
Seed Dormancy and Seedling Ecophysiology Reveal the Ecological Amplitude of the Threatened Endemism Picris willkommii (Schultz Bip.) Nyman (Asteraceae)


Abstract
:
1. Introduction
2. Results
2.1. Brief Characteristics of the Natural Population and Its Ability to Produce Seeds
2.1.1. Seasonal Variation of Natural Population Density and Seed Production
2.1.2. Physiological Activity (Gas Exchange)
2.2. Seed Analysis
2.2.1. Seed Viability
2.2.2. Effect of Environmental Conditions and Pre-Germination Treatments on Seed Germination
2.2.3. Effect of the Substrate, the Time since the Seed Collection and the Seed Size on Germination
2.3. Effect of Environmental Variables on Plant Development
2.3.1. Effect of Air Temperature and Light Radiation on Morpho–Physiological Traits
2.3.2. Effect of Substrate and Mineral Nutrients on Plant Growth
2.3.3. Effect of Planting Time under Field Conditions on Plant Development
3. Discussion
3.1. Brief Ecological Characteristics of the Natural Population and Seed Production
3.2. Seed Viability and Germination
3.3. Effect of Environmental Variables on Plant Development
3.4. Field Development after Outplanting
4. Materials and Methods
4.1. Brief Ecological Characteristics of the Natural Population
4.1.1. Seasonal Variation in Natural Population Density and Seed Production
4.1.2. Physiological Activity (Gas Exchange)
4.1.3. Data Analysis of Eco–Physiological Characteristics
4.2. Seed Analysis
4.2.1. Seed Viability
4.2.2. Effect of Environmental Conditions and Pre-Germination Treatments on Germination
- First, a germination test at different temperatures under white light, at 10, 15, 20, 25 and 30 °C, in order to determine the optimal germination temperature.
- Second, germination and pre-germination treatments: once the optimal germination temperature was established in the previous test (close to 20 °C), germination tests were carried out at that temperature applying various germination and pre-germination treatments. The germination treatments were gibberellic acid (GA3) at 500 mg L–1, KNO3 at 0.2%, and type of light radiation, as well as variable temperature (20/14 °C, day/night). The pre-germination treatments were warm and cold stratification.
- Third, a germination test using peripheral seeds collected in October 2006, directly from the dry plant (the temporary air seed bank).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Germination Conditions | Collection Date and Seed Type | |||||
|---|---|---|---|---|---|---|
| June | 2005 | June | 2006 | October 2006 | ||
| Temperature | Other Conditions | Central | Peripheral | Central | Peripheral | Peripheral |
| 10 °C | WL + H2Od | X | X | X | X | |
| 15 °C | WL + H2Od | X | X | X | X | |
| 20 °C | WL + H2Od | X | X | X | X | X |
| 25 °C | WL + H2Od | X | X | X | X | |
| 30 °C | WL + H2Od | X | X | X | X | |
| WL + GA3 (500 mg L–1) | X | X | X | X | X | |
| WL + KNO3 (0.2%) | X | X | X | X | X | |
| Darkness + H2Od | X | X | X | X | ||
| 20 °C | Red + H2Od | X | X | |||
| Far-Red + H2Od | X | X | ||||
| WL + 20/14 °C + H2Od | X | X | X | X | ||
| Pre3°C + WL + H2Od | X | X | X | X | ||
| Pre30°C + WL + H2Od | X | X | X | X | ||
| Pre30/3°C + WL + H2Od | X | X | X | X | ||
4.2.3. Effect of the Substrate, the Time since the Seed Collection and the Seed Size on Germination
- (a)
- Effect of substrate: On the one hand, in February 2008, the effect on seed germination of the six most abundant soil types in the natural range of the species (Table 12) was tested, and an inert substrate was added as a control (commercial white perlite). This test was carried out at 20 °C, and only peripheral seeds collected in June 2005 were used (Figure S8): 200 seeds per treatment (4 pots of 50 seeds per treatment), moistened with distilled water, randomly distributed in the greenhouse (average temperature 20–22 °C and 10.5–11.5 h photoperiod during the test).
- (b)
- Effect of time since seed collection: On the other hand, in July 2008, the germination potential of the seeds collected and stored at 2–3 °C in the dark for different periods of time was evaluated. Therefore, the storage time of each seed batch was 3, 2, 1 years or 1 month, depending on whether they were collected in 2005, 2006, 2007 or 2008, respectively. Germination conditions were the same as those described above for the soil type test, but central and peripheral seeds were assessed (200 seeds per treatment and seed type).
- (c)
- Effect of seed size: Finally, on July 2008, the effect of the seed size (central and peripheral seeds) on the seedling size was tested. The fresh mass of the seeds (FMs), as well as the fresh mass (FMp), the dry mass (DMp) and the leaf area (LAp) of seedlings at 8 days after germination were assessed. Additionally, the dry mass of leaves (DMl) and the specific leaf area (SLA = LAp/DMl) were determined. This test was performed under laboratory conditions (20 °C, 11 h photoperiod, 4 Petri dishes per seed type).
4.2.4. Data Analysis of Seed Viability and Germination
- Xijkl = μ + STi + TRj + Yk + (ST × TR)ij + (ST × Y)ik + (TR × Y)jk + (ST × TR × Y)ijk + εijkl
- Xijk = μ + STi + Yj + (ST × Y)ij + εijk
- Xij = μ + STi + εij; or Xij = μ + TRi + εij
4.3. Effect of Environmental Variables on Plant Development
4.3.1. Air Temperature and Light Radiation
4.3.2. Substrate and Mineral Nutrients
- Six treatments corresponding to unfertilized natural soils (FL1, FL2, SLT, ABM, RSC and WSD).
- Nine mineral nutrition treatments on inert substrate (mixture of perlite: unfertilized Sphagnum peat: washed river sand (3:1:1 volume)). They were fertilized once a week with their corresponding nutrient solutions, whose pH were 5.5–5.7, with the following treatments:
- -
- Complete treatment: nutrient solution composed of 126, 71 and 90 mg/L of N, P and K, respectively, also containing all other macro and micronutrients in balanced proportions. Nitrogen was added as N-NO3.
- -
- A treatment reduced in N (1/10N): the amount of nitrogen was reduced to a tenth of that provided in the complete treatment, maintaining the contents of the other nutrients in the same quantities as in the complete treatment.
- -
- Seven treatments, each one lacking a mineral nutrient (–N, –P, –K, –Ca, –Mg, –S, –Fe). Only the indicated nutrient was eliminated for each of them, providing the other nutrients in the same amount that in the complete treatment.
4.3.3. Plant Survival According Planting Time in the Field
4.3.4. Data Analysis on Plant Response to the Environment
5. Conclusions
- -
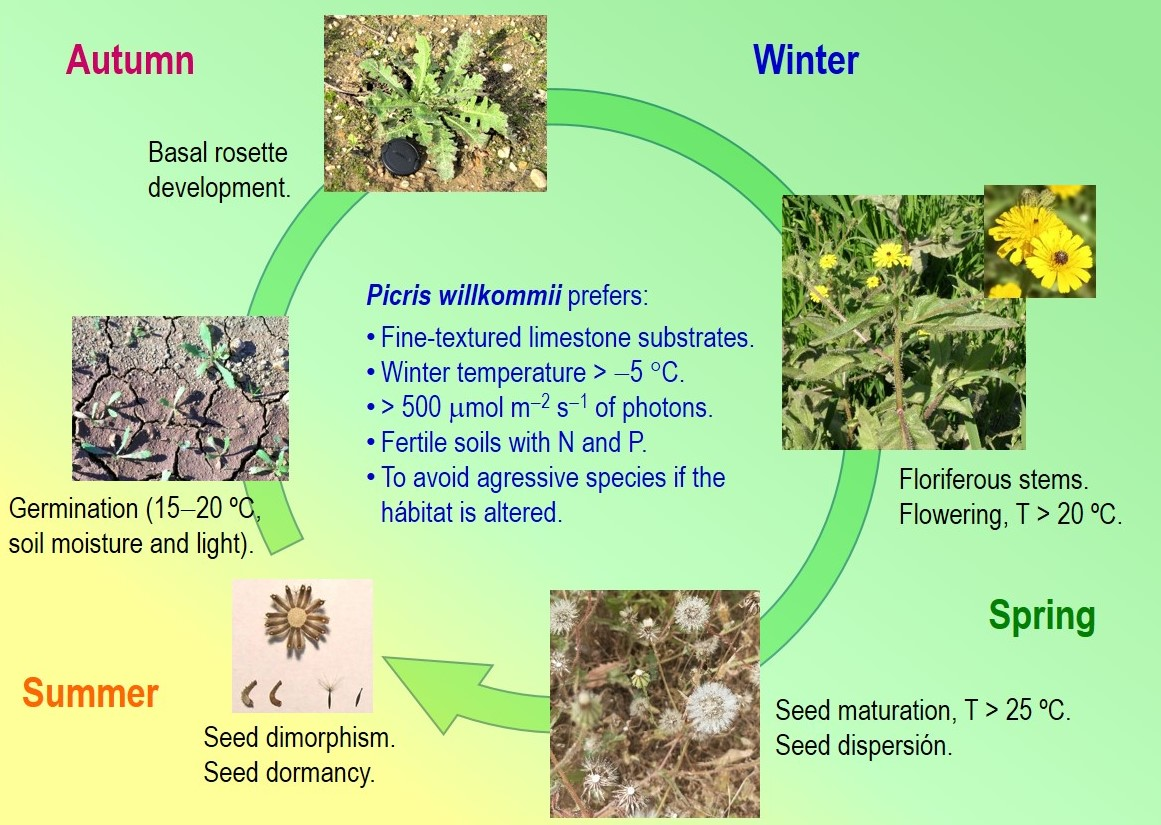
- The timing of plant emergence (early autumn), flowering (spring but extended in time) and seed maturation and dispersal (late spring and extended in time) of Picris willkommii under natural conditions were determined, as well as the environmental conditions (temperature, soil moisture) in which each phase takes place.
- -
- The species presents dimorphism in seeds (central and peripheral), which show different characteristics, not only morphological, related to the size and dispersion medium, but also physiological (germination, dormancy, and photosensibility). This may indicate an evolutionary strategy in order to increase chances of colonization and survival.
- -
- Evidence of physiological endogenous dormancy was found in the seeds at the end of the plant’s maturation period, when they are ready for dispersal.
- -
- At the end of summer, coinciding with the first autumn rains and the drop in temperatures, the plants emerge, which will develop vegetative growth during autumn-winter and will finish the annual reproductive cycle between late spring and the beginning of the following summer.
- -
- The optimum temperature for seed germination of this species is close to 20 °C for both types of seeds (central and peripheral), although with slight differences between them.
- -
- Ripe seeds collected in the field in early summer have high viability and can be stored for a long period of time in a dry and cold environment (2–3 °C).
- -
- Within its natural range, the species prefers limestone soils, with active calcium, clayey or clay loam, and with a certain degree of fertility, mainly with N and P availability.
- -
- The alteration of the ecosystem can lead to the replacement of Picris willkommii by other more aggressive species, both native and invasive.
- -
- Sexual propagation, nursery cultivation and field planting is technically possible. In this case, it is recommended to control other competing herbaceous plants in the early stages of the life cycle.
- -
- Not only the seeds but also the vegetative organs (leaves and stems) are highly photosensitive. It is a species that tolerates shade but prefers exposure to light to optimize its growth and compete with a greater chance of success.
- -
- Winter temperatures below –5 °C harm the normal development of plants and reduce their ability to compete for resources.
- -
- The results obtained are useful for the preservation of the species against the alteration of the ecosystem for any cause, including climate change, as well as being useful for the implementation of conservation and recovery programs for threatened species.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Koornneef, M.; Bentsink, L.; Hilhorst, H. Seed dormancy and germination. Curr. Opin. Plant Biol. 2002, 5, 33–36. [Google Scholar] [CrossRef] [Green Version]
- Escarré, J.; Lepart, J.; Sans, X.; Sentuc, J.J.; Gorse, V. Effects of herbivory on the growth and reproduction of Picris hieracioides in the Mediterranean Region. J. Veg. Sci. 1999, 10, 101–110. [Google Scholar] [CrossRef]
- Grime, J.P. Plant Strategies, Vegetation Processes and Ecosystem Properties, 2nd ed.; John Wiley and Sons Ltd.: Chichester, UK, 2006; 464p. [Google Scholar]
- Matilla, A.; Gallardo, M.; Puga-Hermida, M.I. Structural, physiological and molecular aspects of heterogeneity in seeds: A review. Seed Sci. Res. 2005, 15, 63–76. [Google Scholar] [CrossRef]
- Gandía, M.L.; Del Monte, J.P.; Santín-Montanyá, M.I. Efficiency of Methodologies Used in the Evaluation of the Weed Seed Bank under Mediterranean Conditions. Agronomy 2022, 12, 138. [Google Scholar] [CrossRef]
- Brändel, M. Ecology of Achene Dimorphism in Leontodon saxatilis. Ann. Bot. 2007, 100, 1189–1197. [Google Scholar] [CrossRef] [Green Version]
- Sakaguchi, S.; Horie, K.; Kimura, T.; Nagano, A.J.; Isagi, Y.; Ito, M. Phylogeographic testing of alternative histories of single-origin versus parallel evolution of early flowering serpentine populations of Picris hieracioides L. (Asteraceae) in Japan. Ecol. Res. 2018, 33, 537–547. [Google Scholar] [CrossRef]
- Leubner-Metzger, G. Hormonal and molecular events during seed dormancy release and germination. In The Biology of Seeds: Recent Research Advances; Nicolás, G., Bradford, K.J., Côme, D., Pritchard, H.W., Eds.; CABI Publishing: Wallingford, UK, 2003; pp. 101–112. [Google Scholar]
- Baskin, C.C.; Baskin, J.M. Seeds Ecology, Biogeography, and Evolution of Dormancy and Germination, 2nd ed.; Academic Press: San Diego, CA, USA, 2014. [Google Scholar]
- Monemizadeh, Z.; Ghaderi-Far, F.; Sadeghipour, H.R.; Siahmarguee, A.; Soltani, E.; Torabi, B.; Baskin, C.C. Variation in seed dormancy and germination among populations of Silybum marianum (Asteraceae). Plant Species Biol. 2021, 36, 412–424. [Google Scholar] [CrossRef]
- Zardari, S.; Ghaderi-Far, F.; Sadeghipour, H.R.; Zeinali, E.; Soltani, E.; Baskin, C.C. Deep and intermediate complex morphophysiological dormancy in seeds of Ferula gummosa (Apiaceae). Plant Species Biol. 2019, 34, 85–94. [Google Scholar] [CrossRef]
- Imbert, E. Ecological consequences and ontogeny of seed heteromorphism. Perspect. Plant Ecol. Evol. Syst. 2002, 5, 13–36. [Google Scholar] [CrossRef]
- Slovák, M.; Kučera, J.; Lack, H.W.; Ziffer-Berger, J.; Melicharková, A.; Záveská, E.; Vďačný, P. Diversification dynamics and transoceanic Eurasian-Australian disjunction in the genus Picris (Compositae) induced by the interplay of shifts in intrinsic/extrinsic traits and paleoclimatic oscillations. Mol. Phylogenet. Evol. 2018, 119, 182–195. [Google Scholar] [CrossRef]
- Wang, Y.; Wen, Y.; Gao, J. Anatomy and microscopic characteristics of Picris japónica. Rev. Bras. Farmacogn. 2018, 28, 640–646. [Google Scholar] [CrossRef]
- Monteagudo Sánchez-Movellán, F.J.; Butler Sierra, I.; Bastida Millán, F. Ecology, reproductive biology and conservation of Picris willkommii (Schultz Bip.) Nyman (Asteraceae): An endemic, protected species from therophytic grasslands of South-Western Iberian Peninsula. Investg. Agrar. Sist. Rec. For. 2009, 18, 28–41. [Google Scholar] [CrossRef] [Green Version]
- Monteagudo Sánchez de Movellán, F.J.; Butler Sierra, I. Picris willkommii, endemic to southwestern Iberian Peninsula threatened by the destruction of its pastureland habitat. Pastos 2017, 47, 33–42. Available online: http://polired.upm.es/index.php/pastos/article/view/3858 (accessed on 20 May 2022). (In Spanish).
- Molles, M.C., Jr. Ecology: Concepts and Applications, 4th ed.; McGraw-Hill Higher Education: New York, NY, USA, 2008; 604p, ISBN 9780073050829. [Google Scholar]
- Marschner, P. (Ed.) Mineral Nutrition of Higher Plants, 3rd ed.; Elsevier Ltd.: Amsterdam, The Netherlands, 2011; 672p, ISBN 9780123849052. [Google Scholar]
- Verkaar, H.J.; Schenkeveld, A.J. On the ecology of short-lived forbs in chalk grasslands: Semelparity and seed output of some species in relation to various levels of nutrient supply. New Phytol. 1984, 98, 673–682. [Google Scholar] [CrossRef]
- Luzuriaga, A.L.; Escudero, A. What determines emergence and net recruitment in an early succession plant community? Disentangling biotic and abiotic effects. J. Veg. Sci. 2008, 19, 445–456. [Google Scholar] [CrossRef]
- Zuševica, A.; Celma, S.; Neimane, S.; von Cossel, M.; Lazdina, D. Wood-ash fertiliser and distance from drainage ditch affect the succession and biodiversity of vascular plant species in tree plantings on marginal organic soil. Agronomy 2022, 12, 421. [Google Scholar] [CrossRef]
- Huarte, H.R.; Borlandelli, F.; Varisco, D.; Batlla, D. Understanding dormancy breakage and germination ecology of Cynara cardunculus (Asteraceae). Weed Res. 2018, 58, 450–462. [Google Scholar] [CrossRef]
- Scarici, E.; Ruggeri, R.; Provenzano, M.E.; Rossini, F. Germination and performance of seven native wildflowers in the Mediterranean landscape plantings. Ital. J. Agron. 2018, 13, 1040. [Google Scholar] [CrossRef]
- Yuan, X.; Wen, B. Seed germination response to high temperature and water stress in three invasive Asteraceae weeds from Xishuangbanna, SW China. PLoS ONE 2018, 13, e0191710. [Google Scholar] [CrossRef] [Green Version]
- Gijsman, F.; Vitt, P. Seed size and capitulum position drive germination and dormancy responses to projected warming for the threatened dune endemic Cirsium pitcheri (Asteraceae). Ecol. Evol. 2021, 11, 955–966. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M.; Van Auken, O.W. Dormancy-breaking and germination requirements in seeds of the summer annual Palafoxia callosa (Asteraceae) from Central Texas. Southwest. Nat. 1999, 44, 272–277. Available online: https://www.jstor.org/stable/30055222 (accessed on 10 May 2022).
- Schütz, W.; Milberg, P.; Lamont, B.B. Seed dormancy, after-ripening and light requirements of four annual Asteraceae in South-western Australia. Ann. Bot. 2002, 90, 707–714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoyle, G.L.; Daws, M.I.; Steadman, K.J.; Adkins, S.W. Mimicking a semi-arid tropical environment achieves dormancy alleviation for seeds of Australian native Goodeniaceae and Asteraceae. Ann. Bot. 2008, 101, 701–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoyle, G.L.; Daws, M.I.; Steadman, K.J.; Adkins, S.W. Pre- and post-harvest influences on physiological dormancy alleviation of an Australian Asteraceae species: Actinobole uliginosum (A. Gray) H. Eichler. Seed Sci. Res. 2008, 18, 191–199. [Google Scholar] [CrossRef]
- Karlsson, L.M.; Tamado, T.; Milberg, P. Inter-species comparison of seed dormancy and germination of six annual Asteraceae weeds in an ecological context. Seed Sci. Res. 2008, 18, 35–45. [Google Scholar] [CrossRef] [Green Version]
- Dell, N.D.; Long, Q.G.; Albrecht, M.A. Germination traits in the threatened southeastern grassland endemic, Marshallia mohrii (Asteraceae). Castanea 2019, 84, 212–223. [Google Scholar] [CrossRef]
- Zhang, K.; Yao, L.; Zhang, Y.; Tao, J. Achene heteromorphism in Bidens pilosa (Asteraceae): Differences in germination and possible adaptive significance. AoB Plants 2019, 11, plz025. [Google Scholar] [CrossRef] [Green Version]
- Honda, Y.; Katoh, K. Strict requirement of fluctuating temperatures as a reliable gap signal in Picris Hieracioides var. Japonica seed germination. Plant Ecol. 2007, 193, 147–156. Available online: https://www.jstor.org/stable/40212982 (accessed on 10 May 2022). [CrossRef]
- Guillemin, J.P.; Gardarin, A.; Granger, S.; Reibel, C.; Munier-Jolain, N.; Colbach, N. Assessing potential germination period of weeds with base temperatures and base water potentials. Weed Res. 2013, 53, 76–87. [Google Scholar] [CrossRef]
- Albert, Á.J.; Götzenberger, L.; Jongepierová, I.; Konečná, M.; Lőkkösné-Kelbert, B.; Májeková, M.; Mudrák, O.; Klimešová, J. Restoration of ecosystem functions: Seed production in restored and ancient grasslands. Appl. Veg. Sci. 2021, 24, e12597. [Google Scholar] [CrossRef]
- Picciau, R.; Serra, S.; Porceddu, M.; Bacchetta, G. Seed traits and germination behaviour of four Sardinian populations of Helichrysum microphyllum subsp. Tyrrhenicum (Asteraceae) along an altitudinal gradient. Plant Biol. 2019, 21, 498–506. [Google Scholar] [CrossRef] [Green Version]
- Rees, M.; Childs, D.Z.; Metcalf, J.C.; Rose, K.E.; Sheppard, A.W.; Grubb, P.J. Seed dormancy and delayed flowering in monocarpic plants: Selective interactions in a stochastic environment. Am. Nat. 2006, 168, E53–E71. Available online: https://www.jstor.org/stable/10.1086/505762 (accessed on 17 May 2022). [CrossRef] [Green Version]
- De Waal, C.; Anderson, B.; Ellis, A.G. Dispersal, dormancy and life-history tradeoffs at the individual, population and species levels in southern African Asteraceae. New Phytol. 2016, 210, 356–365. [Google Scholar] [CrossRef] [Green Version]
- Iriskhanova, Z.I.; Ataeva, A.A.; Molochaeva, L.G. Ecological and cenotic analysis of the family Asteraceae in the Chechen Republic. IOP Conf. Ser. Earth Environ. Sci. 2021, 867, 012064. [Google Scholar] [CrossRef]
- Andersen, M.C. An analysis of variability in seed settling velocities of several wind-dispersed Asteraceae. Am. J. Bot. 1992, 79, 1087–1091. Available online: https://www.jstor.org/stable/2445206 (accessed on 17 May 2022). [CrossRef]
- Kadereit, J.W.; Abbott, R.J. Plant speciation in the Quaternary. Plant Ecol. Divers. 2021, 14, 105–142. [Google Scholar] [CrossRef]
- Honda, Y. Ecological correlations between the persistence of the soil seed bank and several plant traits, including seed dormancy. Plant Ecol. 2008, 196, 301–309. Available online: https://www.jstor.org/stable/40305496 (accessed on 10 May 2022). [CrossRef]
- Andersson, S. The relationship between seed dormancy, seed size and weediness, in Crepis tectorum (Asteraceae). Oecologia 1990, 83, 277–280. Available online: https://www.jstor.org/stable/4219330 (accessed on 10 May 2022). [CrossRef]
- Brändel, M. The role of temperature in the regulation of dormancy and germination of two related summer-annual mudflat species. Aquat. Bot. 2004, 79, 15–32. [Google Scholar] [CrossRef]
- Tapias, R.; Gil, L.; Fuentes-Utrilla, P.; Pardos, J.A. Canopy seed banks in Mediterranean pines of southeastern Spain: A comparison between Pinus halepensis Mill., P. pinaster Ait., P. nigra Arn. and P. pinea L. J. Ecol. 2001, 89, 629–638. [Google Scholar] [CrossRef]
- MacArthur, R.H.; Wilson, E.O. The Theory of Island Biogeography; 1967, reprinted 2001; Princeton University Press: Princeton, NJ, USA, 2001; 224p, ISBN 9780691088365. [Google Scholar]
- Nur, M.; Baskin, C.C.; Lu, J.J.; Tan, D.Y.; Baskin, J.M. A new type of non-deep physiological dormancy: Evidence from three annual Asteraceae species in the cold deserts of Central Asia. Seed Sci. Res. 2014, 24, 301–314. [Google Scholar] [CrossRef]
- Pliszko, A.; Kostrakiewicz-Gierałt, K. Effect of cold stratification on seed germination in Solidago × niederederi (Asteraceae) and its parental species. Biologia 2018, 73, 945–950. [Google Scholar] [CrossRef]
- Bu, H.; Chen, X.; Xu, X.; Liu, K.; Jia, P.; Du, G. Seed mass and germination in an Alpine Meadow on the Eastern Tsinghai-Tibet Plateau. Plant Ecol. 2007, 191, 127–149. [Google Scholar] [CrossRef]
- Li, X.; Song, X.; Zhao, J.; Lu, H.; Qian, C.; Zhao, X. Shifts and plasticity of plant leaf mass per area and leaf size among slope aspects in a subalpine meadow. Ecol. Evol. 2021, 11, 14042–14055. [Google Scholar] [CrossRef]
- Liang, J.; Xia, J.; Liu, L.; Wan, S. Global patterns of the responses of leaf-level photosynthesis and respiration in terrestrial plants to experimental warming. J. Plant Ecol. 2013, 6, 437–447. [Google Scholar] [CrossRef] [Green Version]
- Jacob, V.; Zhang, H.; Churchill, A.C.; Yang, J.; Choat, B.; Medlyn, B.E.; Power, S.A.; Tissue, D.T. Warming reduces net carbon gain and productivity in Medicago sativa L. and Festuca arundinacea. Agronomy 2020, 10, 1601. [Google Scholar] [CrossRef]
- Zhou, Z.; Su, P.; Wu, X.; Shi, R.; Ding, X. Leaf and community photosynthetic carbon assimilation of alpine plants under in-situ warming. Front. Plant Sci. 2021, 12, 690077. [Google Scholar] [CrossRef] [PubMed]
- Crous, K.Y.; Uddling, J.; De Kauwe, M.G. Temperature responses of photosynthesis and respiration in evergreen trees from boreal to tropical latitudes. New Phytol. 2022, 234, 353–374. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; He, N.; Hou, J.; Xu, L.; Liu, C.; Zhang, J.; Wang, Q.; Zhang, X.; Wu, X. Factors influencing leaf chlorophyll content in natural forests at the biome scale. Front. Ecol. Evol. 2018, 6, 64. [Google Scholar] [CrossRef] [Green Version]
- Kati, V.; Karamaouna, F.; Economou, L.; Mylona, P.V.; Samara, M.; Mitroiu, M.-D.; Barda, M.; Edwards, M.; Liberopoulou, S. Sown wildflowers enhance habitats of pollinators and beneficial arthropods in a tomato field margin. Plants 2021, 10, 1003. [Google Scholar] [CrossRef] [PubMed]
- McFarquhar, M. Modeling Group-level repeated measurements of neuroimaging data using the univariate general linear model. Front. Neurosci. 2019, 13, 352. [Google Scholar] [CrossRef]
- FAO. A Guide to Forest Seed Handling; FAO Forestry Paper, 20/2; Food and Agriculture Organization of the United Nations: Rome, Italy, 1985; ISBN 92-5-102291-7. Available online: http://www.fao.org/docrep/006/ad232e/AD232E09.htm (accessed on 22 May 2022).
- Barnes, J.D.; Balaguer, L.; Manrique, E.; Elvira, S.; Davison, A.W. A reappraisal of the use of DMSO for the extraction and determination of chlorophylls a and b in lichens and higher plants. Environ. Exp. Bot. 1992, 32, 85–100. [Google Scholar] [CrossRef]
- Herrmann, H.A.; Schwartz, J.M.; Johnson, G.N. From empirical to theoretical models of light response curves—Linking photosynthetic and metabolic acclimation. Photosynth. Res. 2020, 145, 5–14. [Google Scholar] [CrossRef] [Green Version]
- Andivia, E.; Fernández, M.; Vázquez-Piqué, J.; Alejano, R. Two provenances of Quercus ilex ssp. ballota (Desf) Samp. nursery seedlings have different response to frost tolerance and autumn fertilization. Eur. J. Forest Res. 2012, 131, 1091–1101. [Google Scholar] [CrossRef]






| Collected Year | Seed Type | |
|---|---|---|
| Central | Peripheral | |
| 2005 | 97.2 ± 1.1 | 77.0 ± 2.8 |
| 2006 | 96.2 ± 1.5 | 70.7 ± 3.0 |
| 2007 | 85.5 ± 2.3 | 72.3 ± 2.2 |
| 2008 | 98.7 ± 1.0 | 80.1 ± 3.1 |
| Seed Type | Temperature (°C) | GP (%) | GE (%) | GVCz | MGT50 (Days) |
|---|---|---|---|---|---|
| 10 | 13.3 ± 3.2 c | 7.8 ± 1.9 c | 0.6 ± 0.3 c | 10.0 ± 4.0 a | |
| 15 | 51.0 ± 7.7 b | 41.0 ± 7.2 a | 13.6 ± 4.3 b | 3.5 ± 0.3 a | |
| Central | 20 | 80.0 ± 6.3 a | 50.0 ± 6.2 a | 42.2 ± 7.3 a | 2.5 ± 0.3 a |
| 25 | 56.0 ± 12.6 b | 27.0 ± 5.3 b | 19.9 ± 9.4 b | 7.7 ± 3.5 a | |
| 30 | 13.0 ± 5.6 c | 9.5 ± 4.9 c | 0.3 ± 0.2 c | 12.7 ± 5.3 a | |
| 10 | 3.3 ± 2.0 c | 3.3 ± 2.0 c | 0.1 ± 0.1 c | 8.9 ± 4.0 a | |
| 15 | 17.0 ± 2.1 ab | 9.0 ± 1.2 a | 0.8 ± 0.2 b | 9.0 ± 2.0 a | |
| Peripheral | 20 | 19.5 ± 2.6 a | 10.0 ± 2.2 a | 2.1 ± 0.6 a | 6.1 ± 2.0 a |
| 25 | 7.5 ± 2.9 bc | 6.0 ± 1.7 b | 0.4 ± 0.2 bc | 6.6 ± 3.0 a | |
| 30 | 8.8 ± 7.2 bc | 8.8 ± 7.2 ab | 0.3 ± 0.2 bc | 15.4 ± 5.3 a | |
| Temperature | <0.001 | <0.001 | <0.001 | 0.061 | |
| Seed type | <0.001 | <0.001 | <0.001 | 0.003 | |
| Significance | Year | 0.100 | 0.090 | 0.101 | 0.457 |
| level (p) | T × S | <0.001 | <0.001 | <0.001 | 0.349 |
| T × Y | 0.004 | 0.170 | <0.001 | 0.370 | |
| S × Y | <0.001 | 0.003 | <0.001 | 0.563 | |
| T × S × A | 0.151 | 0.245 | 0.101 | 0.269 |
| GP | GE | GVCz | GVDP | MGT50 | |
|---|---|---|---|---|---|
| GP | 1.000 | ||||
| GE | 0.908 ** | 1.000 | |||
| GVCz | 0.914 ** | 0.828 ** | 1.000 | ||
| GVDP | 0.927 ** | 0.812 ** | 0.991 ** | 1.000 | |
| MGT50 | −0.399 * | −0.400 * | −0.233 | −0.233 | 1.000 |
| Seed Type | Treatment | GP (%) | GE (%) | GVCz | MGT50 (Days) |
|---|---|---|---|---|---|
| 20 °C | 80.0 ± 6.3 b | 50.0 ± 6.2 b | 42.2 ± 7.3 b | 2.5 ± 0.3 a | |
| 20/14 °C | 37.0 ± 6.1 c | 18.5 ± 7.9 cd | 11.7 ± 7.9 c | 9.0 ± 4.4 a | |
| GA | 97.0 ± 1.9 a | 81.9 ± 4.7 a | 83.0 ± 6.2 a | 1.8 ± 0.1 a | |
| Central | KNO3 | 45.1 ± 11.5 c | 26.0 ± 9.0 c | 16.7 ± 8.7 c | 2.4 ± 0.5 a |
| Pre3°C | 5.6 ± 2.0 e | 12.5 ± 6.0 de | 0.4 ± 0.3 d | 5.5 ± 3.5 a | |
| Pre30°C | 22.6 ± 4.0 d | 14.0 ± 1.5 d | 1.8 ± 0.5 d | 2.8 ± 0.7 a | |
| Pre30/3°C | 14.1 ± 3.0 e | 6.0 ± 1.5 e | 1.5 ± 0.1 d | 1.6 ± 0.5 a | |
| Darkness | 75.2 ± 7.8 b | 50.0 ± 7.0 b | 46.7 ± 10.5 b | 1.8 ± 0.5 a | |
| 20 °C | 19.5 ± 2.6 b | 10.0 ± 2.2 b | 2.1 ± 0.6 c | 6.1 ± 2.0 a | |
| 20/14 °C | 16.0 ± 2.7 bc | 9.8 ± 1.6 bc | 1.1 ± 0.5 c | 6.1 ± 1.6 a | |
| GA | 65.0 ± 6.6 a | 30.0 ± 4.2 a | 12.3 ± 2.6 a | 6.5 ± 1.5 a | |
| Peripheral | KNO3 | 54.7 ± 10.9 a | 25.0 ± 8.5 a | 5.6 ± 1.5 b | 8.6 ± 2.9 a |
| Pre3°C | 10.8 ± 1.4 c | 8.5 ± 3.0 c | 1.8 ± 0.8 c | 2.3 ± 0.4 a | |
| Pre30°C | 19.6 ± 7.0 bc | 11.0 ± 3.0 b | 1.7 ± 0.7 c | 4.6 ± 0.2 a | |
| Pre30/3°C | 10.8 ± 1.4 c | 5.8 ± 1.2 c | 1.2 ± 0.3 c | 4.0 ± 1.5 a | |
| Darkness | 3.0 ± 1.0 d | 2.8 ± 0.9 d | 0.1 ± 0.1 d | 4.9 ± 2.0 a | |
| Treatment | <0.001 | <0.001 | <0.001 | 0.054 | |
| Seed type | <0.001 | <0.001 | <0.001 | 0.153 | |
| Significance | Year | 0.062 | 0.053 | 0.098 | 0.645 |
| level (p) | T × S | <0.001 | 0.008 | <0.001 | 0.219 |
| T × Y | 0.069 | 0.159 | 0.830 | 0.612 | |
| S × Y | <0.001 | 0.004 | 0.011 | 0.475 | |
| T × S × Y | 0.806 | 0.765 | 0.745 | 0.823 |
| Seed Type | Substrate (Soil Type) | GP (%) | GE (%) | GVCz | MGT50 (Days) |
|---|---|---|---|---|---|
| Perlite | 29.5 ± 3.2 ab | 16.5 ± 4.6 a | 2.6 ± 1.0 ab | 8.4 ± 2.9 a | |
| FL1 | 31.5 ± 2.2 ab | 18.0 ± 2.9 a | 3.0 ± 0.7 ab | 6.8 ± 1.7 a | |
| Peripheral | FL2 | 24.5 ± 2.2 ab | 12.5 ± 3.6 a | 2.1 ± 0.5 a | 4.9 ± 0.2 a |
| SLT | 30.0 ± 2.2 ab | 14.5 ± 2.2 a | 2.6 ± 0.2 ab | 6.5 ± 0.7 a | |
| ABM | 35.5 ± 2.1 b | 27.0 ± 2.5 a | 5.1 ± 0.7 b | 5.8 ± 0.7 a | |
| RSC | 18.5 ± 2.8 a | 12.5 ± 3.4 a | 1.4 ± 0.4 a | 6.3 ± 1.4 a | |
| WSD | 28.5 ± 4.5 ab | 16.0 ± 7.8 a | 2.1 ± 0.6 a | 7.8 ± 2.6 a | |
| p | substrate | 0.012 | 0.090 | 0.013 | 0.717 |
| Seed Type | Collection Year | GP (%) | GE (%) | VGCz | MGT50 (Days) |
|---|---|---|---|---|---|
| 2008 | 16.0 ± 2.8 c | 16.5 ± 2.4 b | 0.5 ± 0.2 c | 8.5 ± 0.5 a | |
| Central | 2007 | 33.8 ± 7.3 b | 25.0 ± 7.9 b | 4.2 ± 0.8 b | 9.4 ± 1.2 a |
| 2006 | 62.8 ± 4.4 a | 45.0 ± 6.2 a | 22.3 ± 6.3 a | 8.5 ± 0.3 a | |
| 2005 | 71.1 ± 4.3 a | 50.0 ± 6.2 a | 24.7 ± 1.2 a | 7.2 ± 0.2 a | |
| 2008 | 16.0 ± 4.0 b | 14.0 ± 4.7 b | 0.5 ± 0.3 b | 11.9 ± 2.7 a | |
| Peripheral | 2007 | 29.3 ± 4.5 a | 23.0 ± 7.5 a | 1.5 ± 0.6 ab | 10.8 ± 0.8 a |
| 2006 | 26.3 ± 3.8 a | 18.0 ± 2.2 ab | 2.1 ± 0.6 a | 6.1 ± 2.1 a | |
| 2005 | 30.0 ± 4.2 a | 19.0 ± 1.9 a | 2.2 ± 0.5 a | 6.3 ± 0.7 a | |
| Significance | Year | <0.001 | 0.009 | <0.001 | 0.747 |
| level (p) | Seed type | 0.005 | 0.016 | <0.001 | 0.742 |
| Y × S | 0.006 | 0.024 | <0.001 | 0.007 |
| Seed Type | Treatment | GP (%) | GE (%) | VGCz | MGT50 (Days) |
|---|---|---|---|---|---|
| Central | H2Od | 16.0 ± 2.8 | 16.5 ± 2.4 | 0.5 ± 0.2 | 8.5 ± 0.5 |
| GA | 94.0 ± 2.6 | 82 ± 4.7 | 14.7 ± 1.3 | 12.2 ± 0.2 | |
| Peripheral | H2Od | 16.0 ± 4.0 | 14.0 ± 4.7 | 0.5 ± 0.3 | 11.9 ± 2.7 |
| GA | 77.0 ± 7.7 | 69 ± 5.9 | 10.3 ± 1.7 | 11.4 ± 0.3 | |
| Significance | Treatment | <0.001 | <0.001 | <0.001 | 0.280 |
| level (p) | Seed type | 0.099 | 0.122 | 0.062 | 0.385 |
| T × S | 0.099 | 0.281 | 0.060 | 0.183 |
| Seed Type | Collection Year | Total Average | |
|---|---|---|---|
| 2005 | 2006 | ||
| Central | 0.62 ± 0.01 | 0.58 ± 0.02 | 0.60 ± 0.01 |
| Peripheral | 2.84 ± 0.08 | 2.60 ± 0.19 | 2.72 ± 0.11 |
| Seed Type | FMp (mg) | LAp (mm2) | DMp (mg) | SLA (m2 kg−1) |
|---|---|---|---|---|
| Central | 13.2 ± 1.4 | 39.7 ± 3.6 | 0.65 ± 0.07 | 61.5 ± 1.1 |
| Peripheral | 21.8 ± 1.6 | 53.3 ± 4.5 | 1.18 ± 0.09 | 44.9 ± 1.3 |
| Significance level (p) | 0.003 | 0.040 | 0.001 | <0.001 |
| Trait | Chla | Chlb |
|---|---|---|
| Fresh mass basis (mg g−1) | 1.47 ± 0.30 | 0.36 ± 0.07 |
| Dry mass basis (mg g−1) | 16.98 ± 0.51 | 4.12 ± 0.33 |
| Leaf area basis (mg m2) | 490.4 ± 110.8 | 118.6 ± 24.6 |
| Treatment | RD (mm) | Flowers (Number) | DMap (g) | DMr (g) | DMap/DMr |
|---|---|---|---|---|---|
| Without additional fertilization | |||||
| Complete | 4.50 b | 16.8 b | 1.84 b | 0.93 b | 2.02 ab |
| –N | 1.50 a | 2.5 a | 0.08 a | 0.11 a | 0.93 ab |
| 1/10N | 1.50 a | 3.5 a | 1.18 a | 0.13 a | 1.38 ab |
| –P | 1.33 a | 1.5 a | 0.11 a | 0.06 a | 2.67 b |
| –K | 4.67 b | 20.3 b | 1.72 b | 0.98 b | 1.97 ab |
| –Mg | 4.17 b | 17.5 b | 1.66 b | 0.87 b | 2.10 ab |
| –S | 5.17 b | 18.7 b | 1.69 b | 1.12 b | 1.70 ab |
| –Ca | 3.83 b | 18.5 b | 1.75 b | 0.89 b | 2.12 ab |
| –Fe | 4.50 b | 16.7 b | 1.51 b | 0.88 b | 1.87 ab |
| FL1 | 1.00 a | 1.8 a | 0.08 a | 0.26 ab | 0.47 a |
| FL2 | 1.50 a | 2.0 a | 0.12 a | 0.13 a | 1.28 ab |
| SLT | 1.50 a | 2.5 a | 0.11 a | 0.09 a | 1.42 ab |
| ABM | 1.75 a | 3.3 a | 0.18 a | 0.10 a | 1.96 ab |
| RSC | 1.75 a | 2.8 a | 0.16 a | 0.21 ab | 0.76 a |
| WSD | 1.00 a | 2.5 a | 0.10 a | 0.10 a | 1.22 ab |
| p | <0.001 | <0.001 | <0.001 | <0.001 | 0.006 |
| With additional fertilization | |||||
| CompleteF | 5.50 b | 37.5 ab * | 3.14 b * | 1.02 ab | 3.11 ab * |
| FL1F | 5.50 b * | 34.0 ab * | 3.79 bc * | 1.24 ab * | 4.01 b * |
| FL2F | 5.50 b * | 56.0 b * | 4.69 c * | 3.09 b * | 1.57 a |
| SLTF | 3.50 a * | 19.0 a * | 1.70 a * | 0.51 a * | 3.36 ab * |
| ABMF | 4.50 ab * | 30.0 ab * | 3.27 b * | 1.42 ab * | 2.29 a * |
| RSCF | 5.00 ab * | 27.5 ab * | 2.49 ab * | 1.06 ab * | 2.37 a |
| WSDF | 5.00 ab * | 31.5 ab * | 3.35 bc * | 1.20 ab * | 2.81 ab * |
| p | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| Soil Type | ||||||
|---|---|---|---|---|---|---|
| FL1 | FL2 | SLT | ABM | RSC | WSD | |
| pH | 8.95 | 8.66 | 8.32 | 8.6 | 6.07 | 8.74 |
| EC (dS/m) | 0.26 | 0.29 | 0.13 | 0.21 | 0.27 | 0.22 |
| OM (% w/w) | 1.2 | 1.6 | 1.1 | 1.3 | 1.3 | 1.4 |
| C/N | 23.5 | 45.0 | 12.9 | 7.7 | 37.2 | 29.2 |
| CO3 (%) | 20.1 | 20.3 | 0.2 | 21.2 | 0.5 | 0.1 |
| ESP (meq/100 g) | 23.5 | 10.2 | 19.1 | 14.7 | 10.9 | 2.1 |
| Active limestone (%) | 10.0 | 3.84 | 0.10 | 6.32 | 0.11 | 0.0 |
| N (mg kg–1) | 304 | 202 | 493 | 962 | 209 | 278 |
| P (mg kg−1) | 28.2 | 19.6 | 33.6 | 39.4 | 40.8 | 31.3 |
| K (meq/100 g) | 0.7 | 0.1 | 0.1 | 0.2 | 0.2 | 0.05 |
| Ca (meq/100 g) | 33.0 | 38.3 | 7.1 | 40.3 | 3.8 | 7.9 |
| Mg (meq/100 g) | 4.7 | 6.2 | 6.1 | 5.7 | 2.6 | 0.8 |
| Na (meq/100 g) | 1.10 | 0.59 | 0.92 | 0.45 | 0.43 | 0.5 |
| B (mg kg–1) | 3.01 | 2.99 | 2.10 | 2.58 | 2.20 | 2.29 |
| Fe (mg kg–1) | 105.5 | 198.4 | 155.6 | 114.4 | 88.1 | 65.6 |
| Cu (mg kg–1) | 2.94 | 5.51 | 1.81 | 27.2 | 0.67 | 1.16 |
| Zn (mg kg–1) | 1.32 | 0.98 | 1.22 | 1.66 | 1.91 | 3.46 |
| Mn mg kg–1) | 64.5 | 67.9 | 115.2 | 60.5 | 2.70 | 8.18 |
| Clay (%) | 49.8 | 18.9 | 40.2 | 29.9 | 21.3 | 1.4 |
| Silt (%) | 31.8 | 3.4 | 13.2 | 15.8 | 7.7 | 0.4 |
| Sand (%) | 18.4 | 77.6 | 46.6 | 54.3 | 71.0 | 98.2 |
| Texture (USDA) | Clay | Loamy sand | Sandy clay | Sandy clay loam | Sandy clay loam | Sand |
| Comments | Soil with active limestone. Low content of OM, N and P, but high of K, Ca and Mg. | Soil with active limestone. Low content of OM, N, P and K, but high of Ca and Mg. | Alkaline soil. Low content of OM and mineral nutrients, except for Mg. | Soil with active limestone. Low content of OM, N, P and K, but high of Ca and Mg. | Red sandy clay gravelly soil. Low content of OM and mineral nutrients. | Sand poor in mineral nutrients and OM. Low ESP. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernández, M.; Tapias, R. Seed Dormancy and Seedling Ecophysiology Reveal the Ecological Amplitude of the Threatened Endemism Picris willkommii (Schultz Bip.) Nyman (Asteraceae). Plants 2022, 11, 1981. https://doi.org/10.3390/plants11151981
Fernández M, Tapias R. Seed Dormancy and Seedling Ecophysiology Reveal the Ecological Amplitude of the Threatened Endemism Picris willkommii (Schultz Bip.) Nyman (Asteraceae). Plants. 2022; 11(15):1981. https://doi.org/10.3390/plants11151981
Chicago/Turabian StyleFernández, Manuel, and Raúl Tapias. 2022. "Seed Dormancy and Seedling Ecophysiology Reveal the Ecological Amplitude of the Threatened Endemism Picris willkommii (Schultz Bip.) Nyman (Asteraceae)" Plants 11, no. 15: 1981. https://doi.org/10.3390/plants11151981