
Pisum sativum Response to Nitrate as Affected by Rhizobium leguminosarum-Derived Signals

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Effect of Nitrate in Presence or Absence of Rhizobium on Molecular Traits
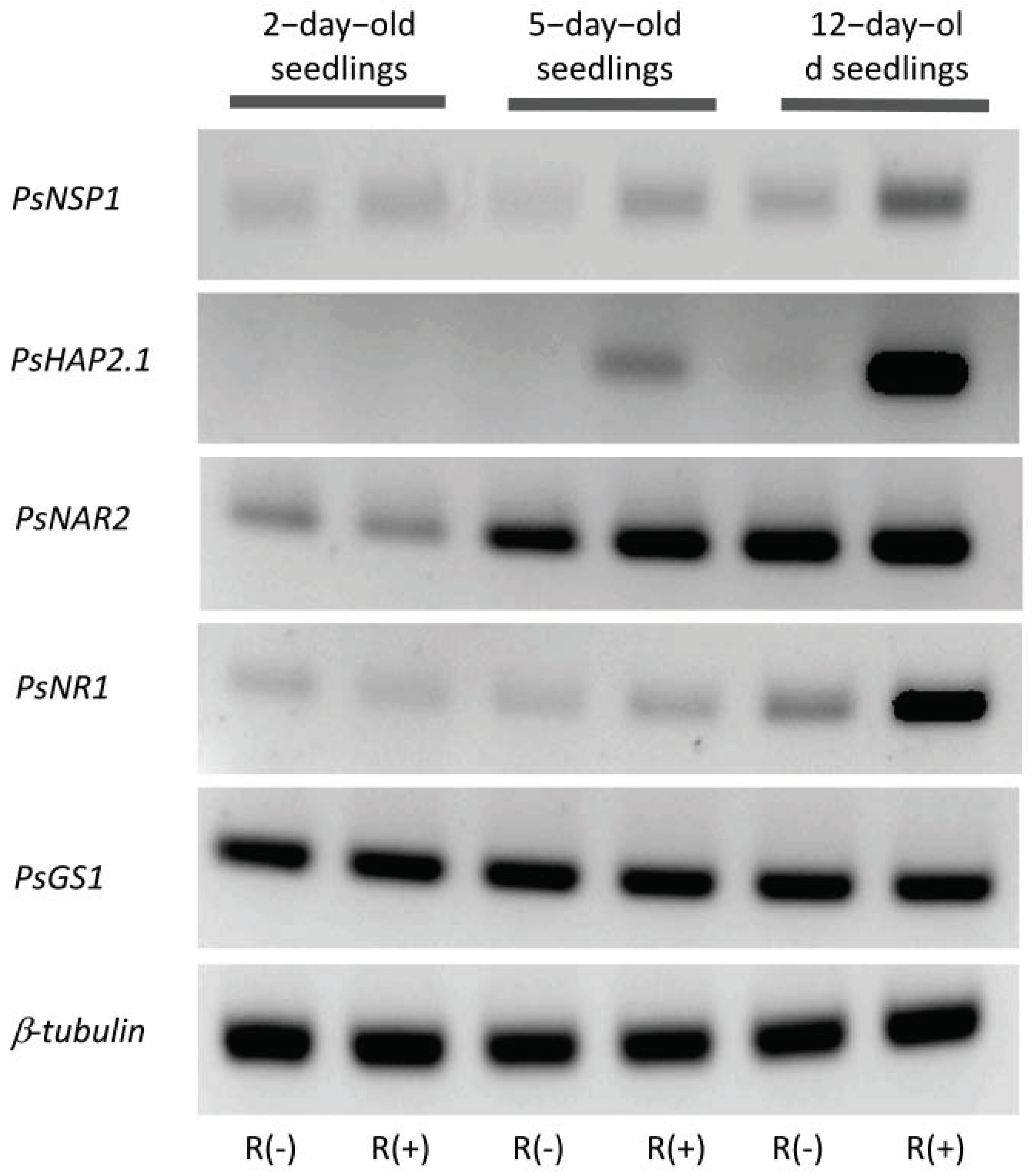
2.1.1. Markers of the Effect of Rhizobium-Derived Signal
2.1.2. Markers of Molecular Response to Nitrate Signal
2.2. Effect of Nitrate in Presence or Absence of Rhizobium on Phenotypic Traits
2.2.1. Root System Length and Branching
2.2.2. Nodules Number
3. Discussion
3.1. Effect of Rhizobium on Pea Seedling Response to NO3− Signal at the Molecular (Transcriptional) Level
3.2. Effect of Rhizobium on Pea Seedling Response to NO3− Signal at Root System Development Level
4. Materials and Methods
4.1. Analysis of Molecular Traits
4.1.1. Seedling Growth Conditions
4.1.2. RNA Extraction and Reverse Transcription
4.1.3. Real-Time Quantitative PCR
4.1.4. Semi Quantitative RT-PCR
4.1.5. Statistical Analysis
4.2. Analysis of Phenotypic Traits
4.2.1. Biological Material and Experimental Conditions
4.2.2. Data Collection
4.2.3. Statistics
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Galloway, J.N.; Cowling, E.B. Reflections on 200 years of Nitrogen, 20 years later: This article belongs to Ambio′s 50th Anniversary Collection. Theme: Eutrophication. Ambio 2021, 50, 745–749. [Google Scholar] [CrossRef] [PubMed]
- Hirsch, A.M.; Lum, M.R.; Downie, J.A. What makes the rhizobia-legume symbiosis so special? Plant Physiol. 2001, 127, 1484–1492. [Google Scholar] [CrossRef] [PubMed]
- Brooker, R.W.; Bennett, A.E.; Cong, W.F.; Daniell, T.J.; George, T.S.; Hallett, P.D.; Hawes, C.; Iannetta, P.P.; Jones, H.G.; Karley, A.J.; et al. Improving intercropping: A synthesis of research in agronomy, plant physiology and ecology. New Phytol. 2015, 206, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Annicchiarico, P.; Nazzicari, N.; Notario, T.; Martin, C.M.; Romani, M.; Ferrari, B.; Pecetti, L. Pea Breeding for Intercropping with Cereals: Variation for Competitive Ability and Associated Traits, and Assessment of Phenotypic and Genomic Selection Strategies. Front. Plant Sci. 2021, 12, 731949. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Forde, B.G. Regulation of Arabidopsis root development by nitrate availability. J. Exp. Bot. 2000, 51, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Rong, H.; Pilbeam, D. Signalling mechanisms underlying the morphological responses of the root system to nitrogen in Arabidopsis thaliana. J. Exp. Bot. 2007, 58, 2329–2338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.Y.; Cheng, Y.H.; Chen, K.E.; Tsay, Y.F. Nitrate Transport, Signaling, and Use Efficiency. Annu. Rev. Plant Biol. 2018, 69, 85–122. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Jiang, Z.; Wang, W.; Qiu, Y.; Zhang, Z.; Liu, Y.; Li, A.; Gao, X.; Liu, L.; Qian, Y.; et al. Nitrate-NRT1.1B-SPX4 cascade integrates nitrogen and phosphorus signalling networks in plants. Nat. Plants 2019, 5, 401–413. [Google Scholar] [CrossRef]
- Pellizzaro, A.; Clochard, T.; Cukier, C.; Bourdin, C.; Juchaux, M.; Montrichard, F.; Thany, S.; Raymond, V.; Planchet, E.; Limami, A.M.; et al. The nitrate transporter MtNPF6.8 (MtNRT1.3) transports abscisic acid and mediates nitrate regulation of primary root growth in Medicago truncatula. Plant Physiol. 2014, 166, 2152–2165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Jennings, A.; Barlow, P.W.; Forde, B.G. Dual pathways for regulation of root branching by nitrate. Proc. Natl. Acad. Sci. USA 1999, 96, 6529–6534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrbach, V.; Remblière, C.; Gough, C.; Bensmihen, S. Lateral root formation and patterning in Medicago truncatula. J. Plant Physiol. 2014, 171, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Oldroyd, G.E.; Downie, J.A. Coordinating nodule morphogenesis with rhizobial infection in legumes. Annu. Rev. Plant Biol. 2008, 59, 519–546. [Google Scholar] [CrossRef]
- Oláh, B.; Brière, C.; Bécard, G.; Dénarié, J.; Gough, C. Nod factors and a diffusible factor from arbuscular mycorrhizal fungi stimulate lateral root formation in Medicago truncatula via the DMI1/DMI2 signalling pathway. Plant J. 2005, 44, 195–207. [Google Scholar] [CrossRef] [PubMed]
- Phillips, D.A.; Torrey, J.G. Studies on cytokinin production by Rhizobium. Plant Physiol. 1972, 49, 11–15. [Google Scholar] [CrossRef] [Green Version]
- Ernstsen, A.; Sandberg, G.; Crozier, A.; Wheeler, C.T. Endogenous indoles and the biosynthesis and metabolism of indole-3-acetic acid in cultures of Rhizobium phaseoli. Planta 1987, 171, 422–428. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Liu, W.; Nandety, R.S.; Crook, A.; Mysore, K.S.; Pislariu, C.I.; Frugoli, J.; Dickstein, R.; Udvardi, M.K. Celebrating 20 Years of Genetic Discoveries in Legume Nodulation and Symbiotic Nitrogen Fixation. Plant Cell 2020, 32, 15–41. [Google Scholar] [CrossRef] [Green Version]
- Gu, B.; Chen, Y.; Xie, F.; Murray, J.D.; Miller, A.J. Inorganic Nitrogen Transport and Assimilation in Pea. Genes 2022, 13, 158. [Google Scholar] [CrossRef]
- Kreplak, J.; Madoui, M.A.; Cápal, P.; Novák, P.; Labadie, K.; Aubert, G.; Bayer, P.E.; Gali, K.K.; Syme, R.A.; Main, D.; et al. A reference genome for pea provides insight into legume genome evolution. Nat. Genet. 2019, 51, 1411–1422. [Google Scholar] [CrossRef] [PubMed]
- Voisin, A.S.; Bourion, V.; Duc, G.; Salon, C. Using an ecophysiological analysis to dissect genetic variability and to propose an ideotype for nitrogen nutrition in pea. Ann. Bot. 2007, 100, 1525–1536. [Google Scholar] [CrossRef] [Green Version]
- Pellizzaro, A.; Clochard, T.; Planchet, E.; Limami, A.M.; Morère-Paven, M.C. Identification and molecular characterization of Medicago truncatula NRT2 and NAR2 families. Physiol. Plant. 2015, 154, 256–269. [Google Scholar] [CrossRef]
- Zang, L.; Morère-Paven, M.C.; Clochard, T.; Porcher, A.; Satour, P.; Mojović, M.; Vidović, M.; Limami, A.M.; Montrichard, F. Nitrate inhibits primary root growth by reducing accumulation of reactive oxygen species in the root tip in Medicago truncatula. Plant Physiol. Biochem. 2020, 146, 363–373. [Google Scholar] [CrossRef] [PubMed]
- Zang, L.; Tarkowski, Ł.; Morère-Paven, M.C.; Zivy, M.; Balliau, T.; Clochard, T.; Bahut, M.; Balzergue, S.; Pelletier, S.; Landès, C.; et al. The Nitrate Transporter MtNPF6.8 is a Master Sensor of Nitrate Signal in the Primary Root Tip of Medicago truncatula. Front. Plant Sci. 2022, 13, 832246. [Google Scholar] [CrossRef] [PubMed]
- Combier, J.P.; Frugier, F.; de Billy, F.; Boualem, A.; El-Yahyaoui, F.; Moreau, S.; Vernié, T.; Ott, T.; Gamas, P.; Crespi, M.; et al. MtHAP2-1 is a key transcriptional regulator of symbiotic nodule development regulated by microRNA169 in Medicago truncatula. Genes Dev. 2006, 20, 3084–3088. [Google Scholar] [CrossRef] [Green Version]
- Alves-Carvalho, S.; Aubert, G.; Carrère, S.; Cruaud, C.; Brochot, A.L.; Jacquin, F.; Klein, A.; Martin, C.; Boucherot, K.; Kreplak, J.; et al. Full-length de novo assembly of RNA-seq data in pea (Pisum sativum L.) provides a gene expression atlas and gives insights into root nodulation in this species. Plant J. 2015, 84, 1–19. [Google Scholar] [CrossRef]
- Wang, Q.; Huang, Y.; Ren, Z.; Zhang, X.; Ren, J.; Su, J.; Zhang, C.; Tian, J.; Yu, Y.; Gao, G.F.; et al. Transfer cells mediate nitrate uptake to control root nodule symbiosis. Nat. Plants 2020, 6, 800–808. [Google Scholar] [CrossRef]
- Léran, S.; Varala, K.; Boyer, J.C.; Chiurazzi, M.; Crawford, N.; Daniel-Vedele, F.; David, L.; Dickstein, R.; Fernandez, E.; Forde, B.; et al. A unified nomenclature of NITRATE TRANSPORTER 1/PEPTIDE TRANSPORTER family members in plants. Trends Plant Sci. 2014, 19, 5–9. [Google Scholar] [CrossRef]
- Orsel, M.; Chopin, F.; Leleu, O.; Smith, S.J.; Krapp, A.; Daniel-Vedele, F.; Miller, A.J. Characterization of a two-component high-affinity nitrate uptake system in Arabidopsis. Physiology and protein-protein interaction. Plant Physiol. 2006, 142, 1304–1317. [Google Scholar] [CrossRef] [Green Version]
- Schulze, J.; Liese, R.; Ballesteros, G.; Casieri, L.; Salinas, G.; Cabeza, R.A. Ammonium acts systemically while nitrate exerts an additional local effect on Medicago truncatula nodules. Plant Sci. 2020, 292, 110383. [Google Scholar] [CrossRef] [PubMed]
- Mergaert, P.; Kereszt, A.; Kondorosi, E. Gene Expression in Nitrogen-Fixing Symbiotic Nodule Cells in Medicago truncatula and other nodulating plants. Plant Cell 2020, 32, 42–68. [Google Scholar] [CrossRef]
- Ho, C.; Lin, S.; Hu, H.; Tsay, Y. CHL1 functions as a nitrate sensor in plants. Cell 2009, 138, 1184–1194. [Google Scholar] [CrossRef] [Green Version]
- Krouk, G.; Lacombe, B.; Bielach, A.; Perrine-Walker, F.; Malinska, K.; Mounier, E.; Hoyerova, K.; Tillard, P.; Leon, S.; Ljung, K.; et al. Nitrate-regulated auxin transport by NRT1.1 defines a mechanism for nutrient sensing in plants. Dev. Cell 2010, 18, 927–937. [Google Scholar] [CrossRef] [PubMed]
- Yong, Z.; Kotur, Z.; Glass, A.D. Characterization of an intact two-component high-affinity nitrate transporter from Arabidopsis roots. Plant J. 2010, 63, 739–748. [Google Scholar] [CrossRef] [PubMed]
- Little, D.Y.; Rao, H.; Oliva, S.; Daniel-Vedele, F.; Krapp, A.; Malamy, J.E. The putative high-affinity nitrate transporter NRT2.1 represses lateral root initiation in response to nutritional cues. Proc. Natl. Acad. Sci. USA 2005, 102, 13693–13698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallardo, K.; Besson, A.; Klein, A.; le Signor, C.; Aubert, G.; Henriet, C.; Térézol, M.; Pateyron, S.; Sanchez, M.; Trouverie, J.; et al. Transcriptional Reprogramming of Pea Leaves at Early Reproductive Stages. Front. Plant Sci. 2019, 10, 1014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishida, H.; Suzaki, T. Nitrate-mediated control of root nodule symbiosis. Curr. Opin. Plant Biol. 2018, 44, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Nishida, H.; Ito, M.; Miura, K.; Kawaguchi, M.; Suzaki, T. Autoregulation of nodulation pathway is dispensable for nitrate-induced control of rhizobial infection. Plant Signal. Behav. 2020, 15, 1733814. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rhizobium | − | + | ||
|---|---|---|---|---|
| Nitrate Concentration | 5 mM | 14 mM | 5 mM | 14 mM |
| Nodules Number | 8 ± 2b | 1 ± 1a | 65 ± 4c | 42 ± 6c |
| Total Root Length (cm) | 842 ± 23 | 796 ± 50 | 794 ± 44 | 850 ± 11 |
| Secondary Roots Number | 49 ± 3 | 55 ± 3 | 53 ± 2 | 45 ± 1 |
| Tertiary Roots Number | 117 ± 14 | 85 ± 16 | 134 ± 18 | 112 ± 13 |
| Branching Ratio | 2.37 ± 0.30 | 1.52 ± 0.23 | 2.50 ± 0.27 | 2.47 ± 0.26 |
| Sample size | 10 | 8 | 9 | 9 |
| Gene ID | Gene Name | Primers | |
|---|---|---|---|
| Psat5g198720 | β-tubulin | Forward | CAGAACAAGAACTCGTCATACT |
| Reverse | AGCCTTCCTCCTGAACATA | ||
| Psat5g063760 | Actin | Forward | CTAAGGGTGAATATGATGAGTCTGG |
| Reverse | GAGACACCAAAAAAGCAACCACATC | ||
| Psat0s741g0280 | PsNSP1 | Forward | AAGCATTGACAAACCAGCGT |
| Reverse | ATGTTGCCCCTTCCACCATA | ||
| Psat6g092920 | PsHAP2.1 | Forward | GCTGAGCCGTACAATCGTTT |
| Reverse | AAGTTGGTCCGTCGTCAGAT | ||
| Psat6g242600 | PsNPF7.1 | Forward | TGCGGAAAATGGGATGTTGT |
| Reverse | CTCCACGGATTGTTTCGAGTC | ||
| Psat2g178360 | PsNPF6.4 | Forward | ATTGCAGTGGTGTTTGTGGC |
| Reverse | ACGACTTTGATTGCCGCTTT | ||
| Psat7g149120 | PsNRT2 | Forward | TGCCTTTTTATCCTGGTTGG |
| Reverse | GTAAAGCAGCGCAAAAATCC | ||
| Psat4g061680 | PsNAR2 | Forward | GGACGATCTCTCAAGGGACA |
| Reverse | TATCAGCATTCGTGCTTTGC | ||
| Psat2g077040 | PsNR1 | Forward | GTCGACGGAAAATTGACGAT |
| Reverse | CCTACCGGGCCTTTTACTTC | ||
| Psat5g120440 | PsGS1 | Forward | TTTGCCGGCATCAACATCAG |
| Reverse | AGCACCATTCCAATCACCCT | ||
| Psat0s690g0040 | PsGS2 | Forward | ACGAGGTAATCAAGAAGGCGA |
| Reverse | ATTGAGCTTCCACGGTTTGC | ||
| Psat5g153000 | PsASNS | Forward | TCACTACGATAAGGGCTGCAA |
| Reverse | GCTTTGATCTTGCGGCATGT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boeglin, L.; Morère Le-Paven, M.-C.; Clochard, T.; Fustec, J.; Limami, A.M. Pisum sativum Response to Nitrate as Affected by Rhizobium leguminosarum-Derived Signals. Plants 2022, 11, 1966. https://doi.org/10.3390/plants11151966
Boeglin L, Morère Le-Paven M-C, Clochard T, Fustec J, Limami AM. Pisum sativum Response to Nitrate as Affected by Rhizobium leguminosarum-Derived Signals. Plants. 2022; 11(15):1966. https://doi.org/10.3390/plants11151966
Chicago/Turabian StyleBoeglin, Laure, Marie-Christine Morère Le-Paven, Thibault Clochard, Joëlle Fustec, and Anis M. Limami. 2022. "Pisum sativum Response to Nitrate as Affected by Rhizobium leguminosarum-Derived Signals" Plants 11, no. 15: 1966. https://doi.org/10.3390/plants11151966