
Physiological Studies and Ultrastructure of Vigna sinensis L. and Helianthus annuus L. under Varying Levels of Nitrogen Supply

 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Changes in Growth Parameters
2.2. Changes in Nitrogen Fractions
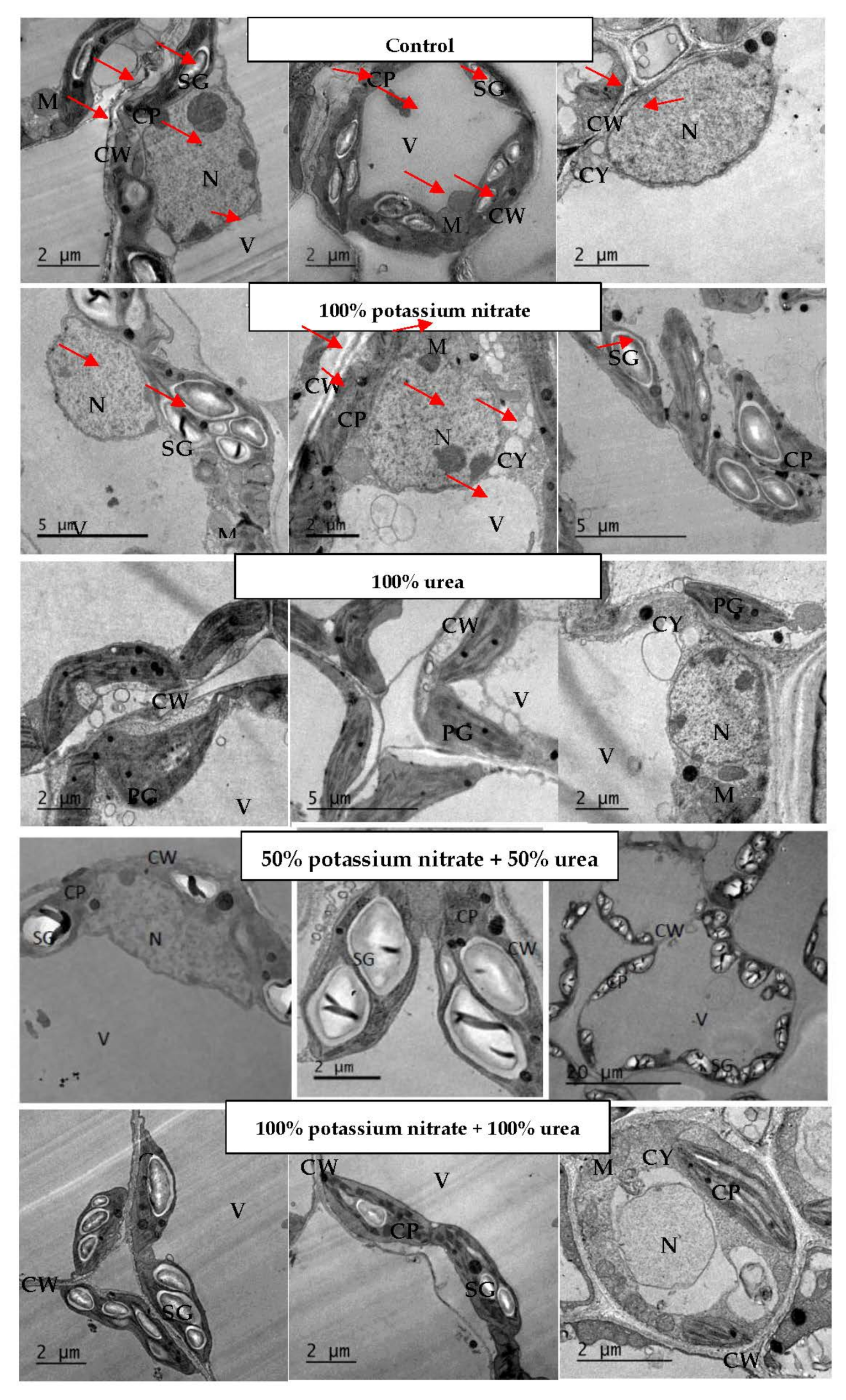
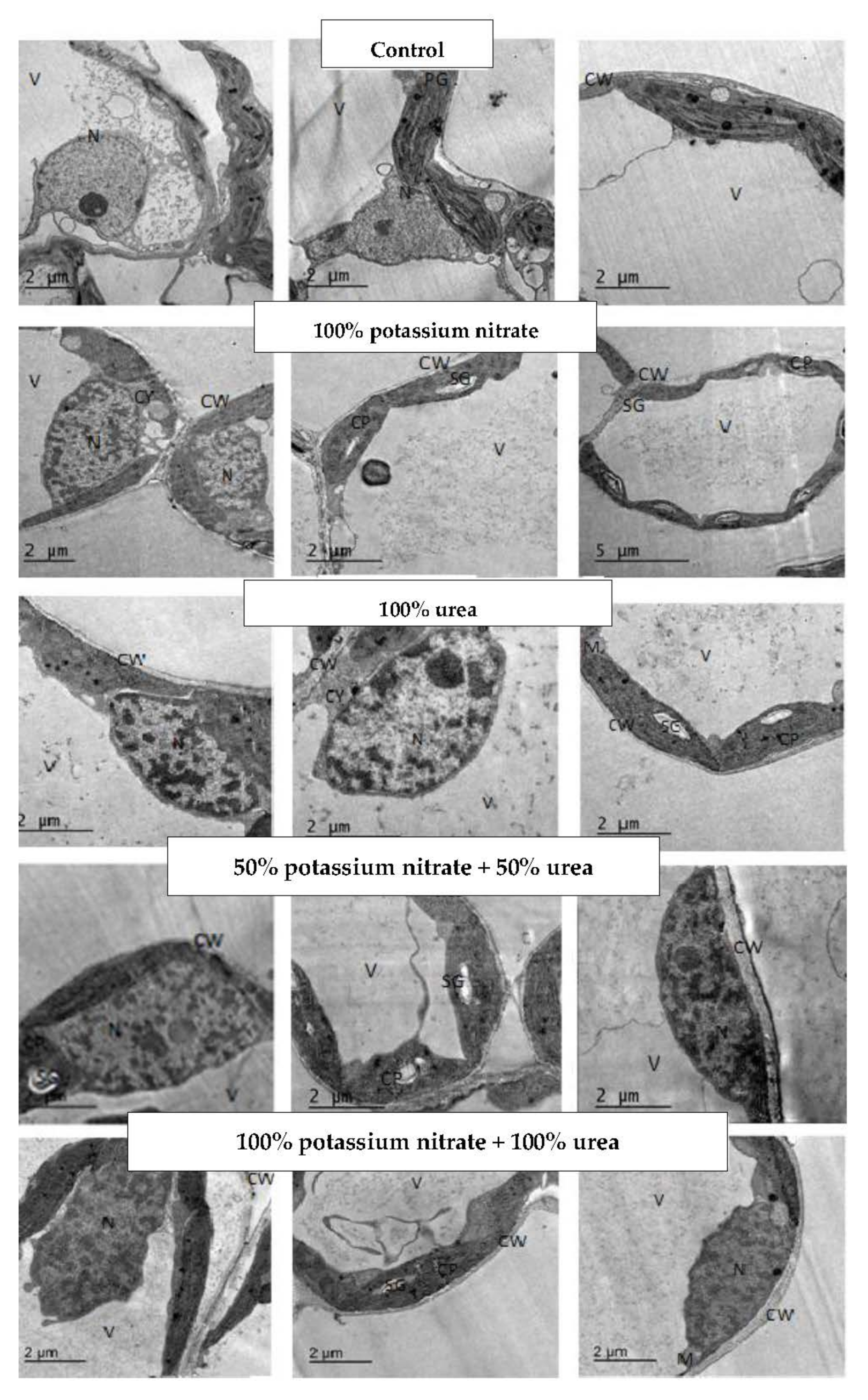
2.3. Changes in Leaf Ultrastructure
3. Discussion
3.1. Changes in Growth Parameters
3.2. Changes in Nitrogen Content
3.3. Changes in Leaf Ultrastructure
4. Materials and Methods
4.1. Plants Used
4.2. Time Course of Experiment
4.3. Analytical Studies
4.4. Transmission Electron Microscopy (TEM)
4.4.1. Fixation
4.4.2. Dehydration
4.4.3. Infiltration
4.4.4. Sectioning
4.4.5. Grid Preparation
4.4.6. Staining
4.4.7. Reynolds Lead Citrate Stains
4.4.8. Examination with TEM
4.4.9. Cell Structure Analysis
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| N | nitrogen |
| K | potassium |
| N/ha | nitrogen/hectare |
| kg N/fed | kilogram nitrogen/feddan |
| kg/h | kilogram/hectare |
| hrs | hours |
| min | minutes |
| mm | millimeter |
| g | gram |
References
- Summerfield, R.J.; Huxley, P.A.; Steelle, W. Cowpea (Vigna unguiculata L. Walp). Field Crops Abstr. 1974, 27, 301–312. [Google Scholar]
- Mohamed, A.A.; Mazrou, Y.; Elhag, D.; Saad, A.; Hamdy, R.; Abdelaal, K.A. Response of some faba bean varieties to amino acids and potassium foliar application under delta soil conditions. Fresenius Environ. Bull. 2021, 30, 11816–11830. [Google Scholar]
- El Nahhas, N.; AlKahtani, M.; Abdelaal, K.A.; Al Husnain, L.; AlGwaiz, H.; Hafez, Y.M.; Attia, K.; El-Esawi, M.; Ibrahim, M.; Elkelish, A. Biochar and jasmonic acid application attenuates antioxidative systems and improves growth, physiology, nutrient uptake and productivity of faba bean (Vicia faba L.) irrigated with saline water. Plant Physiol. Biochem. 2021, 166, 807–817. [Google Scholar] [CrossRef]
- Khaffagy, A.E.; Mazrou, Y.S.A.; Morsy, A.R.; El-Mansoury, M.A.M.; El-Tokhy, A.I.; Hafez, Y.; Abdelaal, K.A.; Khedr, R.A. Impact of Irrigation Levels and Weed Control Treatments on Annual Weeds, Physiological Traits and Productivity of Soybean under Clay Soil Conditions. Agronomy 2022, 12, 1037. [Google Scholar] [CrossRef]
- Abdelaal, K.; El-Okkiah, S.; Metwaly, M.; El-Afry, L. Impact of Ascorbic acid and proline application on the physiological machinery in soybean plants under salinity stress. Fresenius Environ. Bull. 2021, 30, 12486–12497. [Google Scholar]
- El-Afry, M.; Ahmed, H.; Mohsen, R.; Abd El-Kader, N.; Ismail, I.A.; Abdelaal, K.A. Influence of some foliar treatments on growth, yield and quality of pea (Pisum sativum L.). Fresenius Environ. Bull. 2021, 30, 5765–5772. [Google Scholar]
- Abdelaal, K.; Mazrou, Y.; Hafez, Y. Effect of silicon and carrot extract on morphophysiological characters of pea (Pisum sativum L.) under salinity stress conditions. Fresenius Environ. Bull. 2022, 31, 608–615. [Google Scholar]
- Arafa, S.A.; Attia, K.; Niedbała, G.; Piekutowska, M.; Alamery, S.; Abdelaal, K.; Alateeq, T.; Ali, M.A.M.; Elkelish, A.; Attallah, S.Y. Seed Priming Boost Adaptation in Pea Plants under Drought Stress. Plants 2021, 10, 2201. [Google Scholar] [CrossRef]
- Ezin, V.; Tosse, A.G.C.; Chabi, I.B.; Ahanchede, A. Adaptation of Cowpea (Vigna unguiculata (L.) Walp.) to Water Deficit during Vegetative and Reproductive Phases Using Physiological and Agronomic Characters. Int. J. Agron. 2021, 2021, 9665312. [Google Scholar] [CrossRef]
- Fery, R.L. New Opportunities in Vigna. In Trends in New Crops and New Ideas; Janick, J., Whipkey, A., Eds.; ASHS Press: Alexandria, VA, USA, 2000; pp. 424–428. [Google Scholar]
- Ndungu, S.M.; Messmer, M.M.; Ziegler, D.; Gamper, H.A.; Mészáros, E.; Thuita, M.; Vanlauwe, B.; Frossard, E.; Thonar, C. Cowpea (Vigna unguiculata L. Walp) hosts several widespread bradyrhizobial root nodule symbionts across contrasting agro-ecological production areas in Kenya. J. Agric. Ecosyst. Environ. 2018, 261, 161–171. [Google Scholar] [CrossRef]
- Naseem, Z.; Masood, S.A.; Ali, Q.; Ali, A.; Kanwal, N. Study of genetic variability in Helianthus annuus for seedling traits: An Overview. Life Sci. J. 2015, 12, 109–114. [Google Scholar]
- Cevallos-Casals, B.A.; Cisneros-Zevallos, L. Impact of germination on phenolic content and antioxidant activity of 13 edible seed species. Food Chem. 2010, 119, 1485–1490. [Google Scholar] [CrossRef]
- Adeleke, B.S.; Babalola, O.O. Oilseed crop sunflower (Helianthus annuus) as a source of food: Nutritional and health benefits. Food Sci. Nutr. 2020, 8, 4666–4684. [Google Scholar] [CrossRef] [PubMed]
- Khurana, S.; Singh, R. Sunflower (Helianthus annuus) Seed. In Oilseeds: Health Attributes and Food Applications; Tanwar, B., Goyal, A., Eds.; Springer: Singapore, 2021. [Google Scholar] [CrossRef]
- Ewert, F.; Honermeier, B. Spikelet initiation of winter triticale and winter wheat in response to nitrogen fertilization. Eur. J. Agron. 1999, 11, 107–113. [Google Scholar] [CrossRef]
- Kumawat, S.M.; Dhakar, L.L.; Maliwal, P.L. Effect of irrigation regimes and nitrogen on yield, oil content and nutrient uptake of soybean (Glycine max). Indian J. Agron. 2000, 45, 361–366. [Google Scholar]
- Abdelaal, K.A. Pivotal Role of Bio and Mineral Fertilizer Combinations on Morphological, Anatomical and Yield Characters of Sugar Beet Plant (Beta vulgaris L.). Middle East J. Agric. Res. 2015, 4, 717–734. [Google Scholar]
- Abou Khadrah, S.; Gharib, H.S.; Mohamed, A.A.; Elhosary, M.A.; Abdelaal, K.A.; Hafez, Y.M. Combination of nitrogen and potassium fertilizers improve physiological and yield characters of two wheat cultivars. Fresenius Environ. Bull. 2020, 29, 8998–9004. [Google Scholar]
- Hafez, E.M.; Abdelaal, K.A. Impact of Nitrogen fertilization levels on morphophysiological characters and yield quality of some Maize hybrids (Zea mays L.). Egyption J. Agron. 2015, 37, 35–48. [Google Scholar]
- Rosenblueth, M.; Ormeño-Orrillo, E.; López-López, A.; Rogel, M.A.; Reyes-Hernandez, B.J.; Martínez-Romero, J.C.; Reddy, P.M.; Martinez-Romero, E. Nitrogen fixation in cereals. Front. Microbiol. 2018, 9, 1794. [Google Scholar] [CrossRef] [Green Version]
- Hord, N.G.; Tang, Y.; Bryan, N.S. “Food sources of nitrates and nitrites: The physiologic context for potential health benefits” (PDF). Am. J. Clin. Nutr. 2009, 90, 1–10. [Google Scholar] [CrossRef]
- Laue, W.; Thiemann, M.; Scheibler, E.; Wiegand, K.W. Nitrates and Nitrites. In Ullmann’s Encyclopedia of Industrial Chemistry; Wiley-VCH: Weinheim, Germany, 2006. [Google Scholar]
- Salardini, A.A. Soil Fertility; Tehran University Press: Persian, Iran, 1987; p. 44. (In Persian) [Google Scholar]
- Klimczyk, M.; Siczek, A.; Schimmelpfennigb, L. Improving the efficiency of urea-based fertilization leading to reduction in ammonia emission. Sci. Total Environ. 2021, 771, 145483. [Google Scholar] [CrossRef] [PubMed]
- Caliskan, S.; Ozkaya, I.; Caliskan, M.E.; Arslan, M. The effects of nitrogen and iron fertilization on growth, yield and fertilizer use efficiency of soybean in a Mediterranean-type soil. Field Crops Res. 2008, 108, 126–132. [Google Scholar] [CrossRef]
- Harper, J.E.; Hageman, R.H. Canopy and seasonal profiles of nitrate reductase in soybean (Glycine max (L.) Merrill). Plant Physiol. 1972, 62, 662–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osborne, S.L.; Riedell, W.E. Starter nitrogen fertilizer impact on soybean yield and quality in the Northern Great Plains. Agron. J. 2006, 98, 1569–1574. [Google Scholar] [CrossRef] [Green Version]
- Beard, B.H.; Hoover, R.M. Effect of nitrogen on nodulation and yield of irrigated soybeans. Agron. J. 1971, 63, 815–816. [Google Scholar] [CrossRef]
- Koutroubas, S.D.; Papakosta, D.K.; Gagianas, A.A. The importance of early dry matter and nitrogen accumulation in soybean yield. Eur. J. Agron. 1998, 9, 1–10. [Google Scholar] [CrossRef]
- Pervez, M.A.; Ayub, C.M.; Bashart, A.; Nave, A.V.; Nasir, M. Effect of nitrogen levels and spacing on growth and yield of radish (Raphanus sativus L.). Int. J. Agric. Biol. 2004, 6, 504–506. [Google Scholar]
- Aminifard, M.H.; Aroiee, H.; Nemati, H.; Azizi, M.; Khayyat, M. Effect of nitrogen fertilizer on vegetative and reproductive growth of pepper plants under field conditions. J. Plant Nutr. 2012, 35, 235–242. [Google Scholar] [CrossRef]
- Bowen, P.; Frey, B. Response of plasticultare bell pepper to staking, irrigation frequency and fertigated nitrogen rate. J. Hortic. Sci. 2002, 37, 95–100. [Google Scholar]
- Aroiee, H.; Omidbaigi, R. Effects of nitrogen fertilizer on productivity of medicinal pumpkin. J. Acta Hortic. 2004, 629, 415–419. [Google Scholar] [CrossRef]
- Boroujerdnia, M.; Alemzadeh, A.N. Effect of different levels of nitrogen fertilizer and cultivars on growth, yield and yield components of romaine lettuce (Lactuca sativa L.). Middle East. Russ. J. Plant Sci. Biotechnol. 2007, 1, 47–53. [Google Scholar]
- Tei, F.; Benincasa, P.; Guiducci, M. Effect of nitrogen availability on growth and nitrogen uptake in lettuce. Acta Hortic. 2000, 533, 385–392. [Google Scholar] [CrossRef]
- Takebe, M.; Ishihara, T.; Matsuna, K.; Fojimoto, J.; Yoneyama, T. Effect of nitrogen application on the content sugars, ascorbic acid, nitrate and oxalic acid in spinach (Spinacia oleracea L.) and kamatsuna (Nrasica compestris L.). Jpn. J. Soil Sci. Plant Nutr. 1995, 66, 238–246. [Google Scholar]
- Stales, H.; Inze, D. When plant cells decide to divide. Trends Plant Sci. 2001, 8, 359–364. [Google Scholar] [CrossRef]
- Almodares, A.; Taheri, R.; Chug, I.M.; Fathi, M. The Effect of nitrogen and potassium fertilizers on growth parameters and carbohydrate contents of sweet sorghum cultivars. J. Environ. Biol. 2008, 29, 849–852. [Google Scholar]
- Kodithuwakku, D.P.; Kirthisinghe, J.P. The Effect of different rates of nitrogen fertilizer application on the growth, yield and postharvest life of cauliflower. Trop. Agric. Res. 2009, 21, 110–114. [Google Scholar] [CrossRef] [Green Version]
- Shirazi, S.M.; Sholichin, M.; Jameel, M.; Akib, S.; Azizi, M. Effects of different irrigation regimes and nitrogenous fertilizer on yield and growth parameters of maize. Int. J. Phys. Sci. 2011, 6, 677–683. [Google Scholar]
- Tolba, M.S. Influence of Different Nitrogenous and Potassic Fertilization Levels on Vegetative Growth, Heads Yield and Chemical Composition of Broccoli (Brassica oleracea var. italica). Ph.D. Thesis, Faculty of Agriculture, Fayoum, Cairo University, Cairo, Egypt, 2005. [Google Scholar]
- El-Masry, T.A.; Ashraf, S.H.; Mofreh, O.S.; Abd El-Mohsen, Y.H. Increasing nitrogen efficiency by humic acid soil application to squash plants (Cucurbita pepo L.) grown in newly reclaimed saline soil. Egypt J. Hort. 2014, 41, 17–38. [Google Scholar]
- Haroun, S.A. Influence of site of oxyfluorfen application on growth, pigments, photosynthesis and yield attributes of Glycine max plants. Pak. J. Biol. Sci. 2002, 5, 292–296. [Google Scholar] [CrossRef] [Green Version]
- Morshed, R.M.; Rahman, M.M.; Rahman, M.A. Effect of nitrogen on seed yield, protein content and nutrient uptake of soybean (Glycine max L.). J. Agric. Rural. Dev. 2008, 6, 13–17. [Google Scholar] [CrossRef] [Green Version]
- Eman, S.S. Response of growth, yield and attributes of soybean plants [(Glycin max (L.) Merr.)] to late soil nitrogen fertilization. Arab. Univ. J. Agric. Sci. 2002, 10, 165–172. [Google Scholar]
- Chandel, G.; Banerjee, S.; See, S.; Meena, R.; Sharma, D.J.; Verulkar, S.B. Effect of different nitrogen fertilizer levels and native soil properties on rice grain Fe, Zn and protein contents. Rice Sci. 2010, 17, 213–227. [Google Scholar] [CrossRef]
- Sugiyama, T.; Mizuno, M.; Hayashi, M. Partitioning of nitrogen among ribulose-1, 5-bisphosphate carboxylase/oxygenase, phosphoenolpyruvate carboxylase, and pyruvate orthophosphate dikinase as related to biomass productivity in maize seedlings. Plant Physiol. 1984, 75, 665–669. [Google Scholar] [CrossRef] [Green Version]
- Greef, J.M. Productivity of maize (Zea mays L.) in relation to morphological and physiological characteristics under varying amounts of nitrogen supply. J. Agron. Crop Sci. 1994, 172, 317–326. [Google Scholar] [CrossRef]
- Taize, L.; Zeiger, E. Plant Physiology; Sinauer Associates: Sunderland, MA, USA, 2000; pp. 259–266. [Google Scholar]
- Faraji, F.; Esfahani, M.; Kavousi, M.; Nahvi, M.; Forghani, A. Effect of nitrogen fertilizer levels on Fe and protein content, grain breakage and grain yield of rice (Oryza sativa L. cv. Khazar). Biharean Biol. 2013, 7, 25–28. [Google Scholar]
- Fageria, N.K. Plant tissue test for determination of optimum concentration and uptake of nitrogen at different growth stages in lowland rice. Commun. Soil Sci. Plant Anal. 2003, 34, 259–270. [Google Scholar] [CrossRef]
- Shinano, T.; Osaki, M.; Tadano, T. Comparison of growth efficiency between rice and soybean at the vegetative growth stage. Soil Sci. Plant Nutr. 1995, 41, 471–480. [Google Scholar] [CrossRef]
- Al Hasnawi, R.A.; AlJanaby, Z.A.A.; Jaafer, A.A.; Mohammed, R.J. Effect of nitrogen fertilization and irrigation water quality on some soil characteristics, growth and yield of sunflower. Plant Arch. 2020, 20, 2703–2705. [Google Scholar]
- Kano, Y.; Nakagawa, H.; Sekine, M.; Goto, H.; Sugiura, A. Effect of nitrogen fertilizer on cell size and sugar accumulation in the leaves of cabbage (Brassica oleracea L.). Hortic. Sci. 2007, 42, 1490–1492. [Google Scholar] [CrossRef] [Green Version]
- Kutik, J.; Lubomir, N.; Demmers-Derks, H.H.; Lawlor, D.W. Chloroplast ultrastructure of sugar beet (Beta vulgaris L.) cultivated in normal and elevated CO2 concentrations with two contrasted nitrogen supplies. J. Exp. Bot. 1995, 46, 1797–1802. [Google Scholar] [CrossRef]
- Hak, R.; Rinderle-Zimmer, U.; Lichtenthaler, H.K.; Natr, L. Chlorophyll a fluorescence signatures of nitrogen deficient barley leaves. Photosynthetica 1993, 28, 151–159. [Google Scholar]
- Evans, J.R. Photosynthesis and nitrogen relationships in leaves of C3 plants. Ecology 1989, 78, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Laza, R.C.; Bergman, B.; Vergara, B.S. Cultivar differences in growth and chloroplast ultrastructure in rice as affected by nitrogen. J. Exp. Bot. 1993, 44, 1643–1648. [Google Scholar] [CrossRef]
- Carmi, A.; Shomer, I. Starch accumulation and photosynthetic activity in primary leaves of bean (Phaseolus vulgaris L.). Ann. Bot. 1979, 44, 479–484. [Google Scholar] [CrossRef]
- Bondada, B.R.; Syvertsen, J.P. Leaf chlorophyll, net gas exchange and chloroplast ultrastructure in citrus leaves of different nitrogen tatus. Tree Physiol. 2003, 23, 553–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ariovich, D.; Cresswell, C.F. The effect of nitrogen and phosphorus on starch accumulation and net photosynthesis in two variants of Panicum maximum Jacque. Plant Cell Environ. 1983, 5, 657–664. [Google Scholar] [CrossRef]
- Lee, Y.J.; Yang, C.M.; Chang, K.W.; Shen, Y. Effects of nitrogen status on leaf anatomy, chlorophyll content and canopy reflectance of paddy rice. Bot. Stud. 2011, 52, 295–303. [Google Scholar]
- Omar, A.; AboYoussef, M.; Shoughy, A.; Abd El-Aty, M.S.; Abdelaal, K.A.; Hafez, Y.; Kamara, M. Response of Egyptian Yasmin rice cultivar to different seeding number per hill and different nitrogen levees. Fresenius Environ. Bull. 2022, 31, 1258–1265. [Google Scholar]
- Mosalem, M.; Mazrou, Y.; Badawy, S.; Abd Ullah, M.A.; Mubarak, M.G.; Hafez, Y.M.; Abdelaal, K.A. Evaluation of sowing methods and nitrogen levels for grain yield and components of durum wheat under arid regions of Egypt. Rom. Biotechnol. Lett. 2021, 26, 3031–3039. [Google Scholar] [CrossRef]
- Nasim, W.; Ahmad, A.; Ahmad, S.; Nadeem, M.; Masood, N.; Shahid, M.; Fahad, S. Response of sunflower hybrids to nitrogen application grown under different agro-environments. J. Plant Nutr. 2017, 40, 82–92. [Google Scholar] [CrossRef]
- Neață, G.; Stoian, E.; Mocuța, D.; Temocico, G.; Fîntîneru, G.; Georgescu, M. Nitrates and nitrites as source of nitrous compounds. Rom. Biotechnol. Lett. 2013, 18, 8583–8586. [Google Scholar]
- Ertek, A.; Erdal, I.; Yilmaz, H.I.; Senyigit, U. Water and nitrogen application levels for the optimum tomato yield and water use efficiency. J. Agric. Sci. Technol. 2012, 14, 889–902. [Google Scholar]
- Jose, S.; Gillespie, A.R. Allelopathy in black walnut (Juglans regia L.) alley cropping: II. Effects of juglone on hydroponically grown corn (Zea mays L.) and soybean (Glycine max L. Merr.) growth and physiology. Plant Soil. 1998, 203, 199–205. [Google Scholar] [CrossRef]
- Poorter, H.; Remkes, C. Leaf area ratio and net assimilation rate of 24 wild species differing in relative growth. Oecologia 1990, 53, 553–559. [Google Scholar] [CrossRef] [PubMed]
- Yemm, E.W.; Willis, A.J. The respiration of barely plants. IX. The metabolism of roots during the assimilation of nitrogen. New Phytol. 1956, 55, 229–252. [Google Scholar] [CrossRef]
- Rees, M.W.; Williams, E.F. The total nitrogen content of egg albumin and other proteins. J. Biochem. 1943, 37, 354–359. [Google Scholar]
- Haroun, S.A. Studies on Pigmentation and Metabolism in Leaves. M.Sc. Thesis, Botany Department, Faculty of Science-Mansoura University, Mansoura, Egypt, 1981. [Google Scholar]
- Delory, M. Colourimetric estimation of ammonia. In Inorganic Chemistry; Vogel, H., Ed.; Longman: London, UK, 1949; pp. 126–132. [Google Scholar]
- Naguib, M.I. Effect of seven on carbohydrates and nitrogen metabolism during the germination of cotton seeds. Ind. J. Exp. Biol. 1964, 2, 149–155. [Google Scholar]
- Pirie, N.W. Proteins. In Modern Methods of Plant Analysis; Paech, K., Tracey, M.B., Eds.; Springer: Berlin, Germany, 1955; Volume 4, pp. 23–68. [Google Scholar]
- Haroun, S.A. Studies on Adaptation of Plants to Water Stress. Ph.D. Thesis, Botany Department, Faculty of Science-Mansoura University, Mansoura, Egypt, 1985. [Google Scholar]
- Association of Official Agricultural Chemists. Official methods of analysis. In Index of Method Number 920, 15th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 1990. [Google Scholar]
- Reynolds, E.S. The use of lead citrate at high PH as an electron- opaque stain in electron microscopy. Cell Biol. 1963, 17, 208–212. [Google Scholar] [CrossRef] [Green Version]
- Snedecor, G.W.; Cochran, W.G. Statistical Methods, 7th ed.; 2nd Printing; The Owa State University Press: Iowa City, IA, USA, 1982; 507p. [Google Scholar]



{kind=link}
{kind=link}
| Plant | Treatments | Parameters | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Shoot Length (cm) | Number of Nodes/Plants | Number of Leaves/Plants | Leaves Area/Plant (cm2) | Shoot Fresh Weight (g) | Shoot Dry Weight (g) | Shoot Water Content | Relative Growth Rate | Net Assimilation Rate | ||
| V. sinensis | T1 | 19.375 | 5.375 | 10.75 | 174.878 | 5.121 | 0.753 | 4.368 | 0.062 | 0.00015 |
| T2 | 22.675 * | 6 | 12 | 161.264 | 6.12 | 0.888 | 5.232 | 0.065 | 0.0002 | |
| T3 | 21.75 | 4.875 | 9.75 | 144.43 | 6.494 * | 0.895 | 5.599 * | 0.078 | 0.00023 * | |
| T4 | 22.063 * | 4.875 | 9.75 | 203.872 | 7.788 * | 1.248 * | 6.540 * | 0.084 * | 0.00025 * | |
| T5 | 21.788 | 5.25 | 10.5 | 235.402 * | 6.486 * | 0.895 | 5.591 * | 0.065 | 0.00014 | |
| H. annuus | T1 | 47.563 | 4.875 | 9.75 | 700.471 | 4.009 | 0.556 | 3.453 | 0.055 | 2.438 |
| T2 | 53.625 | 8.375 * | 16.750 * | 1560.98 * | 9.639 * | 1.198 * | 8.441 * | 0.063 | 2.498 | |
| T3 | 55.500 * | 9.500 * | 19.000 * | 1796.246 * | 12.707 * | 1.549 * | 11.158 * | 0.077 * | 3.017 | |
| T4 | 57.620 * | 9.000 * | 18.000 * | 742.475 | 11.769 * | 1.606 * | 10.163 * | 0.078 * | 7.995 * | |
| T5 | 50.188 | 8.750 * | 17.250 * | 1723.659 * | 8.429 * | 1.090 * | 7.339 * | 0.071 * | 2.163 | |
| Plant | Treatments | Parameters | ||
|---|---|---|---|---|
| Total Nitrogen | Insoluble Protein Nitrogen | Total Soluble Nitrogen | ||
| V. sinensis | T1 | 14.538 | 9.813 | 4.725 |
| T2 | 9.026 * | 6.093 * | 2.933 * | |
| T3 | 9.495 * | 6.409 * | 3.086 * | |
| T4 | 14.875 | 10.041 | 4.834 | |
| T5 | 6.932 * | 4.679 * | 2.253 * | |
| H. annuus | T1 | 6.986 | 4.716 | 2.270 |
| T2 | 13.359 * | 9.017 * | 4.342 * | |
| T3 | 12.276 * | 8.286 * | 3.990 * | |
| T4 | 7.094 | 4.789 | 2.305 | |
| T5 | 14.598 * | 9.854 * | 4.744 * | |
| Plant | Treatments | Parameters | ||
|---|---|---|---|---|
| Total Nitrogen | Insoluble Protein Nitrogen | Total Soluble Nitrogen | ||
| V. sinensis | T1 | 0.880 | 0.594 | 0. 286 |
| T2 | 0.950 * | 0.641 * | 0.309 * | |
| T3 | 1.020 * | 0.688 * | 0.332 * | |
| T4 | 1.240 * | 0.837 * | 0.403 * | |
| T5 | 1.050 * | 0.708 * | 0.342 * | |
| H. annuus | T1 | 0.940 | 0.634 | 0.306 |
| T2 | 0.350 * | 0.236 * | 0.114 * | |
| T3 | 2.380 * | 1.606 * | 0.774 * | |
| T4 | 1.401 * | 0.946 * | 0.455 * | |
| T5 | 1.960 * | 1.323 * | 0.637 * | |
| Treatments | Parameters | |||||||
|---|---|---|---|---|---|---|---|---|
| Cell Size (µm) | Cell Wall Thickness (µm) | Cytoplasm Size (µm) | Vacuole Size (µm) | Nucleus Size (µm) | ||||
| T1 | 165.25 | 0.0022 | 80.145 | 85.112 | 13.827 | |||
| T2 | 139.15 | 0.0081 | 60.537 | 78.617 | 25.910 * | |||
| T3 | 325.86 | 0.0097 | 204.031 * | 121.835 | 16.582 | |||
| T4 | 573.13 * | 0.012 * | 210.661 * | 362.442 * | 17.296 | |||
| T5 | 144.38 | 0.004 | 60.96 | 83.424 | 6.407 | |||
| Treatments | Chloroplast | Starch/Chloroplast | Mitochondria | |||||
| NO | Size (µm) | Total Size (µm) | NO | Size (µm) | NO | Size (µm) | Total Size (µm) | |
| T1 | 6 | 4.787 | 28.722 | 2 | 0.753 | 4 | 0.796 | 3.184 |
| T2 | 9.0 * | 7.967 * | 71.703 * | 2 | 1.869 * | 8 * | 0.834 | 6.672 * |
| T3 | 0.10 * | 8.009 * | 80.09 * | 0 | 0 | 3 * | 0.551 | 1.653 |
| T4 | 0.10 * | 14.058 * | 140.058 * | 3 * | 5.220 * | 5 * | 0.508 | 2.54 |
| Treatments | Parameters | |||||||
|---|---|---|---|---|---|---|---|---|
| Cell Size (µm) | Cell wall thickness (µm) | Cytoplasm Size (µm) | Vacuole Size (µm) | Nucleus Size (µm) | ||||
| T1 | 146.901 | 0.005 | 85.13 | 61.771 | 11.541 | |||
| T2 | 260.098 * | 0.007 | 177.676 * | 82.421 | 11.976 | |||
| T3 | 110.533 | 0.006 | 41.737 | 68.796 | 10.987 | |||
| T4 | 57.984 | 0.005 | 34.674 | 23.81 | 10.446 | |||
| T5 | 220.996 | 0.017 | 107.966 | 113.030 * | 14.924 | |||
| Treatments | Chloroplast | Starch/Chloroplast | Mitochondria | |||||
| Number | Size (µm) | Total Size (µm) | Number | Size (µm) | Number | Size (µm) | Total Size (µm) | |
| T1 | 8 | 5.548 | 44.384 | 0 | 0.000 | 4 | 0.216 | 0.864 |
| T2 | 13.000 * | 4.216 | 54.808 | 1 | 0.424 | 3 * | 0.430 | 1.290 |
| T3 | 8 | 4.066 | 32.048 | 1 | 0.286 | 4 | 0.456 * | 1.824 * |
| T4 | 14.000 * | 3.331 | 46.634 | 2 | 0.244 | 6 * | 0.384 | 2.304 * |
| T5 | 6.000 * | 8.825* | 52.950 | 1 | 0.268 | 3* | 0.450 * | 1.350 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alharbi, K.; Haroun, S.A.; Kazamel, A.M.; Abbas, M.A.; Ahmaida, S.M.; AlKahtani, M.; AlHusnain, L.; Attia, K.A.; Abdelaal, K.; Gamel, R.M.E. Physiological Studies and Ultrastructure of Vigna sinensis L. and Helianthus annuus L. under Varying Levels of Nitrogen Supply. Plants 2022, 11, 1884. https://doi.org/10.3390/plants11141884
Alharbi K, Haroun SA, Kazamel AM, Abbas MA, Ahmaida SM, AlKahtani M, AlHusnain L, Attia KA, Abdelaal K, Gamel RME. Physiological Studies and Ultrastructure of Vigna sinensis L. and Helianthus annuus L. under Varying Levels of Nitrogen Supply. Plants. 2022; 11(14):1884. https://doi.org/10.3390/plants11141884
Chicago/Turabian StyleAlharbi, Khadiga, Samia A. Haroun, Amany M. Kazamel, Mohammed A. Abbas, Safia M. Ahmaida, Muneera AlKahtani, Latifa AlHusnain, Kotb A. Attia, Khaled Abdelaal, and Rasha M. E. Gamel. 2022. "Physiological Studies and Ultrastructure of Vigna sinensis L. and Helianthus annuus L. under Varying Levels of Nitrogen Supply" Plants 11, no. 14: 1884. https://doi.org/10.3390/plants11141884