flower development in sweet cherry framed in the BBCH scale. Sci. Hortic. 2015, 192, 141–147. [Google Scholar] [CrossRef] [Green Version]
- Fishman, S.; Erez, A.; Couvillon, G.A. The temperature dependence of dormancy breaking in plants: Mathematical analysis of a two-step model involving a cooperative transition. J. Theor. Biol. 1987, 124, 473–483. [Google Scholar] [CrossRef]
- Fishman, S.; Erez, A.; Couvillon, G.A. The temperature-dependence of dormancy breaking in plants—Computer-simulation of processes studied under controlled temperatures. J. Theor. Biol. 1987, 126, 309–321. [Google Scholar] [CrossRef]
- Erez, A.; Fishman, S.; Linsley-Noakes, G.C.; Allan, P. The dynamic model for rest completion in peach buds. Acta Hortic. 1990, 276, 165–174. [Google Scholar] [CrossRef]
- Anderson, J.L.; Richardson, E.A.; Kesner, C.D. Validation of chill unit and flower bud phenology models for ‘Montmorency’ sour cherry. Acta Hort. 1986, 184, 71–75. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LF in DOY | t1 in DOY | t1* in DOY | BB in DOY | Duration (LF–t1) in d | Duration (t1–t1*) in d | Duration (t1*–BB) in d | |
|---|---|---|---|---|---|---|---|
| 2011/12 | 312 | 335 | 45 | 105 | 23 | 75 | 60 |
| 2012/13 | 304 | 332 | 85 | 116 | 28 | 119 | 31 |
| 2013/14 | 302 | 323 | 35 | 95 | 21 | 77 | 60 |
| 2014/15 | 322 | 343 | 41 | 111 | 21 | 63 | 70 |
| 2015/16 | 307 | 328 | 61 | 111 | 21 | 98 | 50 |
| 2016/17 | 313 | 341 | 45 | 97 | 28 | 70 | 52 |
| 2017/18 | 305 | 340 | 59 | 108 | 35 | 84 | 49 |
| 2018/19 | 312 | 333 | 45 | 99 | 21 | 77 | 54 |
| Date | 11/06 | 11/30 | 02/21 | 04/15 | - | - | - |
| x | 309.6 | 334.4 | 52.0 | 105.3 | 24.8 | 82.9 | 53.3 |
| s | 6.5 | 6.8 | 15.9 | 7.6 | 5.1 | 17.8 | 11.3 |
| CV | 2.1 | 2.0 | 30.6 | 7.2 | 20.8 | 21.5 | 21.2 |
| Season | CA (244–t1) in CP | CA (t1–t1*) in CP | CA (t1*–BB) in CP | CA (244–BB) in CP | FA (t1–t1*) in GDH | FA (t1*–BB) in GDH | FA (t1–BB) in GDH |
|---|---|---|---|---|---|---|---|
| 2011/12 | 42 | 40 | 50 | 132 | 642 | 3029 | 3671 |
| 2012/13 | 43 | 57 | 17 | 117 | 645 | 3315 | 3960 |
| 2013/14 | 40 | 51 | 42 | 133 | 802 | 3664 | 4466 |
| 2014/15 | 40 | 41 | 49 | 130 | 744 | 3290 | 4034 |
| 2015/16 | 41 | 67 | 36 | 144 | 2460 | 3050 | 5510 |
| 2016/17 | 46 | 37 | 39 | 122 | 312 | 3378 | 3690 |
| 2017/18 | 49 | 53 | 29 | 131 | 763 | 3998 | 4761 |
| 2018/19 | 40 | 54 | 38 | 132 | 711 | 3850 | 4561 |
| x | 42.6 | 50.0 | 37.5 | 130.1 | 884.9 | 3446.8 | 4331.6 |
| s | 3.3 | 10.1 | 10.7 | 8.0 | 654.4 | 357.0 | 623.2 |
| CV | 7.7 | 20.2 | 28.6 | 6.1 | 74.0 | 10.4 | 14.4 |
| Season | ABA(t1) μg/g DW | ABA(t1*) μg/g DW | ABA(OC) μg/g DW | Duration (t1–t1*) in d | ABA Reduction in % | |
|---|---|---|---|---|---|---|
| (t1–t1*) | (t1–OC) | |||||
| 2011/12 | 9.04 | 3.19 | 0.93 | 75 | 64.7 | 89.7 |
| 2012/13 | 6.04 | 2.07 | 0.85 | 119 | 65.8 | 86.0 |
| 2013/14 | 5.60 | 3.19 | 1.09 | 77 | 43.0 | 80.5 |
| 2014/15 | 5.58 | 3.37 | 1.22 | 63 | 39.6 | 78.1 |
| 2015/16 | 8.60 | 3.40 | 0.89 | 98 | 60.5 | 89.7 |
| 2016/17 | 6.19 | 3.15 | 0.92 | 70 | 49.1 | 85.2 |
| 2017/18 | 6.46 | 3.71 | 1.90 | 84 | 42.7 | 70.6 |
| 2018/19 | 6.04 | 3.48 | 0.99 | 77 | 42.3 | 83.5 |
| x | 6.69 | 3.19 | 1.10 | 82.9 | 51.0 | 82.9 |
| s | 1.35 | 0.49 | 0.35 | 17.8 | 10.9 | 6.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chmielewski, F.-M.; Götz, K.-P. ABA and Not Chilling Reduces Heat Requirement to Force Cherry Blossom after Endodormancy Release. Plants 2022, 11, 2044. https://doi.org/10.3390/plants11152044
Chmielewski F-M, Götz K-P. ABA and Not Chilling Reduces Heat Requirement to Force Cherry Blossom after Endodormancy Release. Plants. 2022; 11(15):2044. https://doi.org/10.3390/plants11152044
Chicago/Turabian StyleChmielewski, Frank-M., and Klaus-Peter Götz. 2022. "ABA and Not Chilling Reduces Heat Requirement to Force Cherry Blossom after Endodormancy Release" Plants 11, no. 15: 2044. https://doi.org/10.3390/plants11152044