
Effects of Iron Oxide Nanoparticles (Fe3O4) on Growth, Photosynthesis, Antioxidant Activity and Distribution of Mineral Elements in Wheat (Triticum aestivum) Plants

 , and
, and
Abstract
:1. Introduction
2. Results
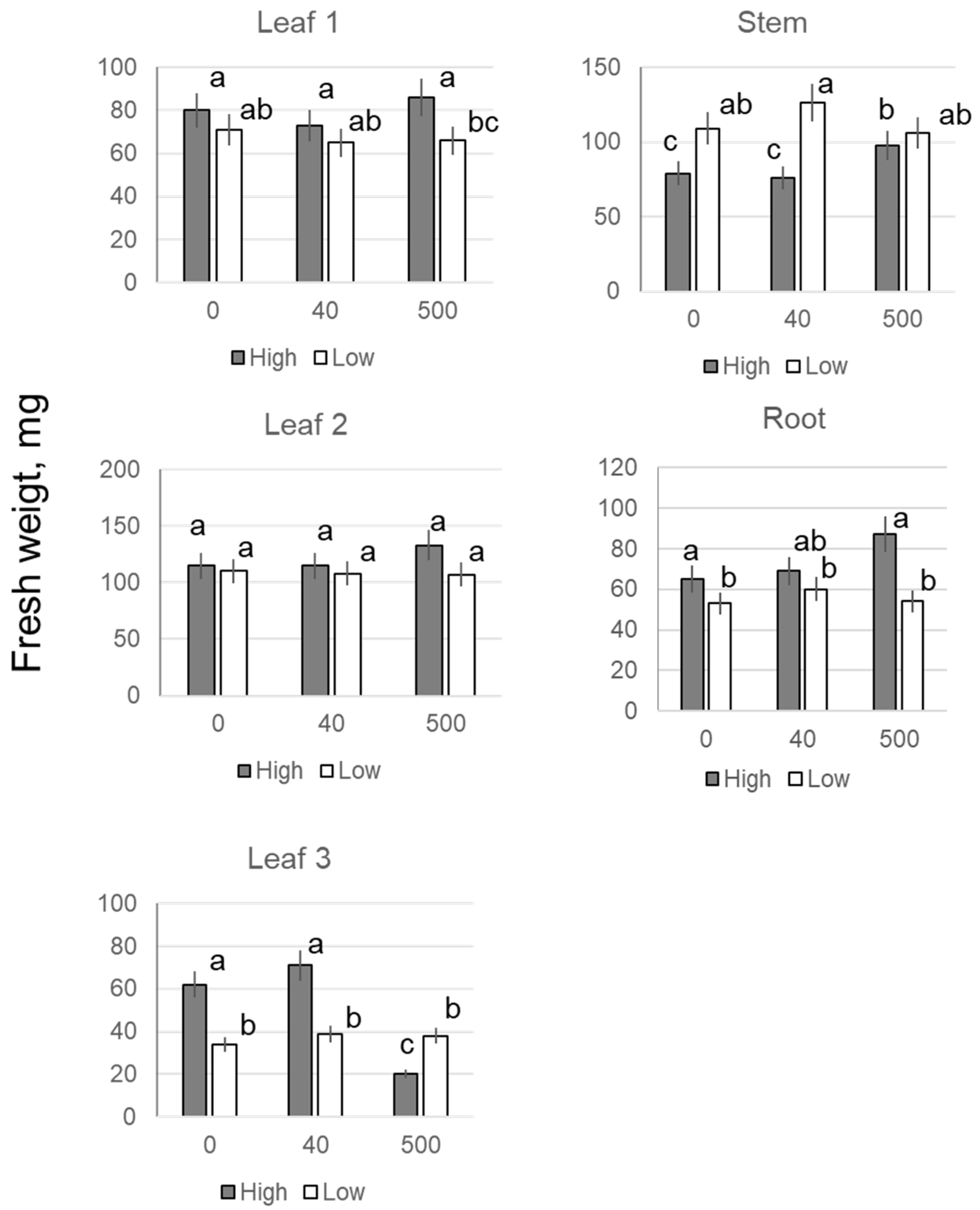
2.1. Growth
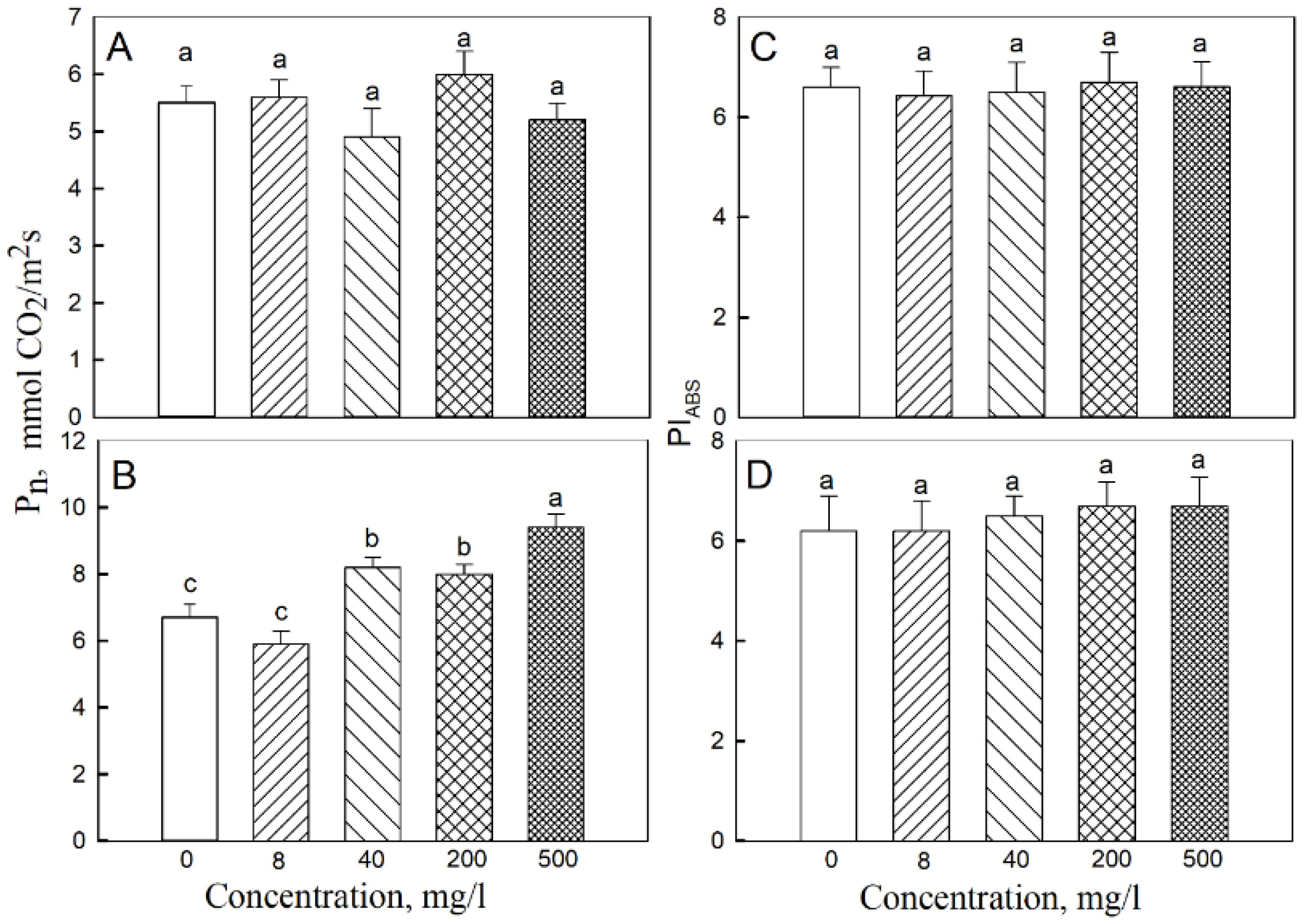
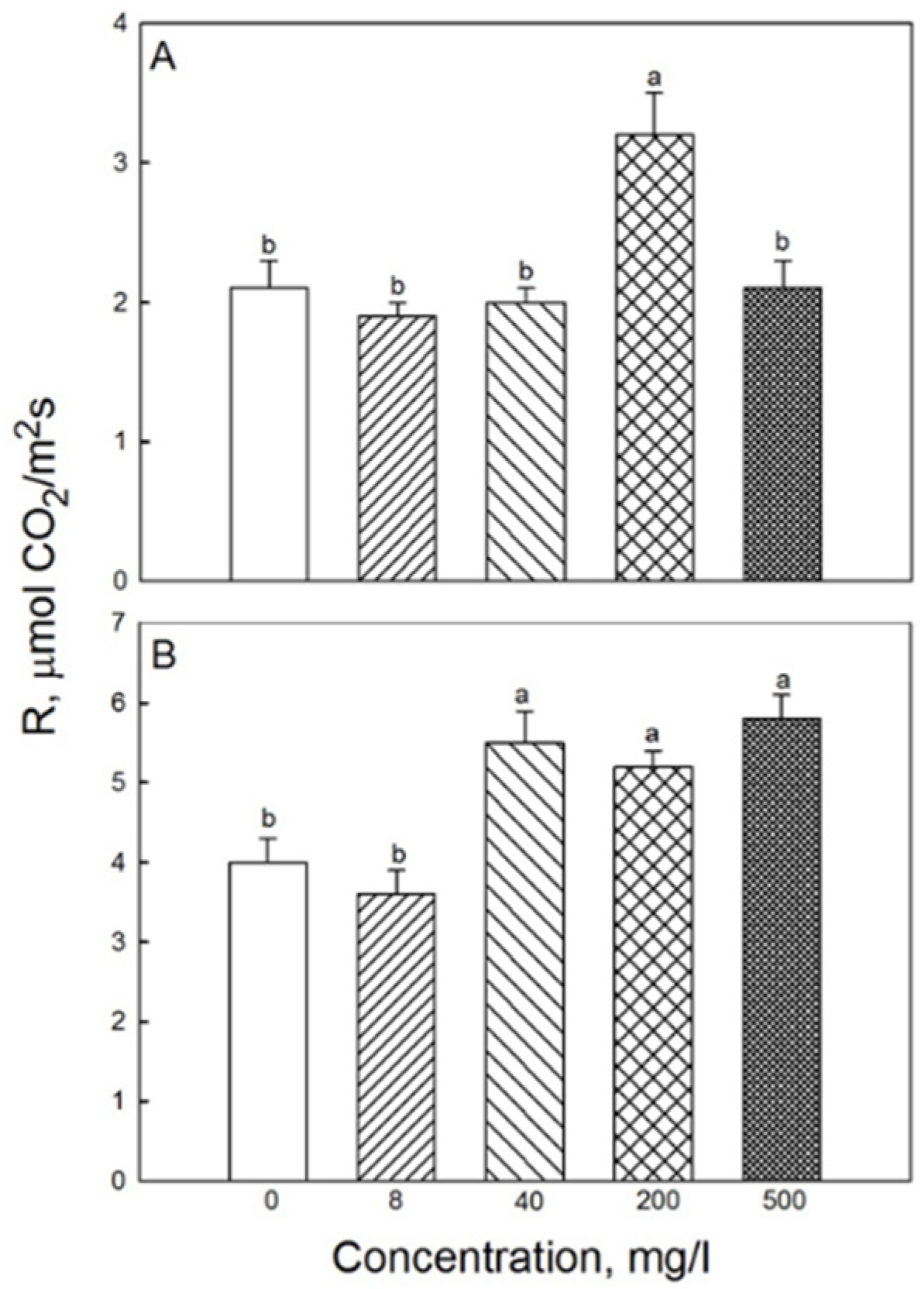
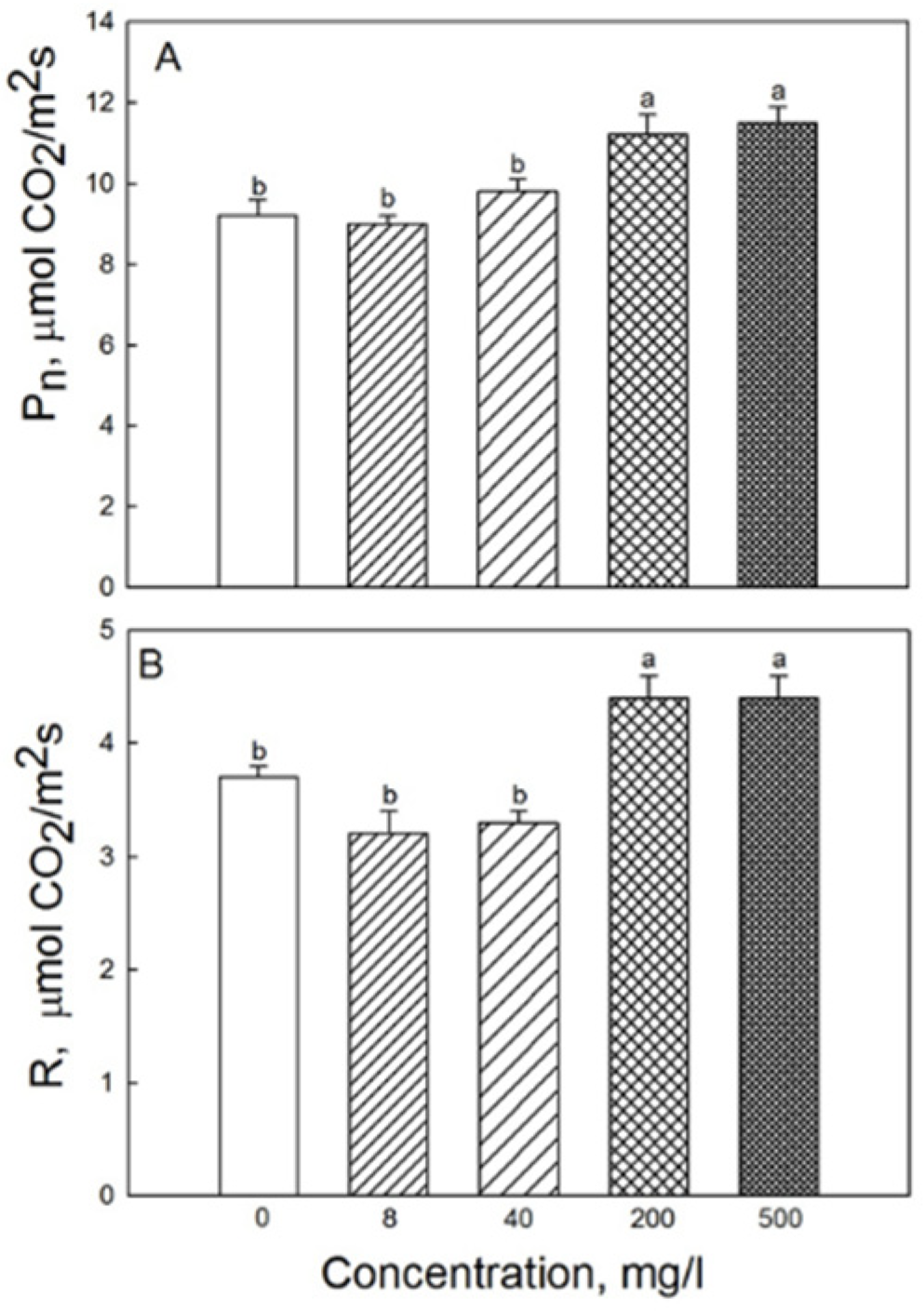
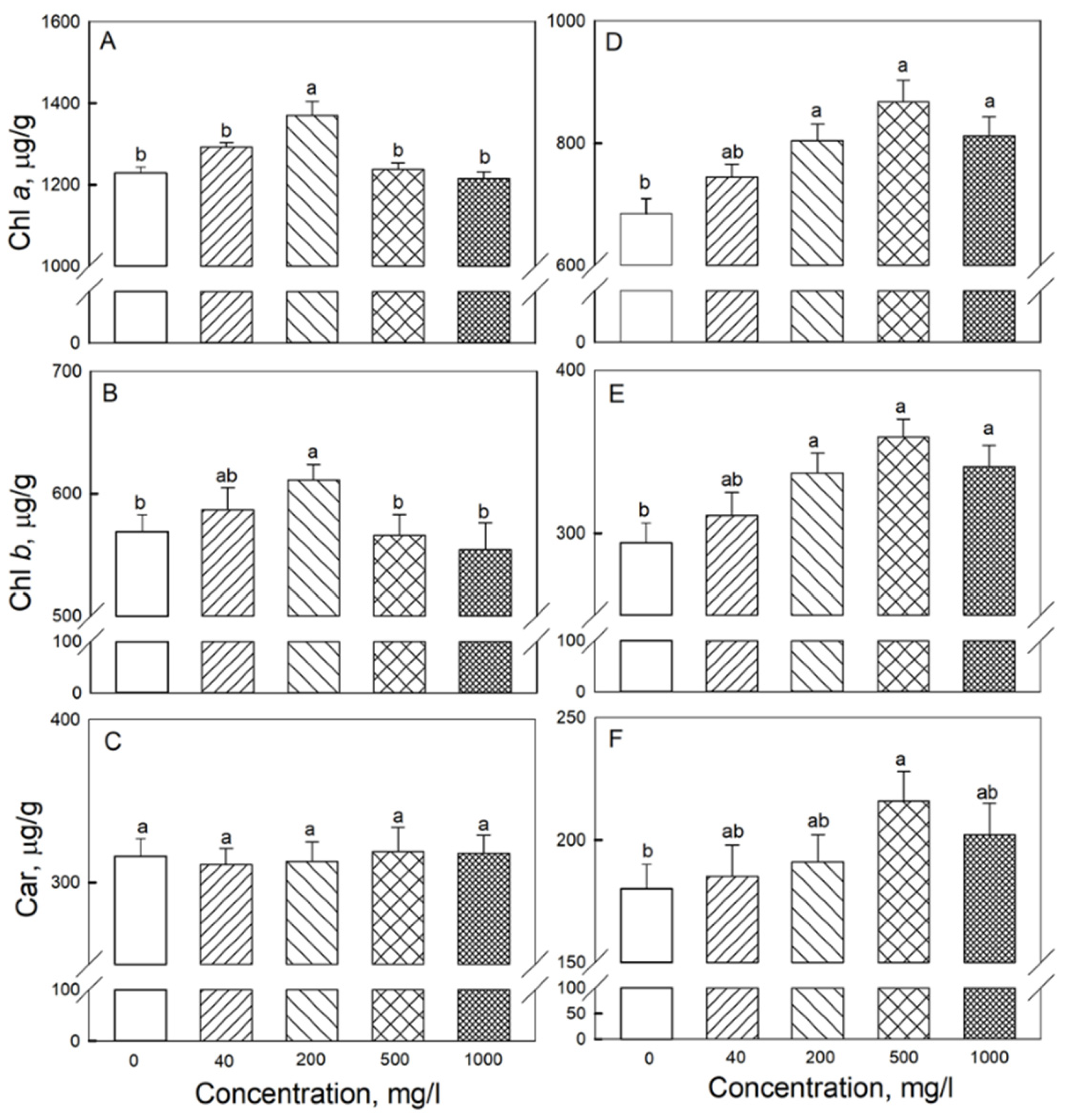
2.2. Photosynthesis and Respiration
2.3. Photochemical Activity
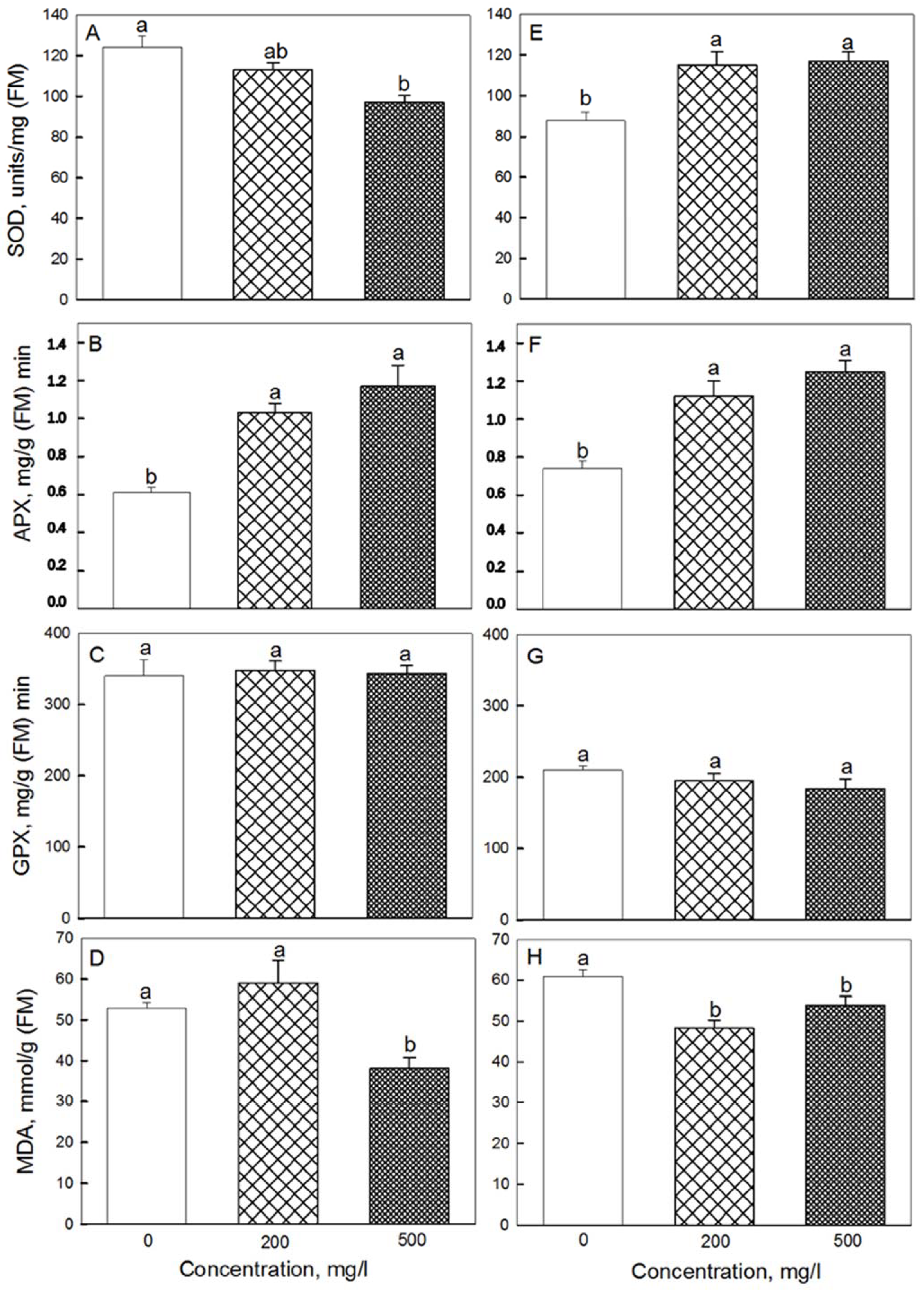
2.4. Antioxidant Activity
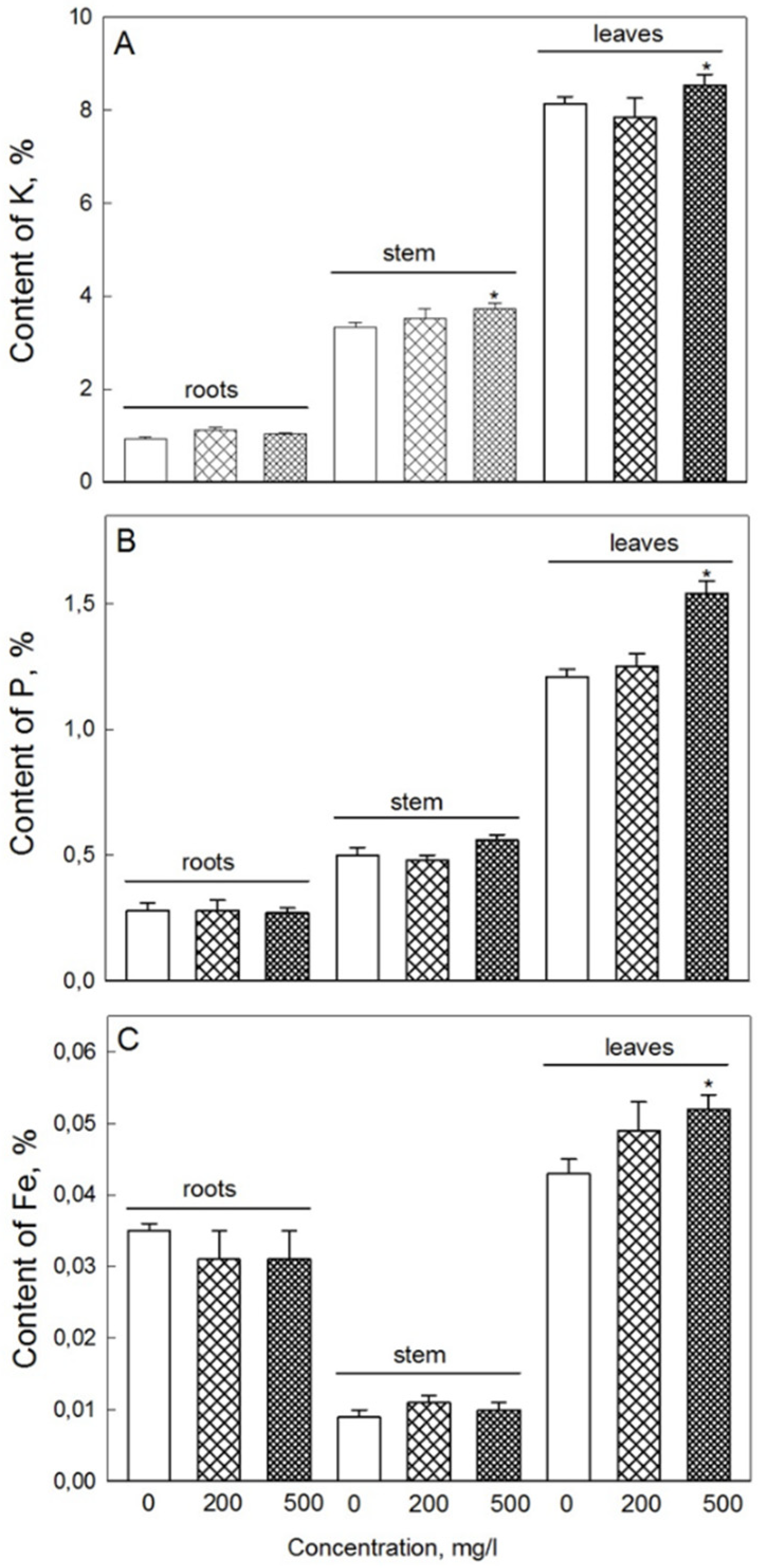
2.5. Elemental Content
3. Discussion
4. Materials and Methods
4.1. Growth and Treatments Conditions
4.2. Agronomical Characteristics
4.3. Pigment Contents
4.4. Gas Exchange Characteristics
4.5. Determination of Photochemical Activity
4.6. Lipid Peroxidation
4.7. Plant Elemental Analysis
4.8. Superoxide Dismutase Activity
4.9. Ascorbate Peroxidase Activity
4.10. Guaiacol-Dependent Peroxidase Activity
4.11. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, R.; Lal, R. Potentials of engineered nanoparticles as fertilizers for increasing agronomic productions. Sci. Total Environ. 2015, 514, 131–139. [Google Scholar] [CrossRef]
- Farooqui, A.; Tabassum, H.; Ahmad, A.; Mabood, A.; Ahmad, A.; Ahmad, I.Z. Role of nanoparticles in growth and development of plants: A review. Int. J. Pharm. 2016, 7, 22–37. [Google Scholar]
- Huang, S.; Wang, L.; Liu, L.; Hou, Y.; Lu, L. Nanotechnology in agriculture, livestock, and aquaculture in China. A review. Agron. Sustain. Dev. 2015, 35, 369–400. [Google Scholar] [CrossRef] [Green Version]
- Siddiqi, K.S.; Rahman, A.; Tajuddin, H.A. Biogenic fabrication of iron/iron oxide nanoparticles and their application. Nanoscale Res. Lett. 2016, 11, 498. [Google Scholar] [CrossRef] [Green Version]
- Rui, M.; Ma, C.; Hao, Y.; Guo, J.; Rui, Y.; Tang, X.; Zhao, Q.; Fan, X.; Zhang, Z.; Hou, T.; et al. Iron oxide nanoparticles as a potential iron fertilizer for peanut (Arachis hypogaea). Front. Plant Sci. 2016, 7, 815. [Google Scholar] [CrossRef] [Green Version]
- Hong, F.; Zhou, J.; Liu, C.; Yang, F.; Wu, C.; Zheng, L.; Yang, P. Effect of Nano-TiO2 on photochemical reaction of chloroplasts of spinach. Biol. Trace Elem. Res. 2005, 105, 269–279. [Google Scholar] [CrossRef]
- Boghossian, A.A.; Sen, F.; Gibbons, B.M.; Sen, S.; Faltermeier, S.M.; Giraldo, J.P.; Zhang, C.T.; Zhang, J.Q.; Heller, D.A.; Strano, M.S. Application of nanoparticle antioxidants to enable hyperstable chloroplasts for solar energy harvesting. Adv. Energy Mater. 2013, 3, 881–893. [Google Scholar] [CrossRef]
- Djanaguiraman, M.; Nair, R.; Giraldo, J.P.; Prasad, P. Cerium oxide nanoparticles decrease drought-induced oxidative damage in sorghum leading to higher photosynthesis and grain yield. ACS Omega 2018, 3, 14406–14416. [Google Scholar] [CrossRef]
- Duran, N.M.; Savassa, S.M.; de Lima, R.G.; de Almeida, E. X-ray spectroscopy uncovering the effects of Cu based nanoparticle concentration and structure on Phaseolus vulgaris germination and seedling development. J. Agric. Food Chem. 2017, 65, 7874–7884. [Google Scholar] [CrossRef]
- Rashad, Y.M.; El-Sharkawy, H.H.A.; Belal, B.E.A.; Razik, E.A.S.; Galilah, D.A. Silica nanoparticles as a probable anti-oomycete compound against downy mildew, and yield and quality enhancer in grapevines: Field evaluation, molecular, physiological, ultrastructural, and toxicity investigations. Front. Plant Sci. 2021, 12, 763365. [Google Scholar] [CrossRef]
- Bakhtiari, M.; Moaveni, P.; Sani, B. The effect of iron nanoparticles spraying time and concentration on wheat. Biol. Forum-Int. J. 2015, 7, 679–683. [Google Scholar]
- Shaw, A.K.; Hossain, Z. Impact of nano-CuO stress on rice (Oryza sativa L.) seedlings. Chemosphere 2013, 93, 906–915. [Google Scholar] [CrossRef]
- Asli, S.; Neumann, P.M. Colloidal suspensions of clay or titanium dioxide nanoparticles can inhibit leaf growth and transpiration via physical effects on root water transport. Plant Cell Environ. 2009, 32, 577–584. [Google Scholar] [CrossRef]
- Lin, D.; Xing, B. Phytotoxicity of nanoparticles: Inhibition of seed germination and root growth. Environ. Polluttion 2007, 150, 243–250. [Google Scholar] [CrossRef]
- Yusefi-Tanha, E.; Fallah, S.; Rostamnejadi, A.; Pokhrel, L.R. Root system architecture, copper uptake and tissue distribution in soybean (Glycine max (L.) Merr.) grown in copper oxide nanoparticle (CuO NP)-amended soil and implications for human nutrition. Plants 2020, 9, 1326. [Google Scholar] [CrossRef]
- Yusefi-Tanha, E.; Fallah, S.; Rostamnejadi, A.; Pokhrel, L.R. Zinc oxide nanoparticles (ZnO NPs) as a novel nanofertilizer: Influence on seed yield and antioxidant defense system in soil grown soybean (Glycine max cv. Kowsar). Sci. Total Environ. 2020, 738, 140240. [Google Scholar] [CrossRef]
- Hossain, Z.; Yasmeen, F.; Komatsu, S. Nanoparticles: Synthesis, morphophysiological effects, and proteomic responses of crop plants. Int. J. Mol. Sci. 2020, 21, 3056. [Google Scholar] [CrossRef]
- Taran, S.; Garip, A.K.; Arslan, H. Theoretical study of the structures and chemical ordering of CoPd nanoalloys supported on MgO(001). Int. J. Mod. Phys. C 2016, 27, 1650146. [Google Scholar] [CrossRef]
- Dimkpa, C.O.; Latta, D.E.; Mclean, J.E.; Britt, D.W.; Boyanov, M.I.; Anderson, A.J. Fate of CuO and ZnO nano- and microparticles in the plant environment. Environ. Sci. Technol. 2013, 47, 4734–4742. [Google Scholar] [CrossRef]
- Angel, B.M.; Batley, G.E.; Jarolimek, C.V.; Rogers, N.J. The impact of size on the fate and toxicity of nanoparticulate silver in aquatic systems. Chemosphere 2013, 93, 359–365. [Google Scholar] [CrossRef]
- Zhu, H.; Han, J.; Xiao, J.Q.; Jin, Y. Uptake, translocation, and accumulation of manufactured iron oxide nanoparticles by pumpkin. J. Environ. Monit. 2008, 10, 713–717. [Google Scholar] [CrossRef]
- Junior, J.; Prado, R.; Soares, M.B.; Silva, J.; Cazetta, J.O. Effect of different foliar silicon sources on cotton plants. J. Soil Sci. Plant Nutr. 2020, 21, 95–103. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, N.; Wang, W.; Sun, J.; Zhu, L. Photosynthesis and related metabolic mechanism of promoted rice (Oryza sativa L.) growth by TiO2 nanoparticles. Front. Environ. Sci. Eng. 2020, 14, 12. [Google Scholar] [CrossRef]
- Rossi, L.; Zhang, W.L.; Lombardini, L.; Ma, X.M. The impact of cerium oxide nanoparticles on the salt stress responses of Brassica napus L. Environ. Pollut. 2016, 219, 28–36. [Google Scholar] [CrossRef] [Green Version]
- Bozorgi, H.R. Study effects of nitrogen fertilizer management under nano iron chelate foliar spraying on yield and yield components of eggplant (Solanum melongena L.). J. Agric. Biol. Sci. 2012, 7, 233–237. [Google Scholar]
- Alidoust, D.; Isoda, A. Effect of gamma Fe2O3 nanoparticles on photosynthetic characteristic of soybean (Glycine max (L) Merr.): Foliar spray versus soil amendment. Acta Physiol. Plant. 2013, 35, 3365–3375. [Google Scholar] [CrossRef]
- Ghasemi, S.; Khoshgoftarmanesh, A.H.; Afyuni, M.; Hadadzadeh, H. Iron(II)–amino acid chelates alleviate damages salt-stress induced oxidative on tomato grown in nutrient solution culture. Sci. Hortic. 2014, 165, 91–98. [Google Scholar] [CrossRef]
- Winder, T.L.; Nishio, J.N. Early iron deficiency stress response in leaves of sugar beet. Plant Physiol. 1995, 108, 1487–1494. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S. Adaptation of photosynthesis under iron deficiency in maize. J. Plant Physiol. 2007, 164, 1261–1267. [Google Scholar] [CrossRef]
- Sharma, P.N.; Tripathi, A.; Kumar, N.; Gupta, S.; Kumar, P.; Chatterjee, J.; Tewari, R.K. Iron plays a critical role in stomatal closure in cauliflower. Environ. Exp. Bot. 2016, 131, 68–76. [Google Scholar] [CrossRef]
- Alkhatib, R.; Alkhatib, B.; Abdo, N.; AL-Eitan, L.; Creamer, R. Physio-biochemical and ultrastructural impact of (Fe3O4) nanoparticles on tobacco. BMC Plant Biol. 2019, 19, 253. [Google Scholar] [CrossRef] [Green Version]
- Mazaherinia, S.; Astaraei, A.; Fotovvat, A.; Monshi, A. Nano iron oxide particles efficiency on fe, mn, zn and cu concentrations in wheat plant. World Appl. Sci. J. 2010, 3, 36–40. [Google Scholar]
- Poddar, K.; Sarkar, D.; Sarkar, A. Nanoparticles on Photosynthesis of Plants: Effects and Role. Green Nanoparticles 2020, 7, 273–287. [Google Scholar] [CrossRef]
- Siddiqi, K.S.; Husen, A. Plant response to strigolactones: Current developments and emerging trends. Appl. Soil Ecol. 2017, 120, 247–253. [Google Scholar] [CrossRef]
- Sundaria, N.; Singh, M.; Upreti, P.; Chauhan, R.P.; Jaiswal, J.P.; Kumar, A. Seed priming with iron oxide nanoparticles triggers iron acquisition and biofortification in wheat (triticum aestivum L.) grains. J. Plant Growth Regul. 2018, 38, 122–131. [Google Scholar] [CrossRef]
- Marschner, H. Mineral Nutrition of Higher Plants, 2nd ed.; Academie Press: London, UK, 1995. [Google Scholar]
- Wu, L.B.; Ueda, Y.; Lai, S.K.; Frei, M. Shoot tolerance mechanisms to iron toxicity in rice (Oryza sativa L.). Plant Cell Environ. 2016, 40, 570–584. [Google Scholar] [CrossRef]
- Nel, A.; Xia, T.; Mädler, L.; Li, N. Toxic potential of materials at the nanolevel. Science 2006, 311, 622–627. [Google Scholar] [CrossRef] [Green Version]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Rizwan, M.; Ali, S.; Ali, B.; Adrees, M.; Arshad, M.; Hussain, A.; Ziaur Rehman, M.; Waris, A.A. Zinc and Iron Oxide Nanoparticles Improved the Plant Growth and Reduced the Oxidative Stress and Cadmium Concentration in Wheat. Chemosphere 2019, 214, 269–277. [Google Scholar] [CrossRef]
- Shabala, S.; Pottosin, I.I. Regulation of potassium transport in plants under hostile conditions: Implications for abiotic and biotic stress tolerance. Physiol. Plant. 2014, 151, 257–279. [Google Scholar] [CrossRef]
- Goltsev, V.N.; Kalaji, H.M.; Paunov, M.; Bąba, W.; Horaczek, T.; Mojski, J.; Allakhverdiev, S.I. Variable chlorophyll fluorescence and its use for assessing physiological condition of plant photosynthetic apparatus. Russ. J. Plant Physiol. 2016, 63, 869–893. [Google Scholar] [CrossRef]
- Hanikenne, M.; Bernal, M.; Urzica, E.I. Ion homeostasis in the chloroplast. Biology 2014, 5, 465–514. [Google Scholar] [CrossRef] [Green Version]
- Yasmeen, F.; Raja, N.I.; Razzaq, A.; Komatsu, S. Proteomic and Physiological Analyses of Wheat Seeds Exposed to Copper and Iron Nanoparticles. Biochim. Biophys. Acta BBA Proteins Proteom. 2017, 1865, 28–42. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Wellburn, A.R. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar] [CrossRef]
- Kreslavski, V.D.; Lankin, A.V.; Vasilyeva, G.K.; Lyubimov, V.Y.; Semenova, G.N.; Schmitt, F.J.; Friedrich, T.; Allakhverdiev, S.I. Effects of polyaromatic hydrocarbons on photosystem II activity in pea leaves. Plant Physiol. Biochem. 2014, 81, 135–142. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Schansker, G.; Ladle, R.J.; Goltsev, V.; Bosa, K.; Allakhverdiev, S.I.; Brestic, M.; Bussotti, F.; Calatayud, A.; Dąbrowski, P.; et al. Frequently asked questions about in vivo chlorophyll fluorescence: Practical issues. Photosynth. Res. 2014, 122, 121–158. [Google Scholar] [CrossRef] [Green Version]
- Uchiyama, M.; Mihara, M. Determination of malonaldehyde precursor in tissues by thiobarbituric acid test. Anal. Biochem. 1978, 86, 287–297. [Google Scholar] [CrossRef]
- Towett, E.K.; Shepherd, K.D.; Cadisch, G. Quantification of total element concentrations in soils using total X-ray fluorescence spectroscopy (txrf). Sci. Total Environ. 2013, 463, 374–388. [Google Scholar] [CrossRef] [Green Version]
- Gupta, A.S.; Webb, R.P.; Holaday, A.S.; Allen, R.D. Overexpression of superoxide dismutase protects plants from oxidative stress. Plant Physiol. 1993, 103, 1067–1073. [Google Scholar] [CrossRef] [Green Version]
- Veljovic-Jovanovic, S.; Kukavica, B.; Stevanovic, B.; Navari-Izzo, F. Senescence- and drought-related changes in peroxidase and superoxide dismutase isoforms in leaves of Ramonda serbica. J. Exp. Bot. 2006, 57, 1759–1768. [Google Scholar] [CrossRef]
- Chance, B.; Maehly, A.C. Assay of catalase and peroxidases. Methods Enzymol. 1955, 2, 764–775. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Seedling Age | [Fe3O4], mg·L−1 | Biomass, mg | ||||||
|---|---|---|---|---|---|---|---|---|
| 1st Leaf | 2nd Leaf | 3rd Leaf | Stem | SL | Roots | 4th Leaf | ||
| 13-d-old | 0 | 75 ± 6 | 99 ± 10 | 4.3 ± 2.1 | 69 ± 6 | 248 ± 23 | - | - |
| 40 | 71 ± 8 | 101 ± 9 | 3.7 ± 1.8 | 65 ± 8 | 241 ± 22 | - | - | |
| 200 | 77 ± 7 | 98 ± 11 | 3.1 ± 1.5 | 62 ± 7 | 220 ± 20 | - | - | |
| 500 | 83 ± 6 | 91 ± 8 | 8.3 ± 2.4 | 72 ± 8 | 254 ± 19 | - | - | |
| 19-d-old | 0 | 80 ± 7 | 115 ± 10 | 62 ± 7c | 79 ± 6 | 338 ± 28 | 66 ± 5 | - |
| 40 | 73 ± 4 | 115 ± 10 | 71 ± 8bc | 76 ± 6 | 337 ± 31 | 69 ± 7 | - | |
| 200 | 79 ± 5 | 129 ± 8 | 87 ± 6 * | 94 ± 11 * | 388 ± 30 | 85 ± 6 * | - | |
| 500 | 86 ± 6 | 133 ± 15 | 120 ± 11 ** | 98 ± 10 * | 437 ± 35 * | 87 ± 7 * | - | |
| 1000 | 87 ± 7 | 130 ± 12 | 122 ± 9 ** | 96 ± 7 * | 441 ± 33 * | 88 ± 8 * | - | |
| 23-d-old | 0 | 71 ± 5 | 97 ± 10 | 148 ± 13 | 92 ± 8 | 411 ± 35 | 79 ± 6 | 3 ± 1 |
| 40 | 69 ± 4 | 94 ± 8 | 156 ± 17 | 89 ± 7 | 434 ± 38 | 80 ± 7 | 26 ± 3 | |
| 200 | 73 ± 7 | 105 ±11 | 174 ± 15 | 95 ± 8 | 467 ± 44 | 93 ± 5 | 20 ± 3 | |
| 500 | 71 ± 6 | 114 ± 9 | 168 ± 12 | 96 ± 6 | 482 ± 37 | 98 ± 6 * | 32 ± 7 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, Y.; Kreslavski, V.D.; Shmarev, A.N.; Ivanov, A.A.; Zharmukhamedov, S.K.; Kosobryukhov, A.; Yu, M.; Allakhverdiev, S.I.; Shabala, S. Effects of Iron Oxide Nanoparticles (Fe3O4) on Growth, Photosynthesis, Antioxidant Activity and Distribution of Mineral Elements in Wheat (Triticum aestivum) Plants. Plants 2022, 11, 1894. https://doi.org/10.3390/plants11141894
Feng Y, Kreslavski VD, Shmarev AN, Ivanov AA, Zharmukhamedov SK, Kosobryukhov A, Yu M, Allakhverdiev SI, Shabala S. Effects of Iron Oxide Nanoparticles (Fe3O4) on Growth, Photosynthesis, Antioxidant Activity and Distribution of Mineral Elements in Wheat (Triticum aestivum) Plants. Plants. 2022; 11(14):1894. https://doi.org/10.3390/plants11141894
Chicago/Turabian StyleFeng, Yingming, Vladimir D. Kreslavski, Alexander N. Shmarev, Anatoli A. Ivanov, Sergey K. Zharmukhamedov, Anatoliy Kosobryukhov, Min Yu, Suleyman I. Allakhverdiev, and Sergey Shabala. 2022. "Effects of Iron Oxide Nanoparticles (Fe3O4) on Growth, Photosynthesis, Antioxidant Activity and Distribution of Mineral Elements in Wheat (Triticum aestivum) Plants" Plants 11, no. 14: 1894. https://doi.org/10.3390/plants11141894