
Genome Size Variation Assessment in Vitis vinifera L. Landraces in Ibiza and Formentera (Balearic Islands)


Abstract
:1. Introduction
2. Results and Discussion
2.1. Comparison with Previous Results on Vitis Genome Size
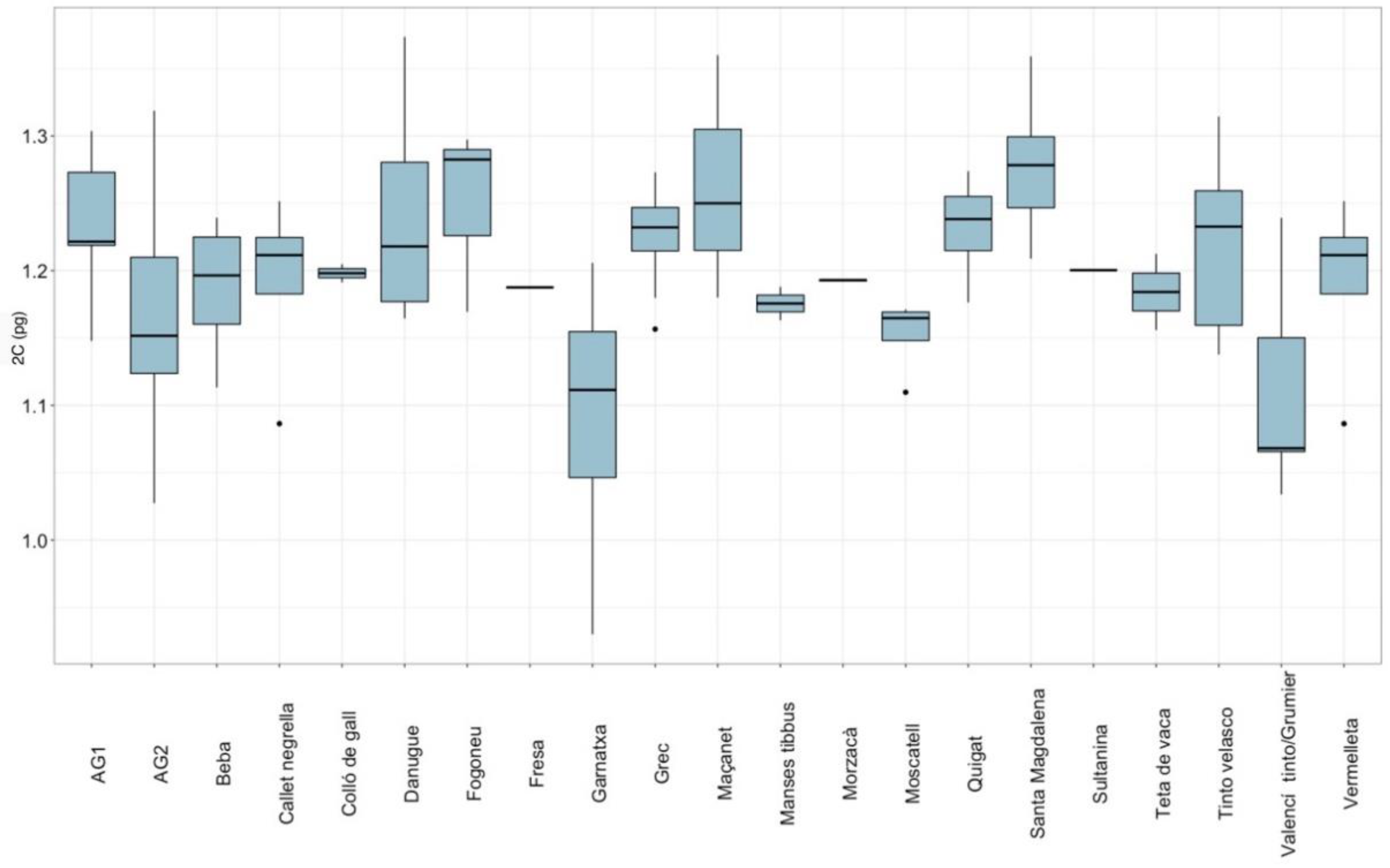
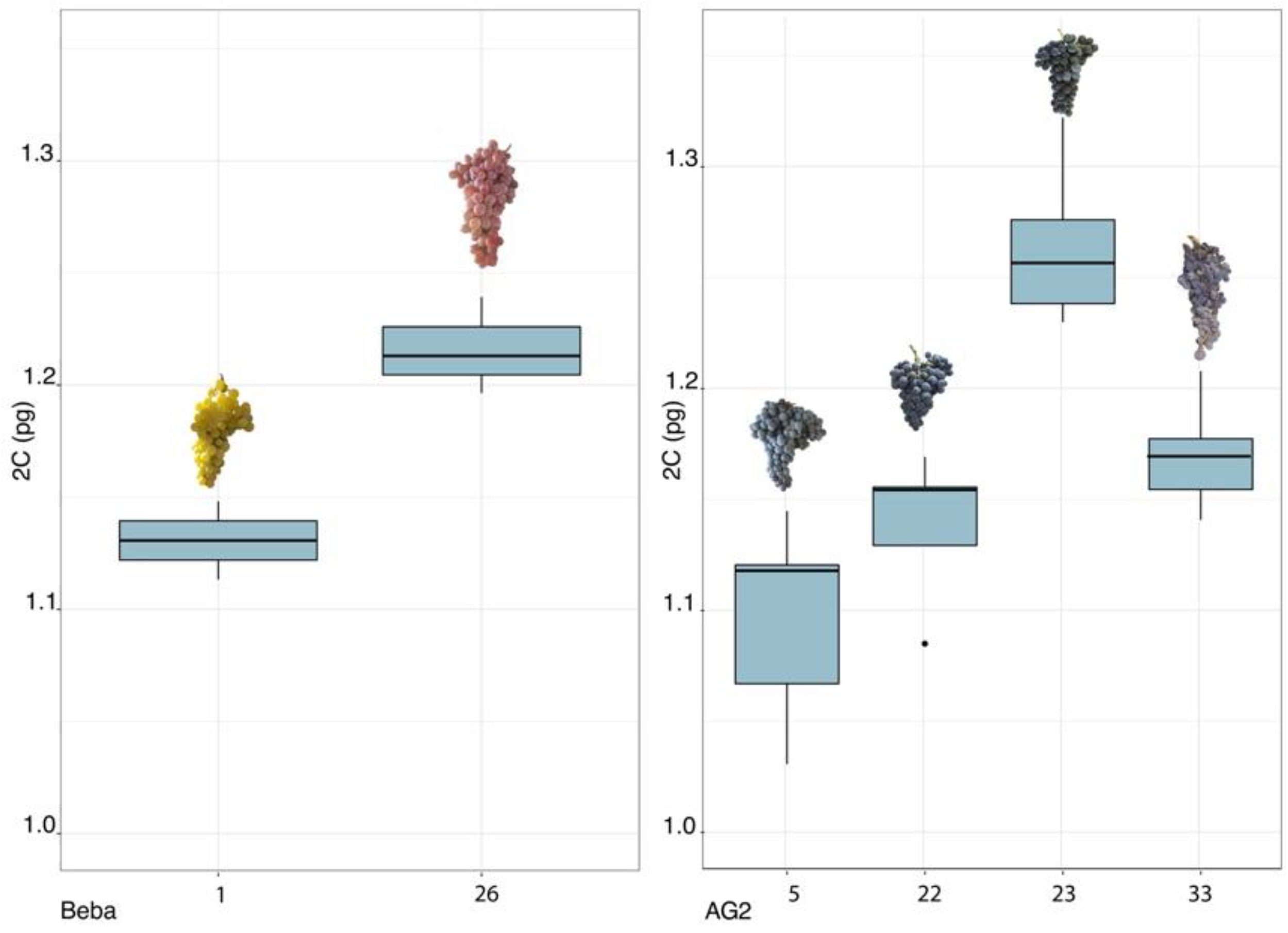
2.2. Genome Size Variation in Pityusic Vitis
3. Materials and Methods
3.1. Plant Material and Origin
3.2. Genome Size Assessment
3.3. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- International Organisation of Vine and Wine. Actualidad de la Coyuntura del Sector Vitivinícola Muncial. Available online: https://www.oiv.int/js/lib/pdfjs/web/viewer.html?file=/public/medias/8780/es-state-of-the-world-vine-and-wine-sector-abril-2022.pdf (accessed on 19 May 2022).
- Escudero, D.; Mancheño, S.; López, J.J. Capítulo 7: Superficie y producciones de cultivos. In Anuario de Estadística. Avance 2021; Ministerio de Agricultura, Pesca, y Alimentación: Madrid, Spain, 2022; pp. 878–882. Available online: https://www.mapa.gob.es/estadistica/pags/anuario/2021-Avance/AVANCE_ANUARIO/AvAE21.pdf (accessed on 5 July 2022).
- Illes Balears Qualitat. Vi de la Terra d’Eivissa. Available online: http://www.illesbalearsqualitat.es/iquafront/producte/353 (accessed on 13 April 2022).
- Illes Balears Qualitat. Vi de la Terra de Formentera. Available online: http://www.illesbalearsqualitat.es/iquafront/producte/627 (accessed on 13 April 2022).
- Ruiz-García, L.; Romero, P.; Tornel, M.; Menéndez, C.; Cabello, F.; Martínez-Cutillas, A. La viticultura frente al cambio climático: Adaptación y estrategias de mejora. In Influencia del Cambio Climático en la Mejora Genética de Plantas, 1st ed.; García, J., Pérez, O., Cos, J., Ruiz-García, L., Sánchez, E., Eds.; Comunidad Autónoma de la Región de Murcia, Consejería de Agua, Agricultura, Ganadería y Pesca, Dirección General de Agricultura, Ganadería, Pesca y Acuicultura, Servicio de Formación y Transferencia Tecnológica: Murcia, Spain, 2018; pp. 165–198. [Google Scholar]
- Bota, J. Estado actual de las variedades minoritarias de las Islas Baleares. In Proceedings of the Reunión del Grupo Español de Seleccionadores de Vid (GESEVID), Madrid, Spain, 12–13 November 2012. [Google Scholar]
- Registre Vitivinícola de les Illes Balears. Available online: http://registreviticola.caib.es/viticolafront/ (accessed on 2 February 2021).
- Royal Decree 1338/2018 of 29 October, Which Regulates Wine Production Potential. BOE nº 262. 30 October 2018. pp. 104825–104885. Available online: https://www.boe.es/buscar/doc.php?id=BOE-A-2018-14803 (accessed on 16 April 2022).
- Oliver, M. Vitivinicultura balear extremos de su historia. In I Encuentro de Historiadores de la Vitivinicultura. In Proceedings of I Encuentro de Historiadores de la Vitivinicultura, El Puerto de Santa María, Spain, 2000; Maldonado, J., Ramos, A., Eds.; Ayuntamiento de El Puerto de Santa María: El Puerto de Santa María, Spain, 2000; pp. 535–552. [Google Scholar]
- Carámbula, C.; Tomás, M.; Medrano, H.; Cifre, J. Selección clonal, saneamiento y conservación de material vegetal vitícola de Baleares. Tecnol. Del Vino Trat. Y Equipos Para Vitic. Y Enol. 2005, 22, 79–84. [Google Scholar]
- Carámbula, C.; Moreno, M.T.; Riera, D.; Cretazzo, E.; Tomás, M.; Pou, A.; Escalona, J.M.; Martorell, A.; Medrano, H.; Cifre, J. Selección clonal de las principales variedades autóctonas de Baleares. In Proceedings of the XXIX Congreso Mundial de la Viña y del Vino; Logroño, Spain, 25–30 June 2006, Ministerio de Agricultura, Pesca y Alimentación: Madrid, Spain, 2006. [Google Scholar]
- Jiménez, J.L. Recuperación de Variedades Locales de Viña Por Saneamiento Mediante Cultivo in Vitro. Bachelor’s Thesis, Enginyeria Agroalimentària i del Medi Rural (GEAM), Universitat de les Illes Balears, Palma de Mallorca, Spain, 2017. [Google Scholar]
- Escalona, J.; March, J.; Luna, J.; Martorell, A. Recuperación de variedades minoritarias de Baleares. In Proceedings of the 5° Foro Mundial del Vino, Logroño, Spain, 4 November 2005. [Google Scholar]
- García, S. Estudio de Variedades Minoritarias de vid (Vitis vinifera L.): Descripción, Caracterización Agronómica y Enológica de Material Procedente de las Islas Baleares. Ph.D. Thesis, Universidad de Valladolid, Valladolid, Spain, 2011. [Google Scholar]
- Medrano, H.; Balda Manzanos, P.J. Informe de Justificación Técnica. Projecte AIA03/15. Les Varietats Minoritàries de Vinya de les Illes Balears: Recuperació i Sanejament de Varietats, Identificació Morfològica i Molecular, Autorització i Certificació de Material Vegetal de Vinya; Govern de les Illes Balears, Conselleria d’Agricultura i Pesca, Fogaiba, Universitat de les Illes Balears, Fogaiba: Palma de Mallorca, Spain, 2018; Available online: http://inagea.com/wp-content/uploads/2018/03/Informe-FINAL.pdf (accessed on 5 May 2021).
- Swift, H. The Constancy of Desoxyribose Nucleic Acid in Plant Nuclei. Proc. Natl. Acad. Sci. USA 1950, 36, 643–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leitch, I. Plant Genome Diversity Volume 2; Greilhuber, J., Dolezel, J., Wendel, J.F., Eds.; Springer: Vienna, Austria, 2013; Volume 2, ISBN 978-3-7091-1159-8. [Google Scholar]
- Pellicer, J.; Hidalgo, O.; Dodsworth, S.; Leitch, I. Genome Size Diversity and Its Impact on the Evolution of Land Plants. Genes 2018, 9, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennet, M.; Leitch, I. Plant Genome Size Research: A Field in Focus. Ann. Bot. 2005, 95, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennett, M.; Leitch, I. Genome size evolution in plants. In The Evolution of the Genome; Gregory, T., Ed.; Elsevier Inc.: San Diego, CA, USA, 2005; pp. 89–162. [Google Scholar]
- Prado, M.J.; Rodriguez, E.; Rey, L.; González, M.V.; Santos, C.; Rey, M. Detection of somaclonal variants in somatic embryogenesis-regenerated plants of Vitis vinifera by flow cytometry and microsatellite markers. Plant. Cell Tiss Organ. Cult. 2010, 103, 49–59. [Google Scholar] [CrossRef]
- Bracho, J. Evaluación del Tamaño del Genoma y Estudio de la Ploidía en una Colección de Berenjenas Cultivadas y Especies Silvestres Relacionadas. Master’s Thesis, Universidad Politécnica de Valencia, Valencia, Spain, 2019. [Google Scholar]
- Arumuganathan, K.; Earle, E.D. Nuclear DNA Content of Some Important Plant Species. Plant. Mol. Biol. Rep. 1991, 93, 208–218. [Google Scholar] [CrossRef]
- Lodhi, M.A.; Reisch, B.I. Nuclear DNA content of Vitis species, cultivars, and other genera of the Vitaceae. Theoret. Appl. Genet. 1995, 90, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Leal, F.; Loureiro, J.; Rodriguez, E.; Pais, M.S.; Santos, C.; Pinto-Carnide, O. Nuclear DNA content of Vitis vinifera cultivars and ploidy level analyses of somatic embryo-derived plants obtained from another culture. Plant. Cell Rep. 2006, 25, 978–985. [Google Scholar] [CrossRef] [PubMed]
- Loureiro, J.O.; Rodriguez, E.; Doležel, J.; Santos, C. Two New Nuclear Isolation Buffers for Plant DNA Flow Cytometry: A Test with 37 Species. Ann. Bot. 2007, 100, 875–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, Z.F.; Wen, J.; Yang, Y.P.; Nie, Z.L.; Meng, Y. Genome size variation and evolution in the grape family Vitaceae. J. Syst. Evol. 2018, 56, 273–282. [Google Scholar] [CrossRef] [Green Version]
- Doležel, J.; Bartos, J.; Voglmayr, H.; Greilhuber, J. Nuclear DNA Content and Genome Size of Trout and Human. Cytometry 2003, 51A, 127–128. [Google Scholar] [CrossRef] [PubMed]
- Garnatje, T.; Garcia, S.; Hidalgo, O.; Pellicer, J.; Sánchez-Jiménez, I.; Vallès, J. Cheirolophus intybaceus (Asteraceae, Centaureinae) o la constància del valor 2C. Collect. Bot. 2009, 28, 7–17. [Google Scholar] [CrossRef] [Green Version]
- Pellicer, J.; Garcia, S.; Garnatje, T.; Vallés, J. Changes in genome size in a fragmented distribution area: The case of Artemisia Crithmifolia L. (Asteraceae, Anthemideae). Caryologia 2009, 62, 152–160. [Google Scholar]
- Marí, V. (Farmer). Personal Communication, 2020.
- González, R.; Vallès, J.; Garnatje, T.; De Andrés, M.T.; Vargas, A.M. Laboratori de Botànica-Unitat associada al CSIC, Facultat de Farmàcia i Ciències de l’Alimentació-Institut de la Biodiversitat IRBio, Universitat de Barcelona, Barcelona, Spain. 2022; manuscript in preparation. [Google Scholar]
- Bennett, M. Allium cepa, L. Cultivars from Four Continents Compared by Flow Cytometry show Nuclear DNA Constancy. Ann. Bot. 2000, 85, 351–357. [Google Scholar] [CrossRef] [Green Version]
- Hidalgo, O.; Vallès, J.; Romo, A.; Canela, M.Á.; Garnatje, T. Genome size variation in gymnosperms under different growth conditions. Caryologia 2015, 68, 92–96. [Google Scholar] [CrossRef] [Green Version]
- Stelzer, C.P.; Pichler, M.; Hatheuer, A. Linking genome size variation to population phenotypic variation within the rotifer, Brachionus asplanchnoidis. Commun. Biol. 2021, 4, 596. [Google Scholar] [CrossRef] [PubMed]
- Wright, S.I. Evolution of Genome Size. In eLS; John Wiley & Sons: Chichester, UK, 2017; pp. 1–6. [Google Scholar]
- Greilhuber, J. Intraspecific variation in genome size in angiosperms: Identifying its existence. Ann. Bot. 2005, 95, 91–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doležel, J.; Binarová, P.; Lucretti, S. Analysis of Nuclear DNA content in plant cells by Flow Cytometry. Biol. Plant. 1989, 31, 113–120. [Google Scholar] [CrossRef]
- Marie, D.; Brown, S.C. A Cytometric exercise in plant DNA histograms, with 2C values for 70 species. Biol. Cell 1993, 78, 41–51. [Google Scholar] [CrossRef]
- Garnatje, T.; Garcia, S.; Vilatersana, R.; Vallès, J. Genome Size Variation in the genus Carthamus (Asteraceae, Cardueae): Systematic Implications and Additive Changes During Allopolyploidization. Ann. Bot. 2006, 97, 461–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Microsoft Corporation Microsoft Excel. Version 16.60. Available online: https://office.microsoft.com/excel (accessed on 22 February 2022).
- IBM SPSS Statistics for Macintosh, Version 26.0; IBM Corp.: Armonk, NY, USA, 2016.
- Wickham, H.H. ggplot2 Elegant Graphics for Data Analysis (Use R!); Springer: Cham, Switzerland, 2016. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]



{kind=link}
{kind=link}
| Group | N 1 | 2C ± SD (pg) 2 | 1C (Mbp) 3 | HPCV ± SD Plant | HPCV ± SD Standard |
|---|---|---|---|---|---|
| Santa Magdalena | 7 | 1.277 ± 0.051 | 624.453 | 3.834 ± 1.441 | 1.909 ± 0.857 |
| Maçanet | 3 | 1.263 ± 0.091 | 617.607 | 3.242 ± 1.961 | 1.912 ± 1.400 |
| Fogoneu | 3 | 1.250 ± 0.070 | 611.250 | 3.180 ± 0.907 | 0.899 ± 0.431 |
| Danugue | 6 | 1.240 ± 0.081 | 606.360 | 4.008 ± 1.656 | 1.856 ± 0.901 |
| Quigat | 4 | 1.232 ± 0.041 | 602.448 | 3.794 ± 1.493 | 2.369 ± 0.902 |
| AG1 | 9 | 1.229 ± 0.050 | 600.981 | 4.073 ± 1.091 | 2.129 ± 0.837 |
| Grec | 9 | 1.227 ± 0.039 | 603.426 | 4.473 ± 0.094 | 1.864 ± 0.880 |
| Tinto velasco | 5 | 1.221 ± 0.073 | 597.069 | 4.594 ± 1.881 | 1.569 ± 0.631 |
| Sultanina | 1 | 1.200 * | 586.800 | 4.875 ± 0.233 | 0.955 ± 0.714 |
| Colló de gall | 2 | 1.198 ± 0.010 | 585.822 | 4.280 ± 0.421 | 1.103 ± 0.734 |
| Morzacà | 2 | 1.193 ± 0.001 | 583.377 | 3.990 ± 0.391 | 2.160 ± 0.944 |
| Callet negrella | 5 | 1.191 ± 0.064 | 582.399 | 4.592 ± 1.501 | 2.210 ± 1.374 |
| Vermelleta | 5 | 1.191 ± 0.064 | 582.399 | 4.366 ± 1.327 | 1.840 ± 1.174 |
| Beba | 7 | 1.188 ± 0.047 | 580.932 | 4.421 ± 0.939 | 1.961 ± 1.019 |
| Fresa | 1 | 1.188 * | 580.932 | 4.845 ± 0.318 | 3.545 ± 0.403 |
| Teta de vaca | 4 | 1.184 ± 0.024 | 578.976 | 4.571 ± 1.346 | 3.068 ± 0.557 |
| Manses tibbus | 2 | 1.176 ± 0.018 | 575.064 | 3.043 ± 2.360 | 1.105 ± 0.527 |
| AG2 | 20 | 1.164 ± 0.072 | 569.196 | 4.186 ± 1.669 | 1.583 ± 0.900 |
| Moscatell | 4 | 1.153 ± 0.029 | 563.817 | 3.705 ± 1.989 | 1.293 ± 0.863 |
| Valencí tinto/Grumier | 5 | 1.111 ± 0.083 | 543.279 | 4.347 ± 1.320 | 1.399 ± 0.795 |
| Garnatxa | 4 | 1.090 ± 0.117 | 533.010 | 4.888 ± 1.658 | 1.783 ± 0.659 |
| Landrace | Accession Code | Accession Code | p < 0.05 |
|---|---|---|---|
| AG2 | 5 | 23 | 0 |
| 5 | 33 | 0.024 | |
| 22 | 23 | 0 | |
| 23 | 33 | 0.004 | |
| Beba | 1 | 26 | 0.026 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
González, R.; Vallès, J.; Garnatje, T. Genome Size Variation Assessment in Vitis vinifera L. Landraces in Ibiza and Formentera (Balearic Islands). Plants 2022, 11, 1892. https://doi.org/10.3390/plants11141892
González R, Vallès J, Garnatje T. Genome Size Variation Assessment in Vitis vinifera L. Landraces in Ibiza and Formentera (Balearic Islands). Plants. 2022; 11(14):1892. https://doi.org/10.3390/plants11141892
Chicago/Turabian StyleGonzález, Raquel, Joan Vallès, and Teresa Garnatje. 2022. "Genome Size Variation Assessment in Vitis vinifera L. Landraces in Ibiza and Formentera (Balearic Islands)" Plants 11, no. 14: 1892. https://doi.org/10.3390/plants11141892