
Biogeographic Patterns of Leaf Element Stoichiometry of Stellera chamaejasme L. in Degraded Grasslands on Inner Mongolia Plateau and Qinghai-Tibetan Plateau

 , , , and
, , , and
Abstract
:
1. Introduction
2. Results
2.1. Pattern of Leaf Ecological Stoichiometry and Soil Physicochemical Properties of S. chamaejasme
2.2. Ecological Stoichiometry Homeostasis of S. chamaejasme in Degraded Grassland
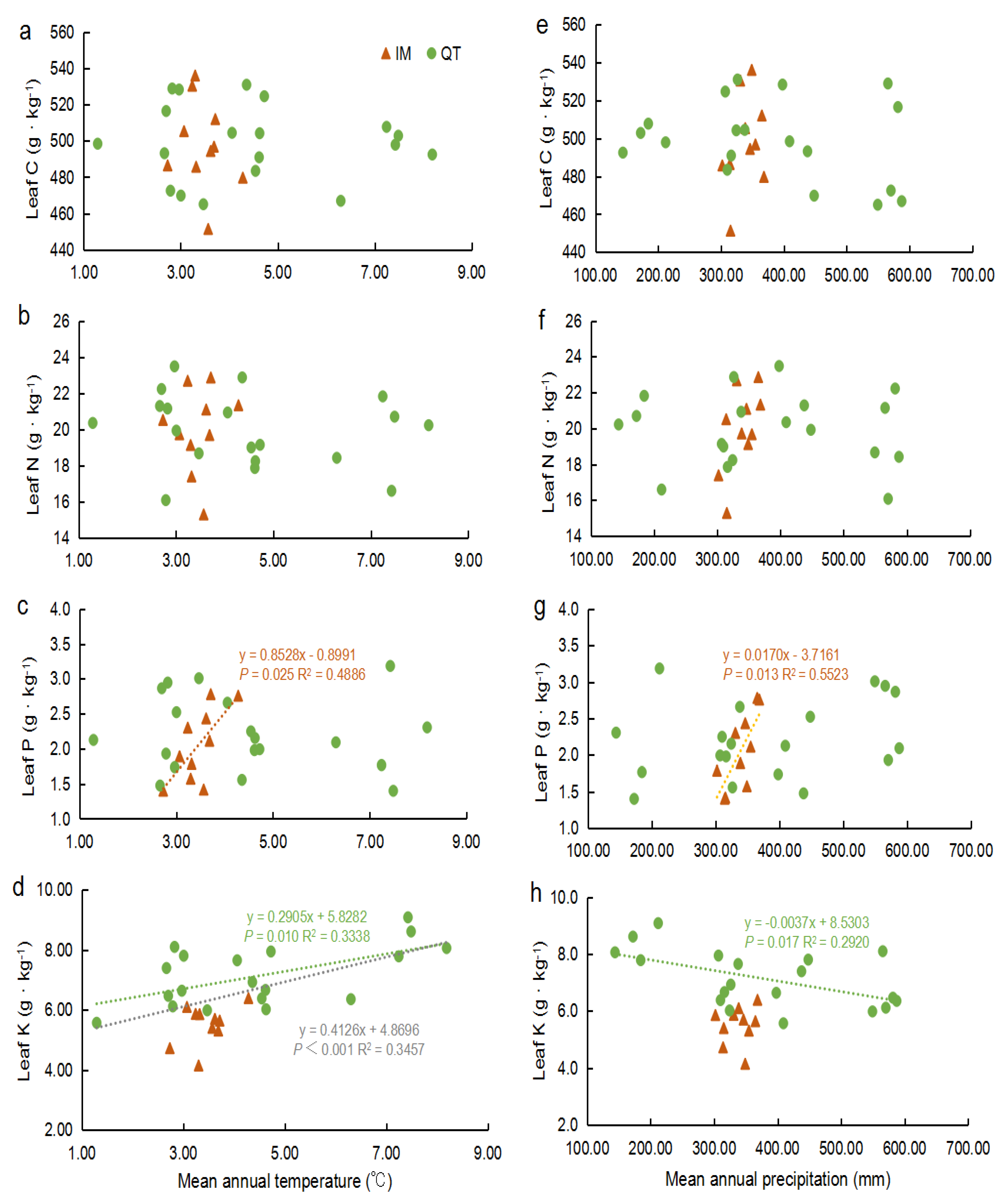
2.3. Spatial Variation of Leaf Elements of S. chamaejasme in Relation to Climatic Factors
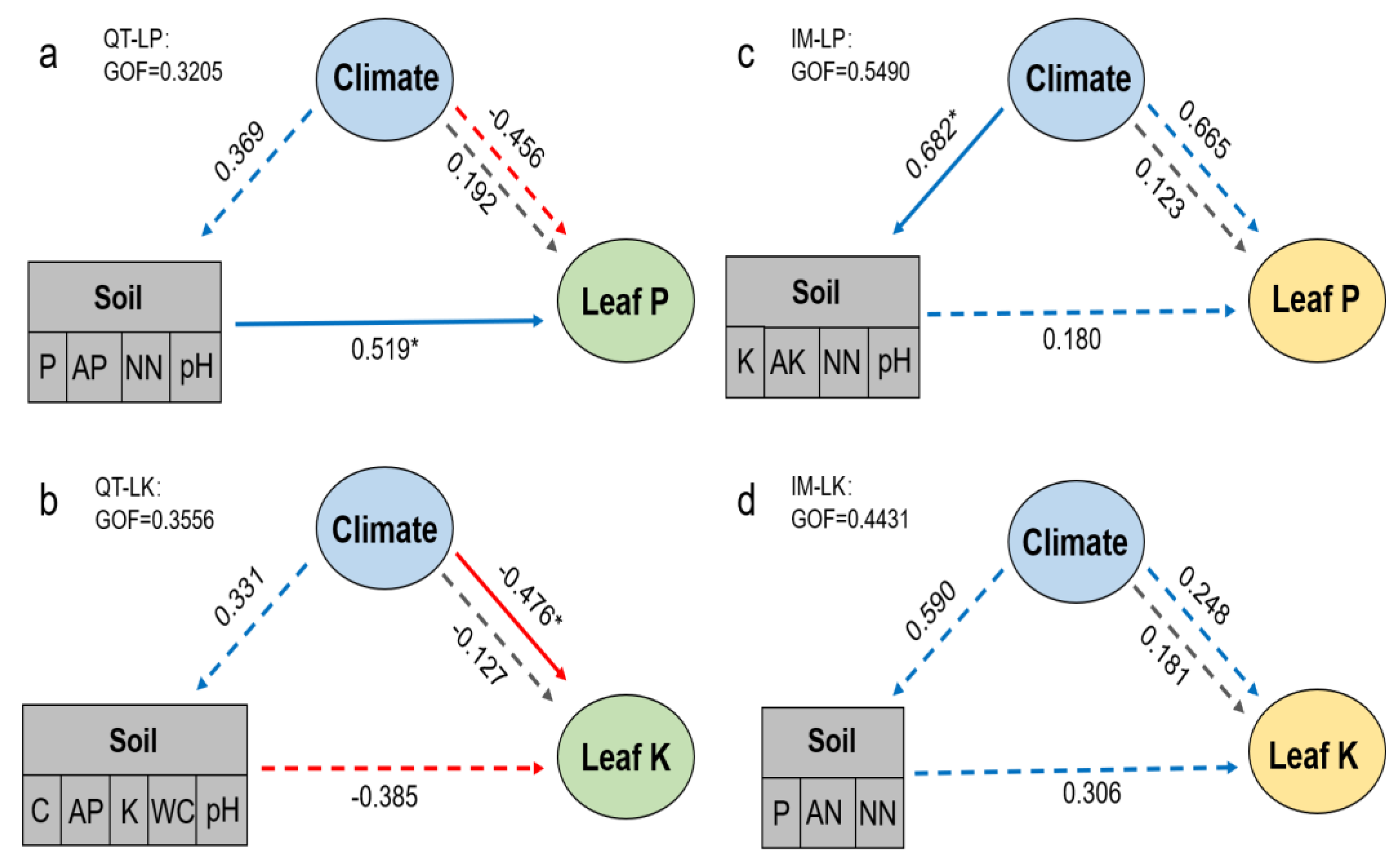
2.4. Relative Roles of Soil and Climatic Factors in Leaf Elements of S. chamaejasme
3. Discussion
3.1. Leaf Ecological Stoichiometry and Soil Physicochemical Properties of S. chamaejasme
3.2. Relationships between Leaf Ecological Stoichiometry and Environmental Variables
4. Materials and Methods
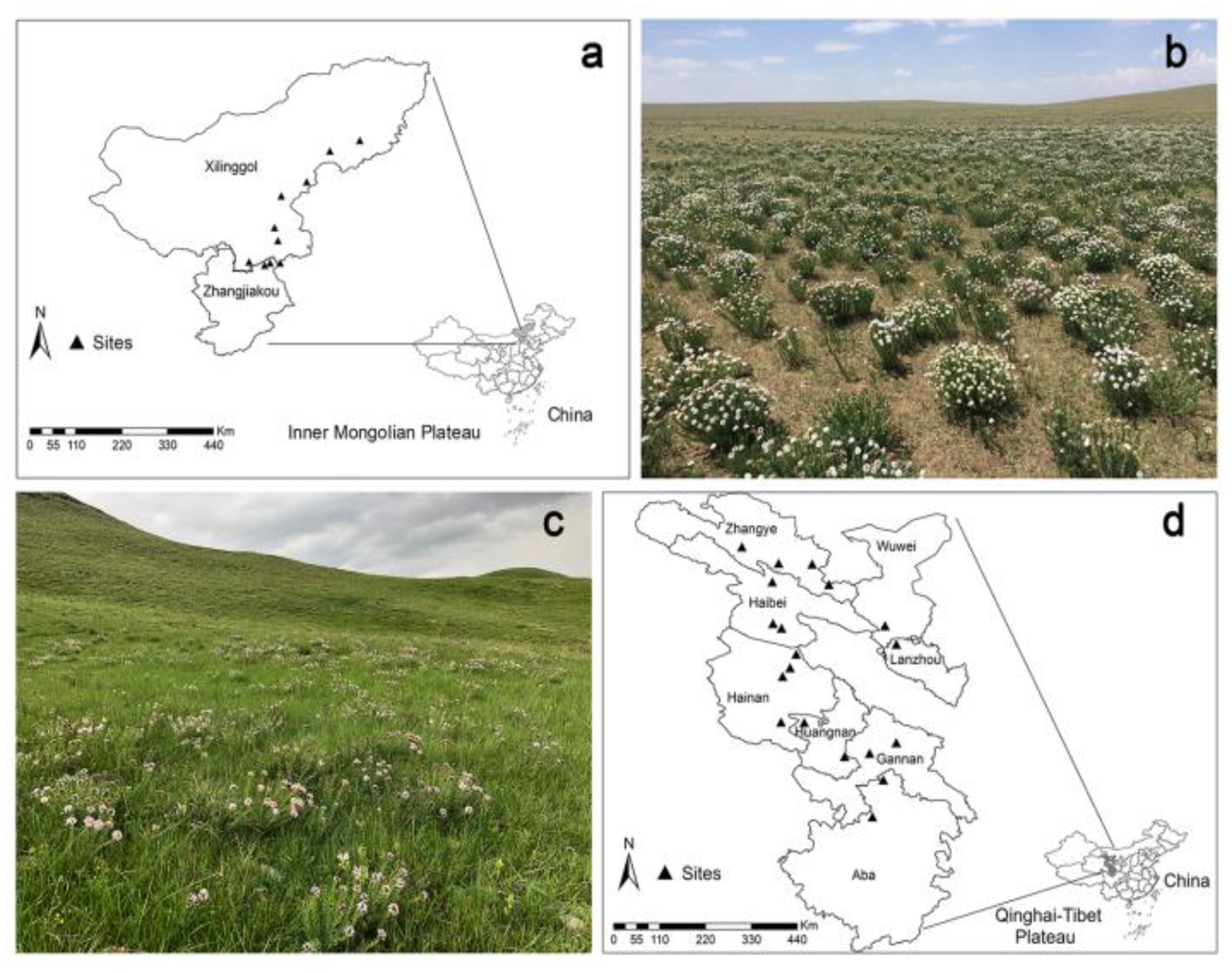
4.1. Study Area
4.2. Plant and Soil Sampling
4.3. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Elser, J.J.; Sterner, R.W.; Gorokhova, E.; Fagan, W.F.; Markow, T.A.; Cotner, J.B.; Harrison, J.F.; Hobbie, S.E.; Odell, G.M.; Weider, L.W. Biological stoichiometry from genes to ecosystems. Ecol. Lett. 2000, 3, 540–550. [Google Scholar] [CrossRef] [Green Version]
- Sterner, R.W.; Elser, J.J. Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere; Princeton University Press: Princeton, NJ, USA, 2002. [Google Scholar]
- Sardans, J.; Janssens, I.A.; Ciais, P.; Obersteiner, M.; Peñuelas, J. Recent advances and future research in ecological stoichiometry. Perspect. Plant Ecol. Evol. Syst. 2021, 50, 125611. [Google Scholar] [CrossRef]
- Güsewell, S. N:P ratios in terrestrial plants: Variation and functional significance. New Phytol. 2004, 164, 243–266. [Google Scholar] [CrossRef]
- Reich, P.B.; Oleksyn, J. Global patterns of plant leaf N and P in relation to temperature and latitude. Proc. Natl. Acad. Sci. USA 2004, 101, 11001–11006. [Google Scholar] [CrossRef] [Green Version]
- He, J.-S.; Wang, L.; Flynn, D.; Wang, X.; Ma, W.; Fang, J. Leaf nitrogen:phosphorus stoichiometry across Chinese grassland biomes. Oecologia 2008, 155, 301–310. [Google Scholar] [CrossRef]
- Wu, P.; Zhou, H.; Cui, Y.C.; Zhao, W.J.; Hou, Y.J.; Zhu, J.; Ding, F.J. Stoichiometric characteristics of leaf nutrients in karst plant species during natural restoration in Maolan national nature reserve, Guizhou, China. J. Sustain. For. 2021, 1–25. [Google Scholar] [CrossRef]
- Sardans, J.; Rivas-Ubach, A.; Peñuelas, J. The elemental stoichiometry of aquatic and terrestrial ecosystems and its relationships with organismic lifestyle and ecosystem structure and function: A review and perspectives. Biogeochemistry 2012, 111, 1–39. [Google Scholar] [CrossRef]
- Chown, S.L.; Gaston, K.J.; Robinson, D. Macrophysiology: Large-scale patterns in physiological traits and their ecological implications. Funct. Ecol. 2004, 18, 159–167. [Google Scholar] [CrossRef]
- Yang, X.J.; Huang, Z.Y.; Zhang, K.L.; Cornelissen, J.H.C. C:N:P stoichiometry of Artemisia species and close relatives across northern China: Unravelling effects of climate, soil and taxonomy. J. Ecol. 2015, 103, 1020–1031. [Google Scholar] [CrossRef]
- Wang, X.G.; Lü, X.T.; Dijkstra, F.A.; Zhang, H.Y.; Wu, Y.N.; Wang, Z.W.; Feng, J.; Han, X.G. Changes of plant N:P stoichiometry across a 3000-km aridity transect in grasslands of northern China. Plant Soil 2019, 443, 107–119. [Google Scholar] [CrossRef]
- Persson, J.; Fink, P.; Goto, A.; Hood, J.M.; Jonas, J.; Kato, S. To be or not to be what you eat: Regulation of stoichiometric homeostasis among autotrophs and heterotrophs. Oikos 2010, 119, 741–751. [Google Scholar] [CrossRef]
- Hessen, D.O.; Agren, G.I.; Anderson, T.R.; Elser, J.J.; De Ruiter, P.C. Carbon sequestration in ecosystems: The role of stoi-chiometry. Ecology 2004, 85, 1179–1192. [Google Scholar] [CrossRef]
- Elser, J.J.; Fagan, W.F.; Kerkhoff, A.J.; Swenson, N.G.; Enquist, B.J. Biological stoichiometry of plant production: Metabolism, scaling and ecological response to global change. New Phytol. 2010, 186, 593–608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Q.; Chen, Q.S.; Elser, J.J.; He, N.P.; Wu, H.H.; Zhang, G.M.; Wu, J.G.; Bai, Y.F.; Han, X.G. Linking stoichiometric homoeostasis with ecosystem structure, functioning and stability. Ecol. Lett. 2010, 13, 1390–1399. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.Y.; Chen, Y.H.; Yan, Z.B.; Han, W.X. Stage-dependent stoichiometric homeostasis and responses of nutrient resorption in Amaranthus mangostanus to nitrogen and phosphorus addition. Sci. Rep. 2016, 6, 37219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, Z.B.; Guan, H.Y.; Han, W.X.; Han, T.S.; Guo, Y.L.; Fang, J.Y. Reproductive organ and young tissues show constrained elemental composition in Arabidopsis thaliana. Ann. Bot. 2016, 117, 431–439. [Google Scholar] [CrossRef] [Green Version]
- Yu, Q.; Elser, J.J.; He, N.P.; Wu, H.H.; Chen, Q.S.; Zhang, G.M.; Han, X.G. Stoichiometric homeostasis of vascular plants in the Inner Mongolia grassland. Oecologia 2011, 166, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Wilcox, K.; La Pierre, K.; Knapp, A.K.; Han, X.G.; Smith, M.D. Stoichiometric homeostasis predicts plant species dominance, temporal stability, and responses to global change. Ecology 2015, 96, 2328–2335. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.; Luo, P.; Wu, N.; Qiu, P.F.; Gao, Y.H.; Chen, H.; Shi, F.S. Stellera chamaejasme L. increases soil N availability, turnover rates and microbial biomass in an alpine meadow ecosystem on the eastern Tibetan Plateau of China. Soil Biol. Biochem. 2009, 41, 86–91. [Google Scholar] [CrossRef]
- Guo, L.Z.; Li, J.H.; He, W.; Liu, L.; Huang, D.; Wang, K. High nutrient uptake efficiency and high water use efficiency facilitate the spread of Stellera chamaejasme L. in degraded grasslands. BMC Ecol. 2019, 19, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.-H.; Zhang, J.-W.; Li, Z.-M.; Sun, H. Genetic diversity of the weed species, Stellera chamaejasme in China inferred from amplified fragment length polymorphism analysis. Weed Biol. Manag. 2015, 15, 165–174. [Google Scholar] [CrossRef]
- Zhao, M.L.; Gao, X.L.; Wang, J.; He, X.L.; Hsn, B. A review of the most economically important poisonous plants to the live-stock industry on temperate grasslands of China. J. Appl. Toxicol. 2013, 33, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; Guo, H.R.; Yang, X.Y.; Xin, A.Y.; Liu, H.Y.; Qin, B. Effect of allelochemicals, soil enzyme activity and environmental factors from Stellera chamaejasme L. on rhizosphere bacterial communities in the northern Tibetan Plateau. Arch. Agron. Soil Sci. 2020, 68, 547–560. [Google Scholar] [CrossRef]
- Guo, L.Z.; Zhao, H.; Zhai, X.J.; Wang, K.L.; Liu, L.; Wang, K.; Huang, D. Study on life history traits of Stellera chamaejasme provide insights into its control on degraded typical steppe. J. Environ. Manag. 2021, 291, 112716. [Google Scholar] [CrossRef]
- Geng, Y.; Wang, Z.H.; Liang, C.Z.; Fang, J.Y.; Baumann, F.; Kuhn, P.; Scholten, T.; He, J.S. Effect of geographical range size on plant functional traits and the relationships between plant, soil and climate in Chinese grasslands. Glob. Ecol. Biogeogr. 2011, 21, 416–427. [Google Scholar] [CrossRef]
- Zhang, H.Y.; Wu, H.H.; Yu, Q.; Wang, Z.W.; Wei, C.Z.; Long, M.; Kattge, J.; Smith, M.; Han, X.G. Sampling Date, Leaf age and root size: Implications for the study of plant C:N:P stoichiometry. PLoS ONE 2013, 8, e60360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ordoñez, J.C.; van Bodegom, P.M.; Witte, J.-P.M.; Wright, I.J.; Reich, P.B.; Aerts, R. A global study of relationships between leaf traits, climate and soil measures of nutrient fertility. Glob. Ecol. Biogeogr. 2009, 18, 137–149. [Google Scholar] [CrossRef]
- Luo, Y.; Peng, Q.W.; He, M.S.; Zhang, M.X.; L iu, Y.Y.; Gong, Y.M.; Eziz, A.; Liu, K.H.; Han, W.X. N, P and K stoichiometry and resorption efficiency of nine dominant shrub species in the deserts of Xinjiang, China. Ecol. Res. 2020, 35, 625–637. [Google Scholar] [CrossRef]
- Sun, L.K.; Zhang, B.G.; Wang, B.; Zhang, G.S.; Zhang, W.; Zhang, B.L.; Chang, S.J.; Chen, T.; Liu, G.X. Leaf elemental stoi-chiometry of Tamarix Lour. species in relation to geographic, climatic, soil, and genetic components in China. Ecol. Eng. 2019, 106, 448–457. [Google Scholar] [CrossRef]
- Albert, C.H.; Thuiller, W.; Yoccoz, N.G.; Soudant, A.; Boucher, F.; Saccone, P.; Lavorel, S. Intraspecific functional variability: Extent, structure and sources of variation. J. Ecol. 2010, 98, 604–613. [Google Scholar] [CrossRef]
- Hu, Y.-K.; Zhang, Y.-L.; Liu, G.-F.; Pan, X.J.; Yang, X.; Li, W.B.; Dai, W.-H.; Tang, S.-L.; Xiao, T.; Chen, L.-Y.; et al. Intraspecific N and P stoichiometry of Phragmites australis: Geographic patterns and variation among climatic regions. Sci. Rep. 2017, 7, 43018. [Google Scholar] [CrossRef] [Green Version]
- Han, W.X.; Fang, J.Y.; Reich, P.B.; Woodward, F.I.; Wang, Z.H. Biogeography and variability of eleven mineral elements in plant leaves across gradients of climate, soil and plant functional type in China. Ecol. Lett. 2011, 14, 788–796. [Google Scholar] [CrossRef] [PubMed]
- Sardans, J.; Peñuelas, J. Potassium: A neglected nutrient in global change. Glob. Ecol. Biogeogr. 2015, 24, 261–275. [Google Scholar] [CrossRef] [Green Version]
- Koerselman, W.; Meuleman, A.F.M. The Vegetation N:P Ratio: A New Tool to Detect the Nature of Nutrient Limitation. J. Appl. Ecol. 1996, 33, 1441–1450. [Google Scholar] [CrossRef]
- Han, W.X.; Fang, J.Y.; Guo, D.L.; Zhang, Y. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China. New Phytol. 2005, 168, 377–385. [Google Scholar] [CrossRef]
- Song, Z.L.; Liu, H.Y.; Zhao, F.J.; Xu, C.Y. Ecological stoichiometry of N:P:Si in China’s grasslands. Plant Soil 2014, 380, 165–179. [Google Scholar] [CrossRef]
- Fan, J.W.; Harris, W.; Zhong, H.P. Stoichiometry of leaf nitrogen and phosphorus of grasslands of the Inner Mongolian and Qinghai-Tibet Plateaus in relation to climatic variables and vegetation organization levels. Ecol. Res. 2016, 31, 821–829. [Google Scholar] [CrossRef]
- Elser, J.J.; Bracken, M.E.S.; Cleland, E.E.; Gruner, D.S.; Harpole, W.S.; Hillebrand, H.; Ngai, J.T.; Seabloom, E.W.; Shurin, J.B.; Smith, J.E. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecol. Lett. 2007, 10, 1135–1142. [Google Scholar] [CrossRef] [Green Version]
- Shin, R. Strategies for improving potassium use efficiency in plants. Mol. Cells 2014, 37, 575–584. [Google Scholar] [CrossRef] [Green Version]
- Adams, E.; Shin, R. Transport, signaling, and homeostasis of potassium and sodium in plants. J. Integr. Plant Biol. 2014, 56, 231–249. [Google Scholar] [CrossRef] [PubMed]
- Venterink, H.O.; Wassen, M.J.; Verkroost, A.W.M.; De Ruiter, P.C. Species richness- productivity patterns differ between N-, P-, and K-limited wetlands. Ecology 2003, 84, 2191–2199. [Google Scholar] [CrossRef]
- Li, L.H.; Chen, J.Q.; Han, X.G.; Zhang, W.H.; Shao, C.L. Grassland Ecosystems of China: A Synthesis and Resume; Springer Nature: Singapore, 2020. [Google Scholar]
- Hong, J.T.; Wang, X.D.; Wu, J.B. Effects of soil fertility on the N:P stoichiometry of herbaceous plants on a nutrient-limited alpine steppe on the northern Tibetan Plateau. Plant Soil 2015, 391, 179–194. [Google Scholar] [CrossRef]
- Giordano, M. Homeostasis: An underestimated focal point of ecology and evolution. Plant Sci. 2013, 211, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.N.; Wang, J.Y.; Wang, L.; Zhang, H.; Guo, Z.W.; Wang, G.G.; Smith, W.K.; Wu, T.G. Does stoichiometric homeostasis differ among tree organs and with tree age? For. Ecol. Manag. 2019, 453, 117637. [Google Scholar] [CrossRef]
- Li, Y.F.; Li, Q.Y.; Guo, D.Y.; Liang, S.; Wang, Y.J. Ecological stoichiometry homeostasis of Leymus chinensis in degraded grassland in western Jilin Province, NE China. Ecol. Eng. 2016, 90, 387–391. [Google Scholar] [CrossRef]
- Ralphs, M.H. Ecological relationships between poisonous plants and rangeland condition: A review. Rangel. Ecol. Manag. 2002, 55, 285. [Google Scholar] [CrossRef]
- Kleijn, D.; Müller-Schärer, H. The Relation Between Unpalatable Species, Nutrients and Plant Species Richness in Swiss Montane Pastures. Biodivers. Conserv. 2006, 15, 3971–3982. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Sun, J.; Liu, M.; Xu, M.; Wang, Y.; Wu, G.L.; Zhou, H.; Ye, C.; Tsechoe, D.; Wei, T. Don’t judge toxic weeds on whether they are native but on their ecological effects. Ecol. Evol. 2020, 10, 9014–9025. [Google Scholar] [CrossRef]
- Bai, X.J.; Wang, B.R.; An, S.S.; Zeng, Q.C.; Zhang, H.X. Response of forest species to C:N:P in the plant-litter-soil system and stoichiometric homeostasis of plant tissues during afforestation on the Loess Plateau, China. CATENA 2019, 183, 104186. [Google Scholar] [CrossRef]
- Johnson, N.C.; Rowland, D.L.; Corkidi, L.; Allen, E.B. Plant winners and losers during grassland N-eutrophication differ in biomass allocation and mycorrhizas. Ecology 2008, 89, 2868–2878. [Google Scholar] [CrossRef] [Green Version]
- Aerts, R.; Chapin, F.S., III. The mineral nutrition of wild plants revisited: A reevaluation of processes and patterns. Adv. Ecol. Res. 2000, 30, 1–67. [Google Scholar]
- Tilman, D.; Reich, P.B.; Knops, J.M.H. Biodiversity and ecosystem stability in a decade-long grassland experiment. Nature 2006, 441, 629–632. [Google Scholar] [CrossRef] [PubMed]
- Berg, G.; Smalla, K. Plant species and soil type cooperatively shape the structure and function of microbial communities in the rhizosphere. FEMS Microbiol. Ecol. 2009, 68, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manrubia, M.; van der Putten, W.; Weser, C.; Veen, C. Rhizosphere and litter feedbacks to range-expanding plant species and related natives. J. Ecol. 2019, 108, 353–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.; Bowatte, S.; Wang, Y.; Newton, P.; Hou, F. Differences in soil ammonia oxidizing bacterial communities under unpalatable (Stellera chamaejasme L.) and palatable (Elymus nutans Griseb.) plants growing on the Qinghai Tibetan Plateau. Soil Biol. Biochem. 2020, 144, 107779. [Google Scholar] [CrossRef]
- Wardle, D.A.; Bardgett, R.D.; Klironomos, J.N.; Setälä, H.; van der Putten, W.H.; Wall, D.H. Ecological Linkages between Aboveground and Belowground Biota. Science 2004, 304, 1629–1633. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Wardle, D.A. Aboveground-Belowground Linkages: Biotic Interactions, Ecosystem Processes, and Global Change; Oxford University Press: Oxford, UK, 2010. [Google Scholar]
- Zhang, P.; Li, B.; Wu, J.H.; Hu, S.J. Invasive plants differentially affect soil biota through litter and rhizosphere pathways: A meta-analysis. Ecol. Lett. 2018, 22, 200–210. [Google Scholar] [CrossRef] [Green Version]
- Pringle, A.; Bever, J.D.; Gardes, M.; Parrent, J.L.; Rillig, M.C.; Klironomos, J.N. Mycorrhizal symbioses and plant invasions. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 699–715. [Google Scholar] [CrossRef] [Green Version]
- Vogelsang, K.M.; Bever, J.D. Mycorrhizal densities decline in association with nonnative plants and contribute to plant in-vasion. Ecology 2009, 90, 399–407. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.J.; Li, Q.H.; Mao, W.Q.; Tang, W.T.; White, J.F., Jr.; Li, H.Y. Endophytic bacterial community of Stellera chamaejasme L. and its role in improving host plants’ competitiveness in grasslands. Environ. Microbiol. 2022. peer-review. [Google Scholar] [CrossRef]
- He, W.; Detheridge, A.; Liu, Y.M.; Wang, L.; Wei, H.C.; Griffith, G.W.; Scullion, J.; Wei, Y.H. Variation in soil fungal composition associated with the invasion of Stellera chamaejasme L. in Qinghai-Tibet Plateau grassland. Microorganisms 2019, 7, 587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivas-Ubach, A.; Sardans, J.; Pérez-Trujillo, M.; Estiarte, M.; Peñuelas, J. Strong relationship between elemental stoichiometry and metabolome in plants. Proc. Natl. Acad. Sci. USA 2012, 109, 4181–4186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sardans, J.; Peñuelas, J.; Coll, M.; Vayreda, J.; Rivas-Ubach, A. Stoichiometry of potassium is largely determined by water availability and growth in Catalonian forests. Funct. Ecol. 2012, 26, 1077–1089. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Peng, Q.W.; Li, K.H.; Gong, Y.M.; Liu, Y.Y.; Han, W.X. Patterns of nitrogen and phosphorus stoichiometry among leaf, stem and root of desert plants and responses to climate and soil factors in Xinjiang, China. CATENA 2021, 199, 105100. [Google Scholar] [CrossRef]
- Kopáček, J.; Turek, J.; Hejzlar, J.; Šantrůčková, H. Canopy leaching of nutrients and metals in a mountain spruce forest. Atmospheric Environ. 2009, 43, 5443–5453. [Google Scholar] [CrossRef]
- Chen, J.H.; Yan, F.; Lu, Q. Spatiotem-poral variation of vegetation on the Qinghai-Tibet plateau and the influence of climatic factors and human activities on vegetation trend (2000–2019). Remote Sens. 2020, 12, 3150. [Google Scholar] [CrossRef]
- Wei, Y.Q.; Lu, H.Y.; Wang, J.N.; Sun, J.; Wang, X.F. Responses of vegetation zones, in the Qinghai-Tibetan Plateau, to climate change and anthropogenic influences over the last 35 years. Pratacultural Sci. 2019, 36, 250–263. (In Chinese) [Google Scholar]
- Bao, S.D. Soil Agricultural Chemical Analysis, 3rd ed.; China Agricultural Press: Beijing, China, 2000. (In Chinese) [Google Scholar]
- Warton, D.I.; Duursma, R.A.; Falster, D.S.; Taskinen, S. SMATR 3—An R package for estimation and inference about allometric lines. Methods Ecol. Evol. 2011, 3, 257–259. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| All (n = 29) | QT (n = 19) | IM (n = 10) | ||||
|---|---|---|---|---|---|---|
| Mean ± SD | CV (%) | Mean ± SD | CV (%) | Mean ± SD | CV (%) | |
| Carbon (g kg−1) | 498.60 ± 22.07 | 4.43 | 498.97 ± 21.14 | 4.24 | 497.89 ± 24.91 | 5.00 |
| Nitrogen (g kg−1) | 19.95 ± 2.09 | 10.47 | 19.94 ± 2.02 | 10.15 | 19.97 ± 2.32 | 11.63 |
| Phosphorus (g kg−1) | 2.15 ± 0.52 | 24.33 | 2.21 ± 0.53 | 24.15 | 2.05 ± 0.52 | 25.20 |
| Potassium (g kg−1) | 6.57 ± 1.18 | 17.94 | 7.13 ± 0.99 a | 13.94 | 5.51 ± 0.66 b | 12.02 |
| C:N | 25.20 ± 2.27 | 8.99 | 25.21 ± 2.18 | 8.65 | 25.18 ± 2.543 | 10.09 |
| C:P | 245.57 ± 61.64 | 25.10 | 239.45 ± 61.85 | 25.83 | 257.20 ± 62.77 | 24.41 |
| N:P | 9.81 ± 2.60 | 26.54 | 9.63 ± 2.89 | 30.05 | 10.15 ± 2.04 | 20.05 |
| N:K | 3.13 ± 0.63 | 20.10 | 2.84 ± 0.45 b | 15.70 | 3.67 ± 0.58 a | 15.83 |
| K:P | 3.21 ± 0.93 | 28.89 | 3.42 ± 1.02 a | 29.76 | 2.81 ± 0.57 b | 20.44 |
| All (n = 29) | QT (n = 19) | IM (n = 10) | ||||
|---|---|---|---|---|---|---|
| Mean ± SD | CV (%) | Mean ± SD | CV (%) | Mean ± SD | CV (%) | |
| Carbon (g kg−1) | 46.11 ± 21.63 | 46.90 | 48.65 ± 18.31 | 37.63 | 41.29 ± 27.30 | 66.11 |
| Nitrogen (g kg−1) | 3.75 ± 1.70 | 45.24 | 3.93 ± 1.68 | 42.70 | 3.41 ± 1.77 | 51.85 |
| Phosphorus (g kg−1) | 0.57 ± 0.17 | 29.24 | 0.61 ± 0.13 a | 22.04 | 0.49 ± 0.20 b | 40.88 |
| Potassium (g kg−1) | 20.80 ± 5.86 | 28.17 | 22.23 ± 5.00 a | 22.49 | 18.09 ± 6.66 b | 36.82 |
| C:N | 13.54 ± 6.72 | 49.62 | 14.80 ± 7.89 | 53.30 | 11.16 ± 2.54 | 22.91 |
| C:P | 77.72 ± 26.00 | 33.46 | 79.31 ± 25.53 | 32.19 | 74.69 ± 28.01 | 36.06 |
| N:P | 6.34 ± 2.16 | 34.13 | 6.25 ± 2.44 | 39.10 | 6.52 ± 1.61 | 25.58 |
| N:P | 0.20 ± 0.14 | 68.81 | 0.18 ± 0.08 | 42.44 | 0.25 ± 0.21 | 85.03 |
| K:P | 38.73 ± 15.12 | 39.04 | 38.68 ± 13.27 | 34.31 | 38.83 ± 18.95 | 48.81 |
| Available phosphorus (mg kg−1) | 5.29 ± 1.96 | 37.07 | 5.84 ± 1.90 a | 32.57 | 4.25 ± 1.70 b | 40.09 |
| Available potassium (mg kg−1) | 175.91 ± 96.39 | 54.79 | 176.69 ± 106.27 | 60.14 | 174.43 ± 79.47 | 45.56 |
| Ammonium nitrogen (mg kg−1) | 19.17 ± 7.89 | 41.14 | 19.05 ± 7.64 | 40.12 | 19.39 ± 8.75 | 45.11 |
| Nitrate nitrogen (mg kg−1) | 14.12 ± 14.20 | 100.59 | 12.95 ± 4.46 | 34.42 | 16.35 ± 24.07 | 147.25 |
| Water content | 0.18 ± 0.08 | 44.51 | 0.21 ± 0.08 a | 36.46 | 0.14 ± 0.07 b | 53.50 |
| pH | 7.90 ± 0.51 | 6.41 | 8.05 ± 0.39 a | 4.90 | 7.63 ± 0.60 b | 7.89 |
| Electrical conductivity (μs cm−1) | 247.21 ± 221.21 | 89.48 | 193.96 ± 74.42 b | 38.37 | 348.38 ± 351.86 a | 101.00 |
| Y | X | 1/H (slope) | p | r2 | Category |
|---|---|---|---|---|---|
| Leaf C | Soil C | 0 | 0.351 | 0.0323 | strictly homeostatic |
| Leaf N | Soil N | 0 | 0.829 | 0.0017 | strictly homeostatic |
| Soil ammonium N | −0.273 | 0.089 | 0.1033 | weakly homeostatic | |
| Soil nitrate N | 0 | 0.291 | 0.0412 | strictly homeostatic | |
| Leaf P | Soil P | 0.660 | 0.042 | 0.1437 | weakly plastic |
| Soil available P | 0.622 | 0.002 | 0.2968 | weakly plastic | |
| Leaf K | Soil K | 0 | 0.112 | 0.0910 | strictly homeostatic |
| Soil available K | 0 | 0.154 | 0.0738 | strictly homeostatic | |
| Leaf C:N | Soil C:N | 0 | 0.789 | 0.0028 | strictly homeostatic |
| Leaf C:P | Soil C:P | −0.622 | 0.018 | 0.1915 | weakly plastic |
| Leaf N:P | Soil N:P | −0.474 | 0.085 | 0.1061 | weakly homeostatic |
| Leaf N:K | Soil N:K | 0 | 0.956 | 0.0001 | strictly homeostatic |
| Leaf K:P | Soil K:P | 0 | 0.774 | 0.0031 | strictly homeostatic |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, L.; Liu, L.; Meng, H.; Zhang, L.; Silva, V.J.; Zhao, H.; Wang, K.; He, W.; Huang, D. Biogeographic Patterns of Leaf Element Stoichiometry of Stellera chamaejasme L. in Degraded Grasslands on Inner Mongolia Plateau and Qinghai-Tibetan Plateau. Plants 2022, 11, 1943. https://doi.org/10.3390/plants11151943
Guo L, Liu L, Meng H, Zhang L, Silva VJ, Zhao H, Wang K, He W, Huang D. Biogeographic Patterns of Leaf Element Stoichiometry of Stellera chamaejasme L. in Degraded Grasslands on Inner Mongolia Plateau and Qinghai-Tibetan Plateau. Plants. 2022; 11(15):1943. https://doi.org/10.3390/plants11151943
Chicago/Turabian StyleGuo, Lizhu, Li Liu, Huizhen Meng, Li Zhang, Valdson José Silva, Huan Zhao, Kun Wang, Wei He, and Ding Huang. 2022. "Biogeographic Patterns of Leaf Element Stoichiometry of Stellera chamaejasme L. in Degraded Grasslands on Inner Mongolia Plateau and Qinghai-Tibetan Plateau" Plants 11, no. 15: 1943. https://doi.org/10.3390/plants11151943