
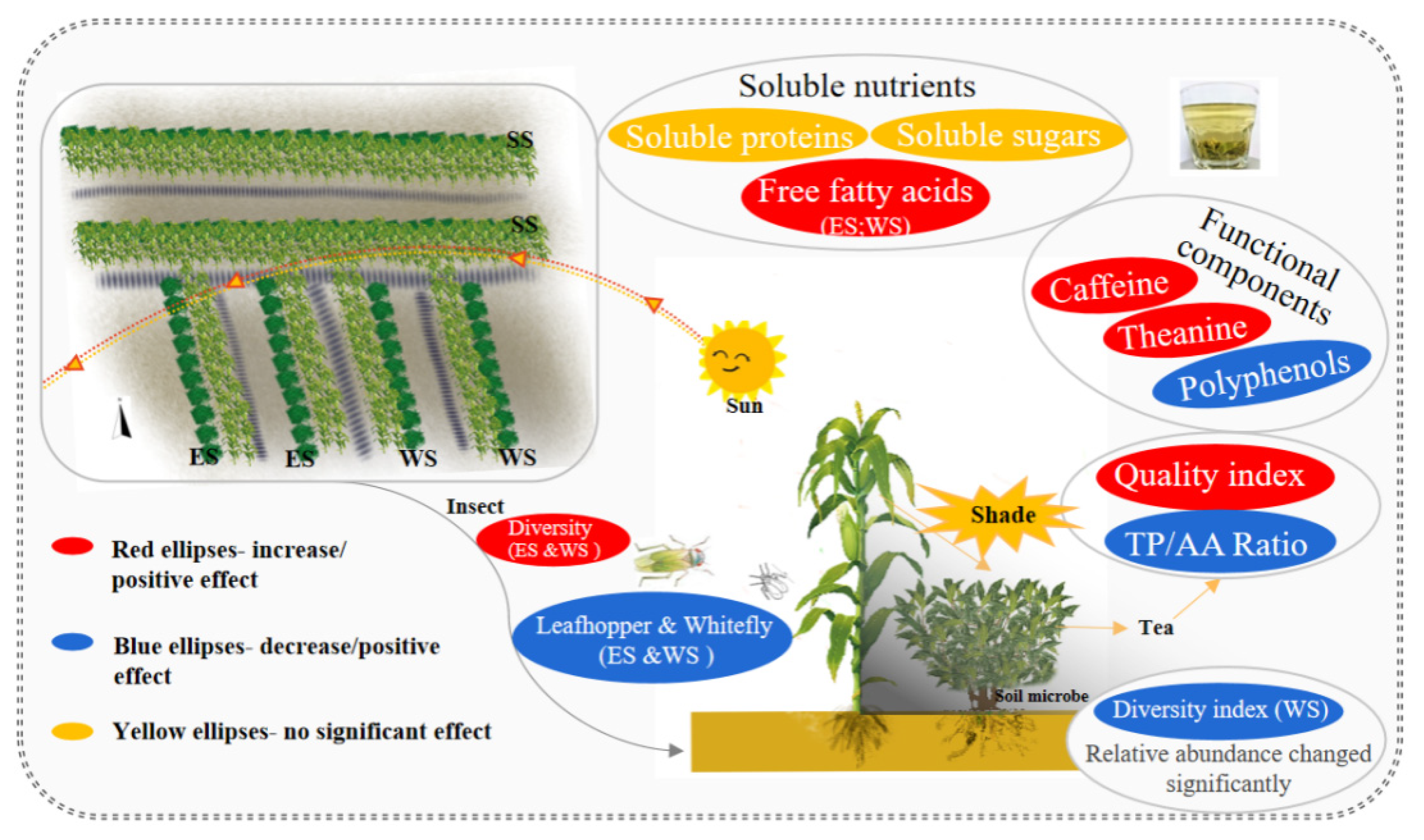
Impacts of Intercropped Maize Ecological Shading on Tea Foliar and Functional Components, Insect Pest Diversity and Soil Microbes




Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site Description
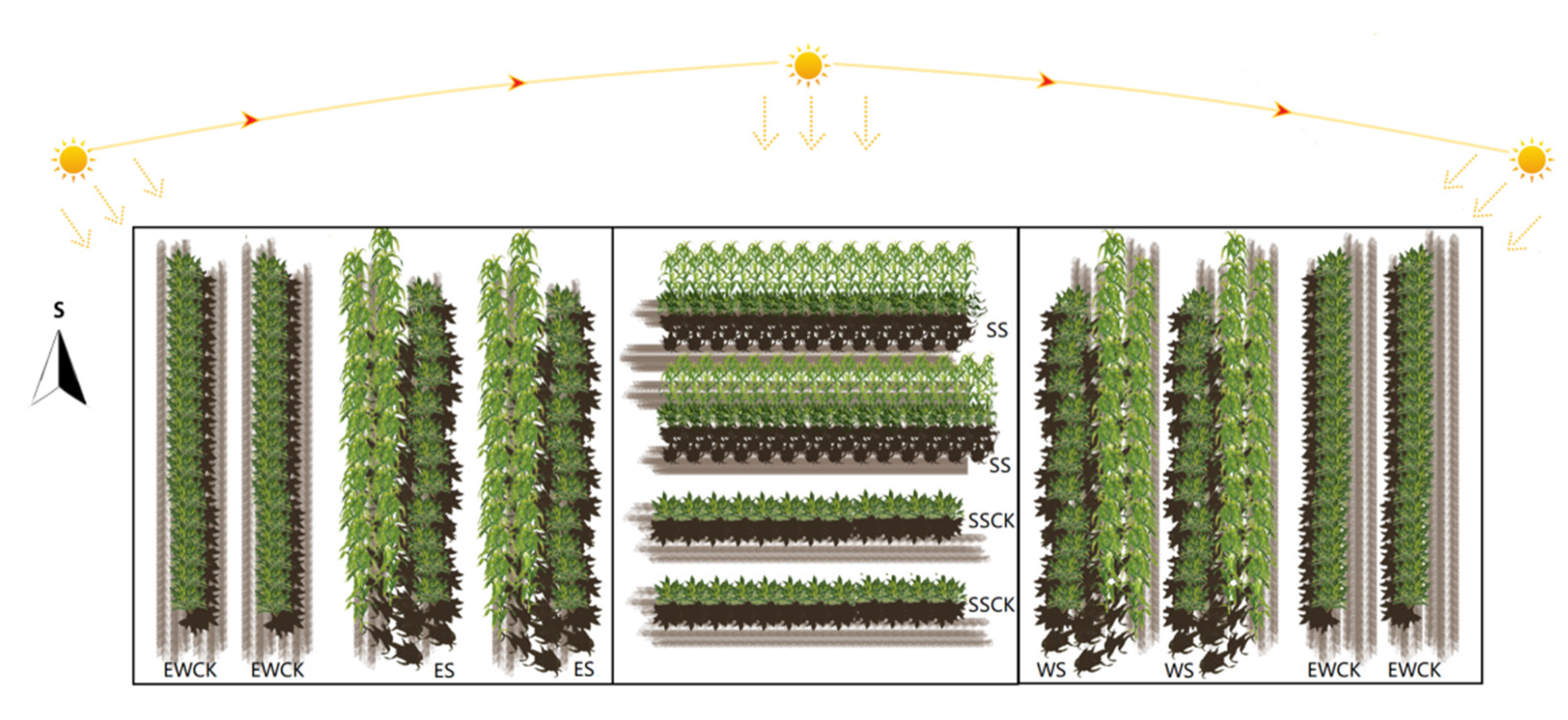
2.2. Maize Intercropping Setup
2.3. Determination of Nutritional and Functional Components in Tea Leaves
2.3.1. Foliar Nutrient Contents
2.3.2. Foliar Functional Component Contents
2.3.3. Leaf Quality Indexes
2.4. Insect Investigation
2.5. Composition and Diversity of Soil Microbial Community in the Tea Plantation
2.6. Data Analysis
3. Results
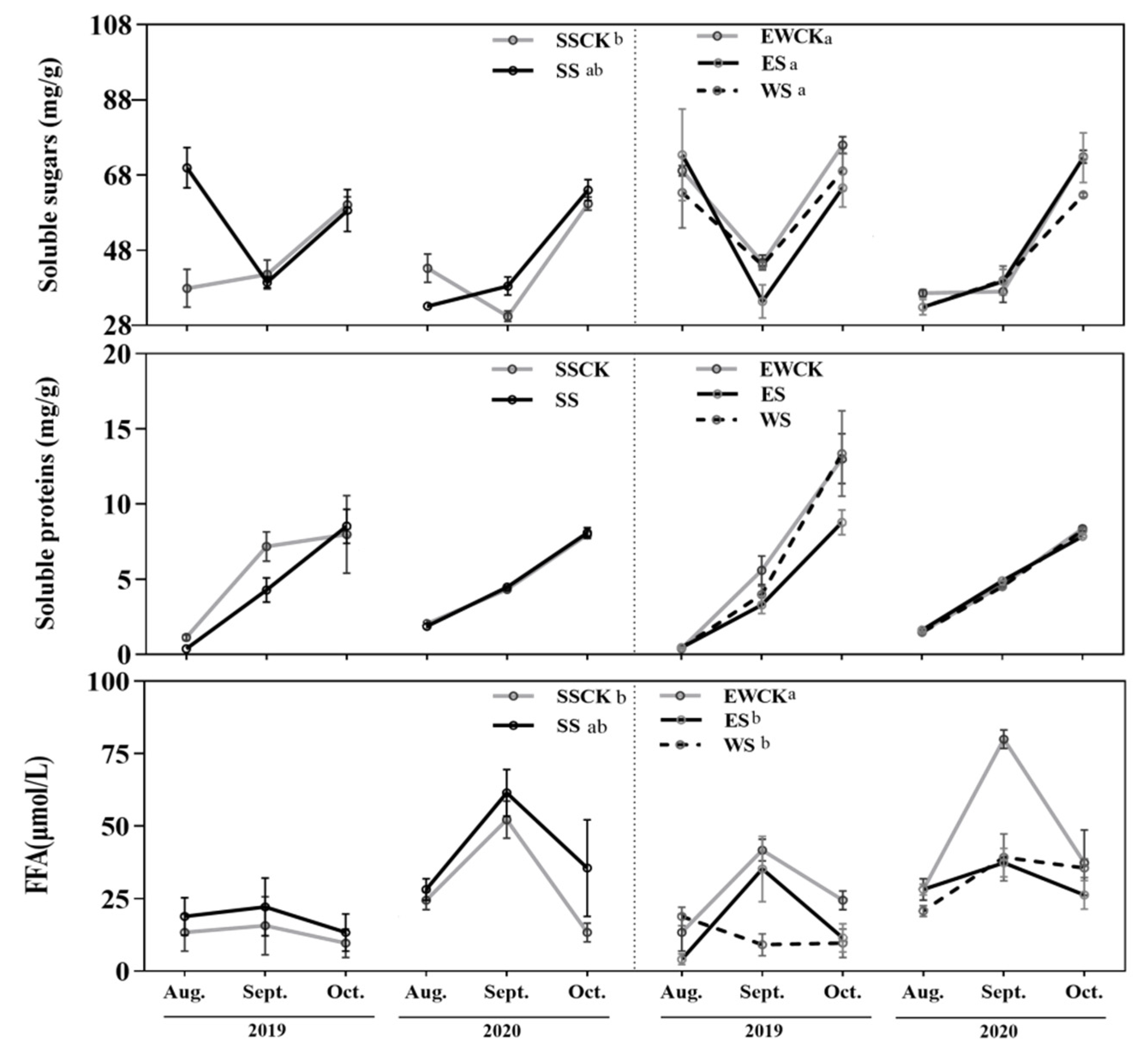
3.1. Effects of Intercropped Maize Ecological Shading on Foliar Soluble Nutrients of Tea Plants
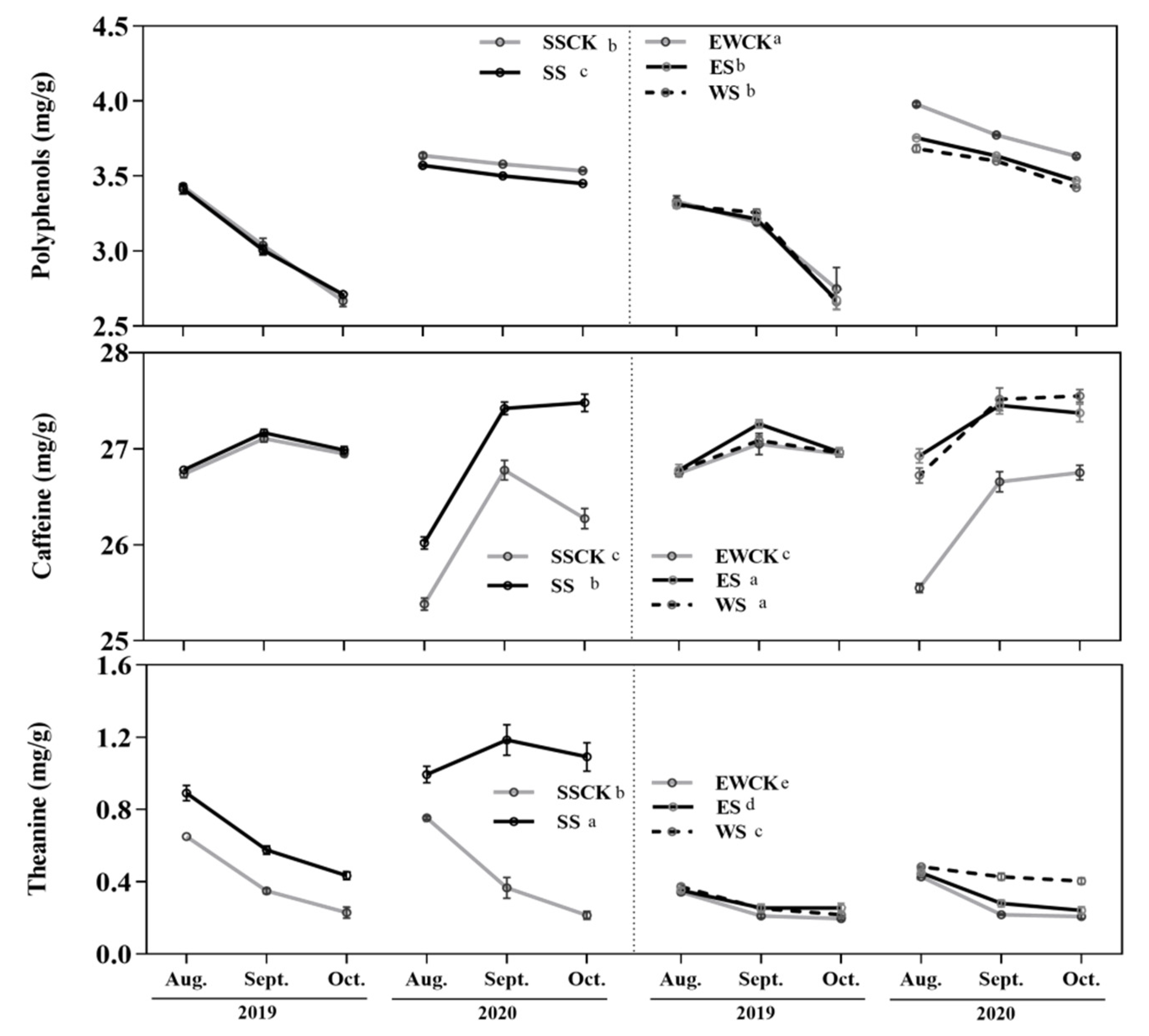
3.2. Effects of Intercropped Maize Ecological Shading on Foliar Functional Components of Tea Plants
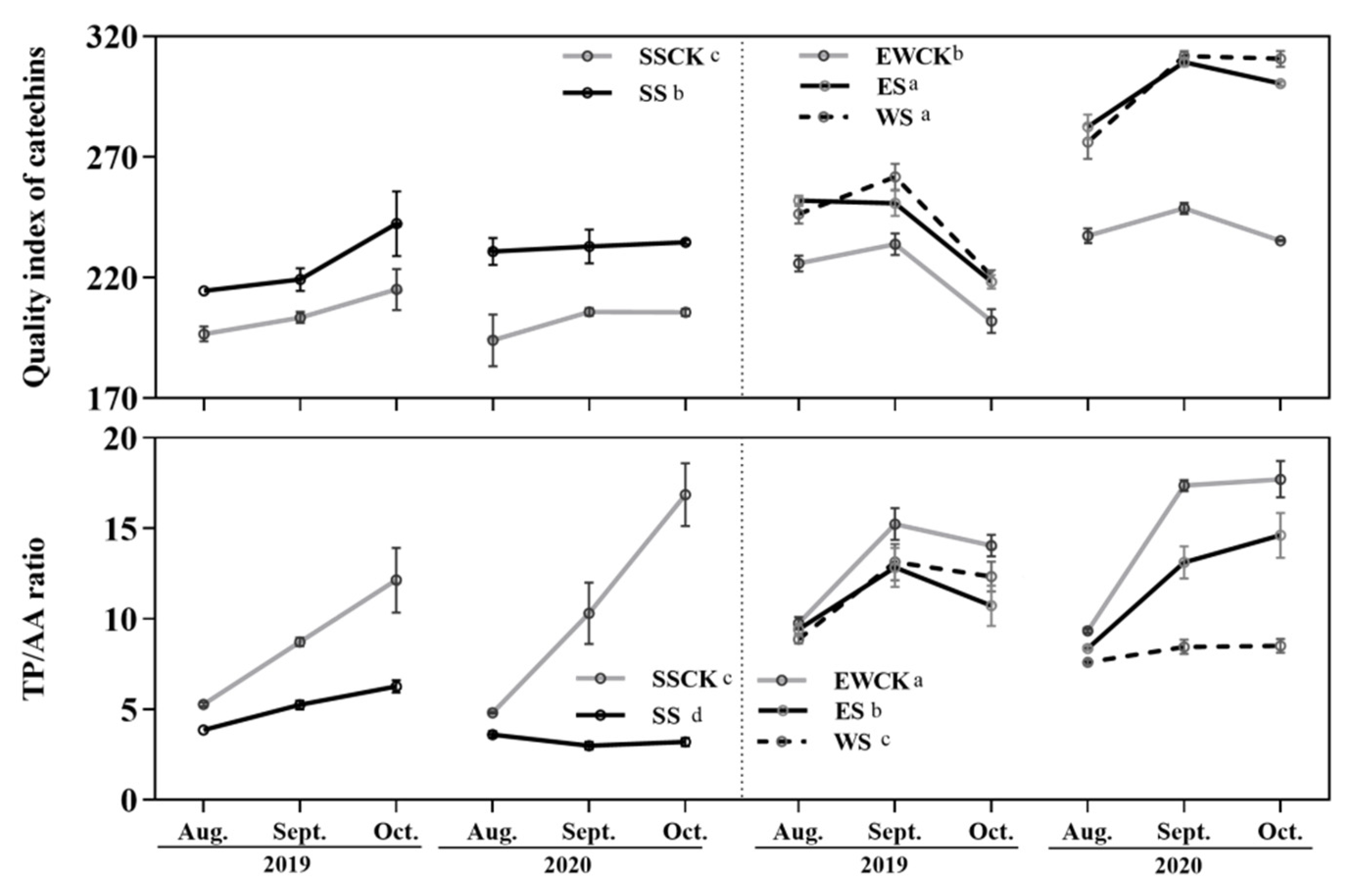
3.3. Effects of Intercropped Maize Ecological Shading on the Leaf Quality Indexes of Tea Plants
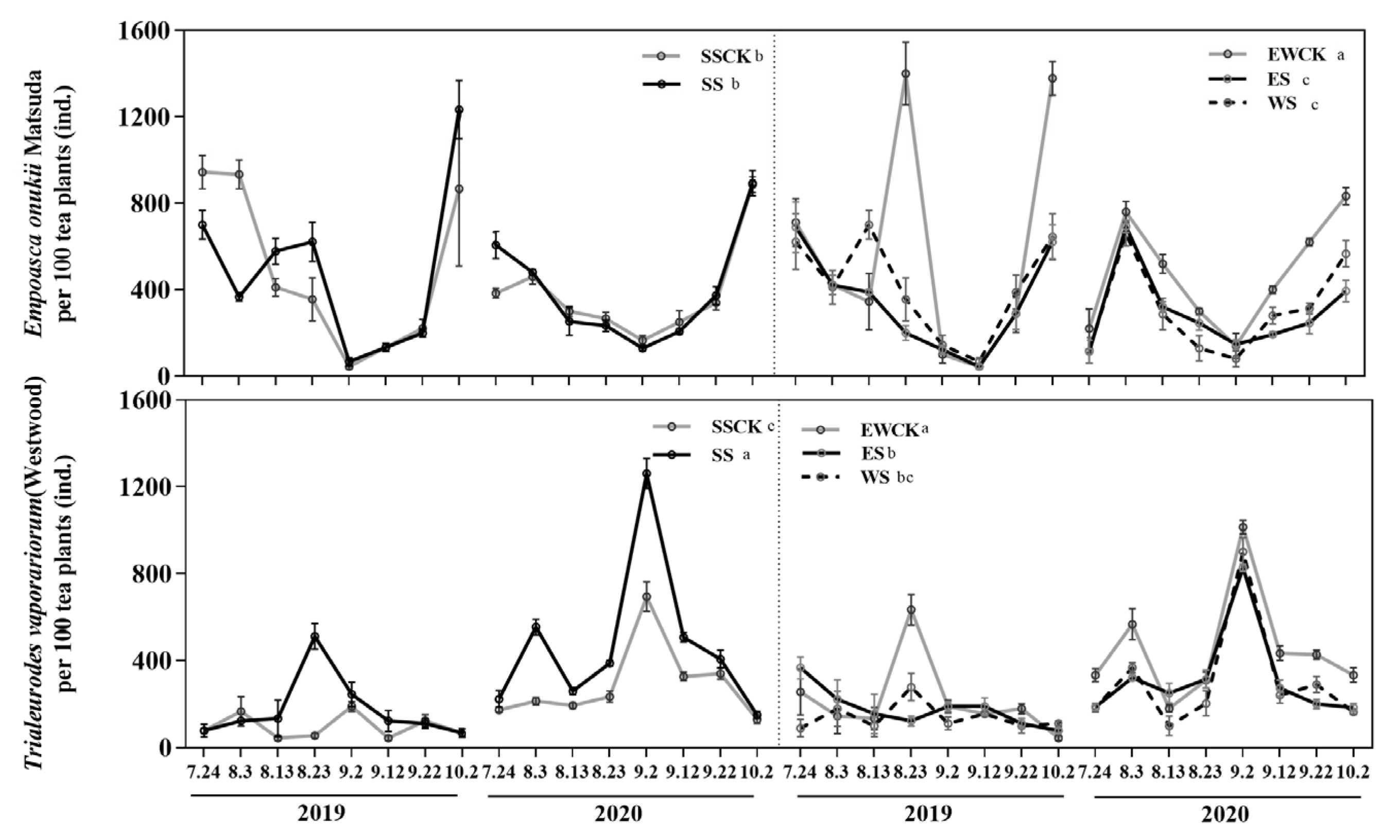
3.4. Effects of Intercropped Maize Ecological Shading on Population Dynamics of Key Tea Pests and Community Diversity in Tea Plantations
3.4.1. Population Dynamics of Key Insect Species
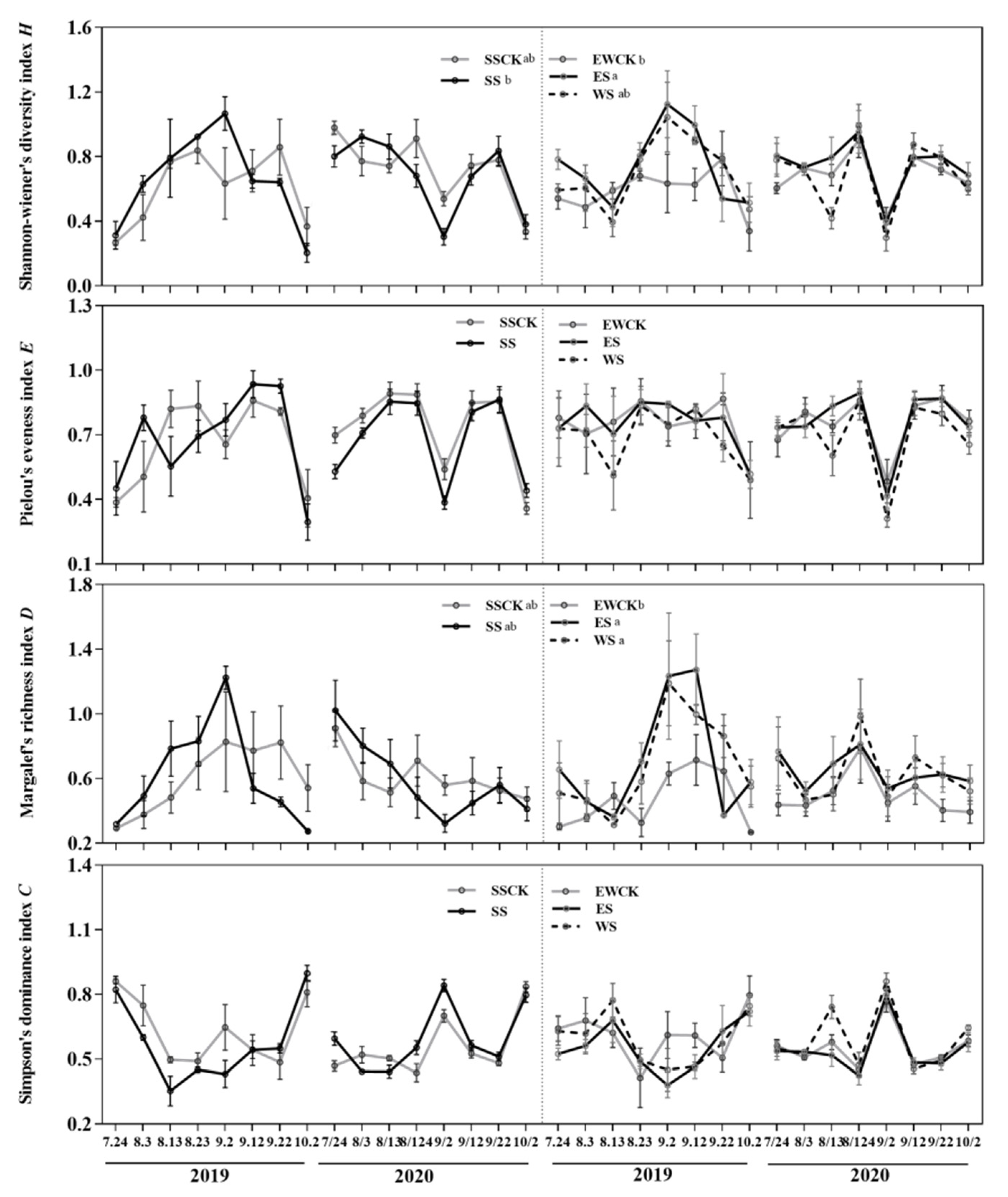
3.4.2. Community Diversity of Insects
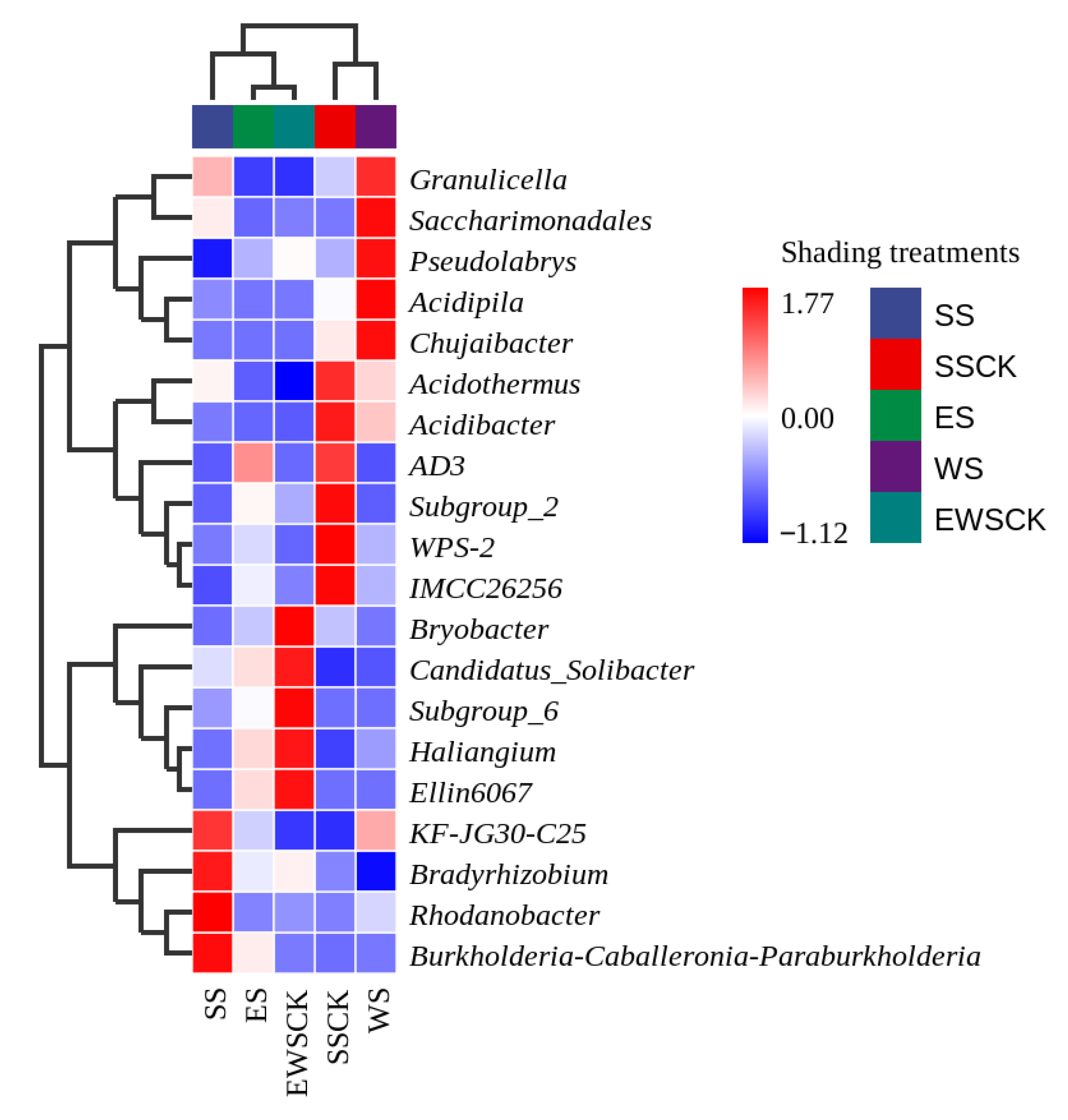
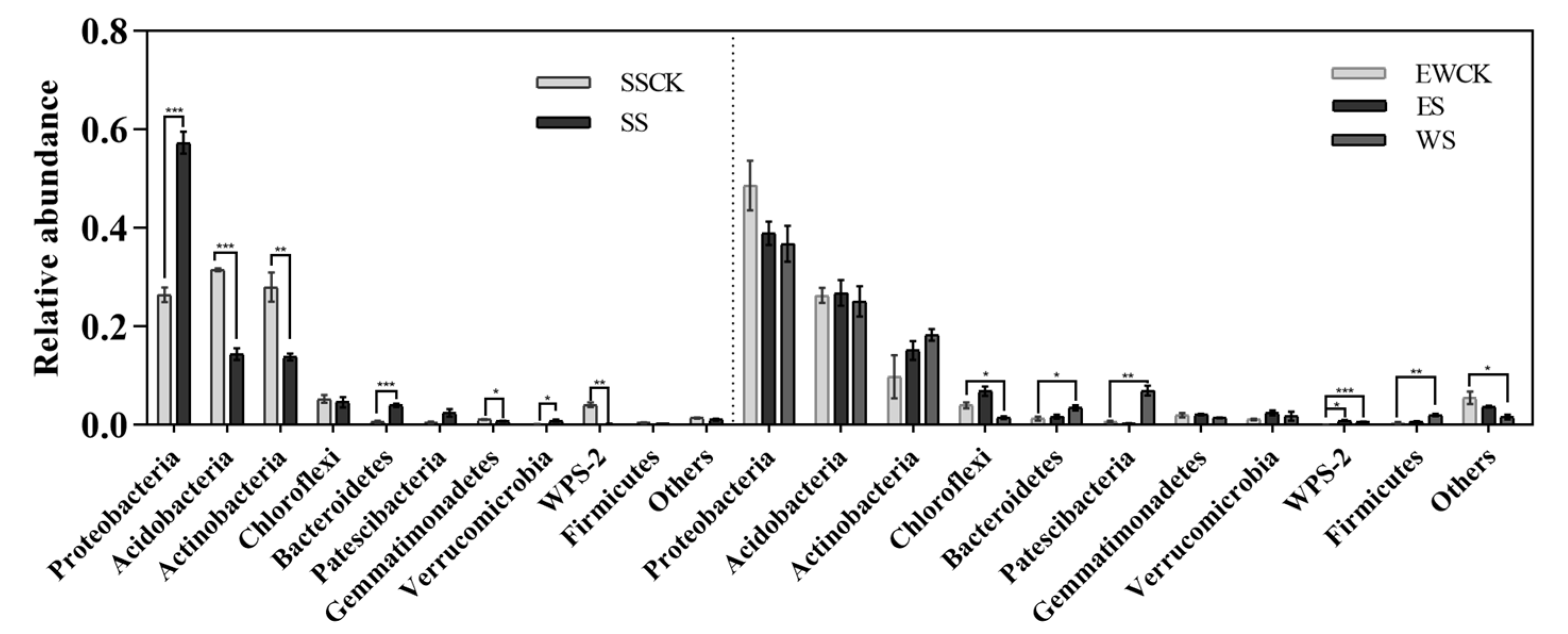
3.5. Effects of Intercropped Maize Ecological Shading on the Community Structure and Diversity of Soil Microorganisms in Tea Plantation
3.5.1. Taxonomic Composition of Soil Microorganisms
3.5.2. Community Diversity of Soil Microbial Microorganisms
4. Discussion
4.1. Effects of Ecological Shade by Intercropping Maize on Foliar Soluble Nutrients of Tea Plants
4.2. Effects of Maize Intercropped Ecological Shading on Foliar Functional Components and Leaf Quality of Tea Plants
4.3. Effects of Maize Intercropped Ecological Shading on Population Dynamics and the Community Diversity of Key Tea Pests in Tea Plantations
4.4. Effects of Maize Intercropped Ecological Shading on Community Structure and Diversity of Soil Microorganisms in Tea Plantations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Measuring Time | Ecological Shading Treatment (klx) | The Control (klx) | F/p |
|---|---|---|---|---|
| ES vs. EWCK | 6:00 a.m. to 10:30 a.m. | 13.90 ± 9.3 | 103.91 ± 50.9 | 1625.21/<0.001 *** |
| WS vs. EWCK | 2:00 p.m. to 6:00 p.m. | 18.73 ± 13.83 | 98.15 ± 58.09 | 679.97/<0.001 *** |
| SS vs. SSCK | 11:00 a.m. to 1:30 p.m. | 28.46 ± 12.99 | 170.13 ± 10.83 | 56.52/<0.001 *** |
| Measured Indexes | SS/SSCK | ES/EWCK | WS/EWCK | SSCK/EWCK | SS/ES | SS/WS | ES/WS | Ecological Shading Effects | Rowing-Direction Effects | |
|---|---|---|---|---|---|---|---|---|---|---|
| Foliar soluble nutrients | Soluble sugars (mg/g) | / | / | |||||||
| Soluble proteins (mg/g) | / | / | ||||||||
| Free fatty acids (mmol/L) | −(ES & WS) | −(ES & WS) | ||||||||
| Foliar functional components | Polyphenols (mg/g) | −(ES & WS > SS); ES & WS > SS | −(ES & WS > SS); ES & WS > SS | |||||||
| Caffeine (mg/g) | +(ES & WS > SS); ES & WS > SS | +(ES & WS > SS); ES & WS > SS | ||||||||
| Theanine (μg/g) | +(SS > ES > WS); SS > ES > WS | +(SS > ES > WS); SS > ES > WS | ||||||||
| Leaf quality | Catechin quality index | +(ES & WS > SS); ES & WS > SS | +(ES & WS > SS); ES & WS > SS | |||||||
| Phenol/ammonia ratio | −(ES, WS, SS); ES > WS > SS | −(ES, WS, SS); ES > WS > SS | ||||||||
| Population dynamics | E. onukii | −(ES & WS); SS > ES & WS | −(ES & WS); SS > ES & WS | |||||||
| T. vaporariorum | +(SS); −(ES & WS); SS > ES & WS | +(SS); −(ES & WS); SS > ES & WS | ||||||||
| Community diversity of insects | Shannon-Wiener index (H) | +(ES); ES > SS & WS | +(ES); ES > SS & WS | |||||||
| Pielou evenness index (E) | / | / | ||||||||
| Margalef richness index (D) | +(ES & WS) | +(ES & WS) | ||||||||
| Simpson dominance index (C) | / | / | ||||||||
| Community diversity of soil microbial microorganisms | Shannon-Wiener index (H) | −(WS); ES & SS > WS | −(WS); ES & SS > WS | |||||||
| Pielou evenness index (E) | −(WS); ES > SS > WS | −(WS); ES > SS > WS | ||||||||
| Chao1 index | −(WS); ES > SS > WS | −(WS); ES > SS > WS | ||||||||
| Simpson dominance index (C) | −(WS); ES > SS > WS | −(WS); ES > SS > WS | ||||||||
 Significant decrease;
Significant decrease;  Significant increase;
Significant increase;  No significant difference; + Increase; − Decrease; / no effect.
No significant difference; + Increase; − Decrease; / no effect.References
- Yang, T.; Xie, Y.; Lu, X.; Yan, X.; Wang, Y.; Ma, J.; Cheng, X.; Lin, S.; Bao, S.; Wan, X.; et al. Shading promoted theanine biosynthesis in the roots and allocation in the shoots of the tea plant (Camellia sinensis L.) Cultivar Shuchazao. J. Agric. Food Chem. 2021, 69, 4795–4803. [Google Scholar] [CrossRef] [PubMed]
- Drew, L. The growth of tea. Nature 2019, 566, S2–S4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teng, R.-M.; Wang, Y.-X.; Li, H.; Lin, S.-J.; Liu, H.; Zhuang, J. Effects of shading on lignin biosynthesis in the leaf of tea plant (Camellia sinensis (L.) O. Kuntze). Mol. Genet. Genom. 2020, 296, 165–177. [Google Scholar] [CrossRef]
- Wu, T.; Zou, R.; Pu, D.; Lan, Z.; Zhao, B. Non-targeted and targeted metabolomics profiling of tea plants (Camellia sinensis) in response to its intercropping with Chinese chestnut. BMC Plant Biol. 2021, 21, 55. [Google Scholar] [CrossRef] [PubMed]
- Qin, Z.M.; Tanui, J.; Feng, W.Y.; Wang, Y.H.; Xiao, R.L.; Li, X.H. Efects of shading on yield index and biochemical components of tea in hilly tea plantation. J. Nanjing Agric. Univ. 2011, 34, 47–52. [Google Scholar]
- Wang, Y.; Yang, X.; Li, K.; Li, C.; Li, L.; Li, J.; Huang, H.; He, Y.; Ye, C.; Song, X. Simultaneous determination of theanine, gallic acid, purine alkaloids, catechins, and theaflavins in black tea using HPLC. Int. J. Food Sci. Technol. 2010, 45, 1263–1269. [Google Scholar] [CrossRef]
- Cheng, S.; Fu, X.; Wang, X.; Liao, Y.; Zeng, L.; Dong, F.; Yang, Z. Studies on the Biochemical Formation Pathway of the Amino Acid l-Theanine in Tea (Camellia sinensis) and Other Plants. J. Agric. Food Chem. 2017, 65, 7210–7216. [Google Scholar] [CrossRef]
- Gurr, G.M.; Wratten, S.D.; Landis, D.A.; You, M. Habitat Management to Suppress Pest Populations: Progress and Prospects. Annu. Rev. Entomol. 2017, 62, 91–109. [Google Scholar] [CrossRef]
- Roy, S.; Muraleedharan, N.; Mukhopadhyay, A. The red spider mite, Oligonychus coffeae (Acari: Tetranychidae): Its status, biology, ecology and management in tea plantations. Exp. Appl. Acarol. 2014, 63, 431–463. [Google Scholar] [CrossRef]
- De Costa, W.A.J.M.; Surenthran, P. Tree-crop interactions in hedgerow intercropping with different tree species and tea in Sri Lanka: 1. Production and resource competition. Agrofor. Syst. 2005, 63, 199–209. [Google Scholar] [CrossRef]
- Farooq, T.; Kumar, U.; Mo, J.; Shakoor, A.; Wang, J.; Rashid, M.; Tufail, M.; Chen, X.; Yan, W. Intercropping of Peanut–Tea Enhances Soil Enzymatic Activity and Soil Nutrient Status at Different Soil Profiles in Subtropical Southern China. Plants 2021, 10, 881. [Google Scholar] [CrossRef]
- Dingha, B.N.; Jackai, L.E.; Amoah, B.A.; Akotsen-Mensah, C. Pollinators on Cowpea Vigna unguiculata: Implications for Intercropping to Enhance Biodiversity. Insects 2021, 12, 54. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.L.; Yuan, P.; Pozsgai, G.; Chen, P.; Zhu, H.; You, M.S. The impact of cover crops on the predatory mite Anystis baccarum (Acari, Anystidae) and the leafhopper pest Empoasca onukii (Hemiptera, Cicadellidae) in a tea plantation. Pest. Manag. Sci. 2019, 75, 3371–3380. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhou, Y.; Zhou, B.; Tang, H.; Chen, Y.; Qiao, X.; Tang, J. Habitat management as a safe and effective approach for improving yield and quality of tea (Camellia sinensis) leaves. Sci. Rep. 2019, 9, 433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, J.; Luo, Y.; Sun, P.; Gao, J.; Zhao, D.; Yang, P.; Hu, T. Effects of shade stress on turfgrasses morphophysiology and rhizosphere soil bacterial communities. BMC Plant Biol. 2020, 20, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M.; Lester, P. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteu reagent. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1999; Volume 299, pp. 152–178. [Google Scholar]
- Kim, Y.-S.; Sano, H. Pathogen resistance of transgenic tobacco plants producing caffeine. Phytochemistry 2008, 69, 882–888. [Google Scholar] [CrossRef]
- Su, Y.L.; Leung, L.K.; Huang, Y.; Chen, Z.-Y. Stability of tea theaflavins and catechins. Food Chem. 2003, 83, 189–195. [Google Scholar] [CrossRef]
- Wang, H.F.; Tsai, Y.S.; Lin, M.L.; Ou, A.S.-M. Comparison of bioactive components in GABA tea and green tea produced in Taiwan. Food Chem. 2006, 96, 648–653. [Google Scholar] [CrossRef]
- Vuong, Q.V.; Golding, J.B.; Nguyen, M.; Roach, P.D. Extraction and isolation of catechins from tea. J. Sep. Sci. 2010, 33, 3415–3428. [Google Scholar] [CrossRef]
- Li, X.; Ahammed, G.J.; Li, Z.-X.; Zhang, L.; Wei, J.-P.; Shen, C.; Yan, P.; Zhang, L.-P.; Han, W.-Y. Brassinosteroids Improve Quality of Summer Tea (Camellia sinensis L.) by Balancing Biosynthesis of Polyphenols and Amino Acids. Front. Plant Sci. 2016, 7, 1304. [Google Scholar] [CrossRef] [Green Version]
- Tikkanen, M.; Nurmi, M.; Kangasjärvi, S.; Aro, E.M. Core protein phosphorylation facilitates the repair of photodamaged photosystem II at high light. Biochim. Biophys. Acta 2018, 1777, 1432–1437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.-E.; Liu, W.-J.; Su, Y.-Q.; Cui, J.-M.; Zhang, Z.-W.; Yuan, M.; Zhang, H.-Y.; Yuan, S. Different response of photosystem II to short and long-term drought stress in Arabidopsis thaliana. Physiol. Plant. 2016, 158, 225–235. [Google Scholar] [CrossRef]
- Asada, K. Production and Scavenging of Reactive Oxygen Species in Chloroplasts and Their Functions. Plant Physiol. 2006, 141, 391–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; Van Breusegem, F. Reactive oxygen gene network of plants. Trends Plant Sci. 2004, 9, 490–498. [Google Scholar] [CrossRef] [PubMed]
- Nixon, P.J.; Michoux, F.; Yu, J.; Boehm, M.; Komenda, J. Recent advances in understanding the assembly and repair of photosystem II. Ann. Bot. 2010, 106, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sano, S.; Takemoto, T.; Ogihara, A.; Suzuki, K.; Masumura, T.; Satoh, S.; Takano, K.; Mimura, Y.; Morita, S. Stress Responses of Shade-Treated Tea Leaves to High Light Exposure after Removal of Shading. Plants 2020, 9, 302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohotti, A.; Lawlor, D. Diurnal variation of photosynthesis and photoinhibition in tea: Effects of irradiance and nitrogen supply during growth in the field. J. Exp. Bot. 2002, 53, 313–322. [Google Scholar] [CrossRef]
- Toshikazu, S.; Takashi, I.; Akio, M. The effect of shading in winter on the carbohydrate content of tea plants and productivity of first crop of tea. Jpn. J. Crop Sci. 2013, 82, 345–352. [Google Scholar]
- Li, Y.; Chen, C.; Li, Y.; Ding, Z.; Shen, J.; Wang, Y.; Zhao, L.; Xu, M. The identification and evaluation of two different color variations of tea. J. Sci. Food Agric. 2016, 96, 4951–4961. [Google Scholar] [CrossRef]
- Kito, M.; Kokura, H.; Izaki, J.; Sasaoka, K. Theanine, a precursor of the phloroglucinol nucleus of catechins in tea plants. Phytochemistry 1968, 7, 599–603. [Google Scholar] [CrossRef]
- Deng, W.-W.; Fei, Y.; Wang, S.; Wan, X.-C.; Zhang, Z.-Z.; Hu, X.-Y. Effect of shade treatment on theanine biosynthesis in Camellia sinensis seedlings. Plant Growth Regul. 2013, 71, 295–299. [Google Scholar] [CrossRef]
- Zhang, Q.; Shi, Y.; Ma, L.; Yi, X.; Ruan, J. Metabolomic Analysis Using Ultra-Performance Liquid Chromatography-Quadrupole-Time of Flight Mass Spectrometry (UPLC-Q-TOF MS) Uncovers the Effects of Light Intensity and Temperature under Shading Treatments on the Metabolites in Tea. PLoS ONE 2014, 9, e112572. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Hu, R.; Sreedasyam, A.; Garcia, T.M.; Lipzen, A.; Wang, M.; Yerramsetty, P.; Liu, D.; Ng, V.; Schmutz, J.; et al. Light-responsive expression atlas reveals the effects of light quality and intensity in Kalanchoë fedtschenkoi, a plant with crassulacean acid metabolism. Gigascience 2020, 9, giaa018. [Google Scholar] [CrossRef] [PubMed]
- Beillouin, D.; Ben-Ari, T.; Malézieux, E.; Seufert, V.; Makowski, D. Positive but variable effects of crop diversification on biodiversity and ecosystem services. Glob. Chang. Biol. 2021, 27, 4697–4710. [Google Scholar] [CrossRef]
- Das, S.; Sarker, M.; Mukhopadhyay, A. Changing diversity of hymenopteran parasitoids from organically and conventionally managed tea-ecosystem of North Bengal, India. J. Environ. Biol. 2005, 26, 505–509. [Google Scholar]
- Rojas, L.; Godoy, C.; Hanson, P.; Hilje, L. A survey of homopteran species (Auchenorrhyncha) from coffee shrubs and poró and laurel trees in shaded coffee plantations, in Turrialba, Costa Rica. Rev. De Biol. Trop. 2002, 49, 1057–1065. [Google Scholar]
- Song, Y.; Liu, J.; Chen, F. Azotobacter chroococcum inoculation can improve plant growth and resistance of maize to armyworm, Mythimna separata even under reduced nitrogen fertilizer application. Pest. Manag. Sci. 2020, 76, 4131–4140. [Google Scholar] [CrossRef]
- Tian, Y.; Zhao, Y.; Zhang, L.; Mu, W.; Zhang, Z. Morphological, Physiological, and Biochemical Responses of Two Tea Cultivars to Empoasca onukii (Hemiptera: Cicadellidae) Infestation. J. Econ. Entomol. 2018, 111, 899–908. [Google Scholar] [CrossRef]
- Liao, Y.; Yu, Z.; Liu, X.; Zeng, L.; Cheng, S.; Li, J.; Tang, J.; Yang, Z. Effect of Major Tea Insect Attack on Formation of Quality-Related Nonvolatile Specialized Metabolites in Tea (Camellia sinensis) Leaves. J. Agric. Food Chem. 2019, 67, 6716–6724. [Google Scholar] [CrossRef]
- Kim, Y.-S.; Uefuji, H.; Ogita, S.; Sano, H. Transgenic tobacco plants producing caffeine: A potential new strategy for insect pest control. Transgenic Res. 2006, 15, 667–672. [Google Scholar] [CrossRef]
- Zelles, L. Fatty acid patterns of phospholipids and lipopolysaccharids in the characterization of microbial communities in soil, a review. Biol. Fert. Soils 1999, 29, 111–129. [Google Scholar] [CrossRef]
- Neal, A.L.; Hughes, D.; Clark, I.M.; Jansson, J.K.; Hirsch, P.R. Microbiome Aggregated Traits and Assembly Are More Sensitive to Soil Management than Diversity. Msystems 2021, 6, e01056-20. [Google Scholar] [CrossRef] [PubMed]
- Shigyo, N.; Umeki, K.; Hirao, T. Plant functional diversity and soil properties control elevational diversity gradients of soil bacteria. FEMS Microbiol. Ecol. 2019, 95, fiz025. [Google Scholar] [CrossRef]
- Li, N.; Gao, D.; Zhou, X.; Chen, S.; Li, C.; Wu, F. Intercropping with Potato-Onion Enhanced the Soil Microbial Diversity of Tomato. Microorganisms 2020, 8, 834. [Google Scholar] [CrossRef]
- Dang, K.; Gong, X.; Zhao, G.; Wang, H.; Ivanistau, A.; Feng, B. Intercropping Alters the Soil Microbial Diversity and Community to Facilitate Nitrogen Assimilation: A Potential Mechanism for Increasing Proso Millet Grain Yield. Front. Microbiol. 2020, 11, 601054. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Jiang, J.; Huang, Z.; Wu, H.; Wang, J.; He, L.; Xiong, F.; Zhong, R.; Liu, J.; Han, Z.; et al. Sugarcane/peanut intercropping system improves the soil quality and increases the abundance of beneficial microbes. J. Basic Microbiol. 2021, 61, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Li, T.; Zheng, Z.; Chen, H.Y. Soil aggregate-associated bacterial metabolic activity and community structure in different aged tea plantations. Sci. Total Environ. 2018, 654, 1023–1032. [Google Scholar] [CrossRef]
- Jeanneret, P.; Aviron, S.; Alignier, A.; Lavigne, C.; Helfenstein, J.; Herzog, F.; Kay, S.; Petit, S. Agroecology landscapes. Landsc. Ecol. 2021, 36, 2235–2257. [Google Scholar] [CrossRef]









| Measured Indexes | Ecological Shading Treatments (S) | Sampling Years (Y) | S × Y | |
|---|---|---|---|---|
| Foliar soluble nutrients | Soluble sugars (mg/g) | 3.83/0.02 * | 32.56/<0.001 *** | 1.66/0.20 |
| Soluble proteins (mg/g) | 1.73/0.18 | 2.09/0.16 | 1.75/0.18 | |
| Free fatty acids (μmol/L) | 5.39/0.004 ** | 53.80/<0.001 *** | 0.45/0.77 | |
| Foliar functional components | Polyphenols (mg/g) | 24.20/<0.001 *** | 2331.38/<0.001 *** | 13.29/<0.001 *** |
| Caffeine (mg/g) | 118.21/<0.001 *** | 51.33/<0.001 *** | 97.48/<0.001 *** | |
| Theanine (μg/g) | 379.93/<0.001 *** | 168.34/<0.001 *** | 54.42<0.001 *** | |
| Leaf quality | Catechin quality index | 151.57/<0.001 *** | 171.59/<0.001 *** | 35.33/<0.001 *** |
| Phenol/ammonia ratio | 148.85/<0.001 *** | 0.06/0.80 | 16.06/<0.001 *** | |
| Population dynamics | Empoasca onukii | 30.61/<0.001 *** | 52.62/<0.001 *** | 0.73/0.58 |
| Trialeurodes vaporariorum | 37.00/<0.001 *** | 514.35/<0.001 *** | 9.04/<0.001 *** | |
| Community diversity of insects | Shannon-Wiener index (H) | 2.76/0.04 * | 5.15/0.03 * | 1.44/0.25 |
| Pielou evenness index (E) | 2.64/0.05 | 1.08/0.31 | 0.46/0.77 | |
| Margalef richness index (D) | 2.86/0.04 * | 0.25/0.62 | 0.19/0.94 | |
| Simpson dominance index (C) | 2.51/0.06 | 6.34/0.01 * | 2.57/0.06 | |
| Diversity Indices | SSCK | SS | EWCK | ES | WS | F/P |
|---|---|---|---|---|---|---|
| Chao1 index | 6539 ± 61 ab | 6556 ± 150 ab | 7900 ± 805 a | 7784 ± 242 a | 5078 ± 222 b | 8.42/0.003 ** |
| Shannon-Wiener index (H) | 10.38 ± 0.07 b | 10.23 ± 0.19 b | 11.18 ± 0.22 a | 11.45 ± 0.03 a | 9.31 ± 0.10 c | 52.78/<0.001 *** |
| Pielou evenness index (E) | 0.840 ± 0.004 b | 0.829 ± 0.010 b | 0.887 ± 0.104 a | 0.898 ± 0.001 a | 0.873 ± 0.005 c | 43.26/<0.001 *** |
| Simpson dominance index (C) | 0.995 ± 0.002 b | 0.995 ± 0.001 b | 0.999 ± 0.001 a | 0.999 ± 0.001 a | 0.991 ± 0.001 c | 27.43/<0.001 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zou, Y.; Shen, F.; Zhong, Y.; Lv, C.; Pokharel, S.S.; Fang, W.; Chen, F. Impacts of Intercropped Maize Ecological Shading on Tea Foliar and Functional Components, Insect Pest Diversity and Soil Microbes. Plants 2022, 11, 1883. https://doi.org/10.3390/plants11141883
Zou Y, Shen F, Zhong Y, Lv C, Pokharel SS, Fang W, Chen F. Impacts of Intercropped Maize Ecological Shading on Tea Foliar and Functional Components, Insect Pest Diversity and Soil Microbes. Plants. 2022; 11(14):1883. https://doi.org/10.3390/plants11141883
Chicago/Turabian StyleZou, Yan, Fangyuan Shen, Yanni Zhong, Changning Lv, Sabin Saurav Pokharel, Wanping Fang, and Fajun Chen. 2022. "Impacts of Intercropped Maize Ecological Shading on Tea Foliar and Functional Components, Insect Pest Diversity and Soil Microbes" Plants 11, no. 14: 1883. https://doi.org/10.3390/plants11141883